

Зоологический журнал, 2019, T. 98, № 6, стр. 673-683
Экспериментальный сравнительный анализ охотничьего поведения четырех видов хомячков подсемейства Cricetinae
Я. В. Левенец 1, *, С. Н. Пантелеева 1, 3, Ж. И. Резникова 1, 3, А. В. Гуреева 2, Н. Ю. Феоктистова 2, А. В. Суров 2
1 Институт систематики и экологии животных Сибирского отделения РАН
630091 Новосибирск, Россия
2 Институт проблем экологии и эволюции им А.Н. Северцова РАН
119071 Москва, Россия
3 Новосибирский национальный исследовательский государственный университет
630090 Новосибирск, Россия
* E-mail: jan.levenets@gmail.com
Поступила в редакцию 31.05.2018
После доработки 29.08.2018
Принята к публикации 13.12.2018
Аннотация
В лабораторных экспериментах исследовали охотничье поведение по отношению к насекомым у двух видов мохноногих хомячков – джунгарского (Phodopus sungorus Pallas 1773) и хомячка Кэмпбелла (P. сampbelli Thomas 1905) и двух представителей группы эверсманновых хомячков – хомячка Эверсманна (Allocricetulus eversmanni Brandt 1859) и монгольского (A. curtatus Allen 1925). У всех видов впервые детально описано целенаправленное взаимодействие с подвижной добычей по типу “хищник–жертва”. Стереотипы охотничьего поведения оказались у них сравнимыми с таковыми у специализированных хищных хомячков. У всех видов в четверти случаев наблюдалось начало атаки с захвата лапами, что является признаком эволюционно более продвинутого охотничьего поведения, чем захват зубами. Самый простой охотничий стереотип выявлен у джунгарского хомячка, у остальных видов отмечены черты специализации в виде разных форм манипуляторной активности. Наиболее стремительную и эффективную атаку на добычу демонстрирует монгольский хомячок. Облигатное и не требующее предварительного опыта проявление паттерна у эверсманновых хомячков по-видимому говорит о большей специализации хищнического поведения у этой эволюционно более молодой группы по сравнению с мохноногими хомячками, у которых атаки на добычу проявляются факультативно и совершенствуются с приобретением опыта.
Грызуны, являясь преимущественно растительноядными животными, как правило, включают в рацион пищу и животного происхождения (Landry, 1970). Главным образом это мелкие беспозвоночные (кольчатые черви, моллюски, насекомые), но некоторые виды употребляют в пищу и более крупные объекты: ракообразных, рыб, амфибий, рептилий, птиц и даже млекопитающих (Башенина, 1977; Babinska-Werka, Garbarczyk, 1981). Если для истинно хищных животных стереотипные комплексы охотничьего поведения подробно описаны (Caro, 1980; обзоры в: Зорина и др., 2013; Резникова, 2016), то для грызунов реакции на подвижную добычу описаны у ограниченного количества видов, а стереотипизация охотничьего поведения у них почти не исследована.
Среди грызунов поведенческие адаптации к активной охоте на беспозвоночных были ранее экспериментально изучены у южного (Onychomys torridus Coues 1874) и северного (O. leucogaster Wied-Neuwied 1841) кузнечиковых хомячков (Langley, 1986, 1994), всеядных белоногого (Peromyscus leucopus Rafinesque 1818) и оленьего хомячков (P. maniculatus Wagner 1845) (Kreiter, Timberlake, 1988), сирийского хомяка (Mesocricetus auratus Waterhouse 1839) (Polsky, 1977). Роль индивидуального опыта оказалась различна: у хищных кузнечиковых хомячков охотничье поведение проявлялось без приобретения предварительного опыта, по известному в этологии (см: Зорина и др., 2013) принципу “все и сразу” (Langley, 1986), тогда как у эврифагов оленьего и белоногого хомячков взаимодействие с добычей требовало накопления опыта (Kreiter, Timberlake, 1988) в той или иной мере. На основе частоты атак на живую добычу (сверчков) и модель (движущийся шарик) несколько видов ранжировали по степени “хищности” (Timberlake, Washburne, 1989). Этот показатель уменьшался от эврифагов (белоногого и оленьего хомячков) к видам, тяготеющим к питанию растительной пищей: калифорнийскому хомячку (P. californicus Gambel 1848), когтистой песчанке (Meriones unguiculatus Milne-Edwards 1867) и каирской мыши (Acomys cahirinus Geoffroy 1803); у щетинистого хлопкового хомяка (Sigmodon hispidus Say & Ord 1825) охотничье поведение не выявлено.
В последние годы у северных O. leucogaster и южных O. torridus кузнечиковых хомячков выявлены морфологические и физиологические адаптации к взаимодействию с относительно крупной и подвижной добычей, даже с ядовитой (скорпионы), что позволяет считать их специализированными хищниками (Rowe, Rowe, 2008; Sarko et al., 2011; Rowe et al., 2013). Охотничье поведение было недавно описано у целого ряда видов с различными пищевыми предпочтениями. Так, у рыжей полевки (Myodes glareolus Schreber 1780), вида со смешанной диетой, обнаружено охотничье поведение по отношению к сверчкам (Sadowska et al., 2008; Konczal et al., 2016). Детальные лабораторные эксперименты впервые выявили у зерноядного вида – полевой мыши (Apodemus agrarius Pallas 1771) – эффективное охотничье поведение по отношению к насекомым (в том числе, таким агрессивным и опасным как рыжие лесные муравьи Formica polyctena Forster 1850), в котором последовательность действий организована в стереотип с постоянными составляющими (Panteleeva et al., 2013; Reznikova et al., 2017). По тем же схемам нами были выявлены и описаны стереотипы хищнического поведения у зерноядной малой лесной мыши (Sylvaemus uralensis Pallas 1811) и даже у зеленоядных видов: узкочерепной (Lasiopodomys gregalis Pallas 1779), плоскочерепной (Alticola strelzovi Kastschenko 1899) и тувинской (A. tuvinicus Ognev 1950) полевок (Левенец и др., 2016, 2016а; Левенец, 2017).
Использование в качестве добычи подвижных и безопасных насекомых стандартных размеров (тараканы) послужило удачной поведенческой моделью. Оказалось, что разные виды грызунов применяют различные приемы для быстрого и эффективного захвата подвижной добычи, и при этом все они демонстрируют структурированные охотничьи стереотипы, во многом сходные с таковыми специализированных хищников, таких как насекомоядный вид – обыкновенная бурозубка (Sorex araneus Linnaeus 1758) (Левенец и др., 2016, 2016а; Левенец, 2017). По-видимому, стереотип охотничьего поведения распространен у грызунов достаточно широко. Это позволяет поставить вопрос об адаптивном значении и эволюции хищнического поведения в этой разнообразной таксономической группе.
В данной работе мы делаем попытку оценить роль филогенетической и экологической составляющих в становлении хищничества у грызунов на примере сравнения охотничьего поведения двух пар видов хомячков, которые принадлежат к родам с разным уровнем дивергенции и которые существенно различаются по морфологии, экологическим особенностям и склонности к охоте на подвижную добычу. В качестве объектов исследования выбраны два представителя рода Allocricetulus – хомячок Эверсманна (A. eversmanni) и монгольский хомячок (A. curtatus) – и два вида рода Phodopus – хомячок Кэмпбелла (P. campbelli) и джунгарский хомячок (P. sungorus). Охотничье поведение этих видов ранее экспериментально не изучалось.
Среди достаточно широкого набора жизненных форм палеарктических видов хомяков и хомячков подсемейства Cricetinae род Allocricetulus (эверсманновы хомячки, см.: Павлинов, 2006) обращает на себя внимание своим “хищным” обликом и поведением: прогонистое тело, выраженный шейный перехват (рис. 1), а также резкие порывистые движения. Ранее в экспериментах с парным ссаживанием зверьков на нейтральной территории было обнаружено, что хомячки Эверсманна (реже – монгольские хомячки) – ведут себя по отношению к конспецифику, скорее, как хищник к жертве, чем как к конкуренту. В течение нескольких секунд происходит схватка и, если экспериментатор не успеет разнять противников, укус одного из них в шейный отдел приводит к немедленному параличу (Рюриков и др., 2003; Ryurikov et al., 2004). В желудках хомячка Эверсманна, наряду с семенами растений, обнаруживали остатки беспозвоночных животных и мелких позвоночных, в том числе амфибий, рептилий, млекопитающих (Попов, 1960; Крыльцов, Шубин, 1964; Воронцов, 1982). В Кустанайской обл. этот вид поедает чернотелок, листоедов и жужелиц, и они нередко составляют основу его рациона (Алимбаев, 1965), а на зиму он заготавливает как семена, так и насекомых (Крыльцов, Шубин, 1964; Алимбаев, 1968). По нашим наблюдениям в Саратовской обл., хомячки Эверсманна запасают насекомых, а также поедают трупы лягушек и ящериц. В их норах встречались фрагменты мышей и полевок, в основном детенышей. У монгольского хомячка в степях Тувы и Монголии, по данным Флинта и Головкина (1961), доля животных кормов в желудках достигала 95%, а по результатам анализа содержимого защечных мешков он оказался наиболее “насекомоядным” из пяти исследованных там видов.

Хомячки рода Phodopus (Miller 1910) (мохноногие хомячки) известны как зерноядные формы, которые регулярно потребляют беспозвоночных животных, хотя и не отличаются столь “хищными” повадками, как эверсманновы хомячки (Феоктистова, 2008). Мы наблюдали за хомячком Кэмпбелла (P. campbelli) в Туве в окрестностях оз. Тере-Холь и в Монголии вблизи сомона Баян-Унжул в зоне сухих степей. Отмечали активную охоту этого вида на порхающих около кустов бабочек, сидящих на листве прямокрылых, а также раскапывание и поедание червеобразных личинок жуков. Стоит отметить, что жуки и саранчовые, обнаруженные вблизи или внутри нор этого вида, почти всегда были лишены головы и конечностей.
В целом, на основании весьма отрывочных литературных данных и наших предварительных наблюдений, мы можем считать эверсманновых хомячков более плотоядными видами, чем мохноногих.
Род Phodopus является самой древней группой подсемейства (время обособления от основного ствола оценено как 8.5–12.2 млн. лет). Генетическая дистанция между хомячками Кэмпбелла и джунгарским для последовательности гена cyt b составляет 4.5%, и время их разделения оценивается в 0.8–1 млн лет, а обособление рода Allocricetulus произошло в раннем Плиоцене (около 3.6–5.3 млн лет) (Neumann et al., 2006). Разделение рода на A. eversmanni и A. curtatus по тому же гену составляет 2.4%, что вдвое меньше, чем для пары видов родов Phodopus и составляет около 0.3 млн лет. Сходные датировки получены и по ядерным генам (Lebedev et al., 2018).
В лабораторных экспериментах мы детально проанализировали охотничье поведение четырех видов хомячков по отношению к подвижным насекомым. Основными задачами исследования были: сравнение различий в тактике, скорости и успешности охоты, оценка роли индивидуального опыта, анализ структуры последовательностей поведенческих актов при взаимодействии с добычей и выявление стереотипизации хищнического поведения.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования эверсманновых хомячков A. eversmanni (1 самец и 7 самок) и A. curtatus (5 самцов и 8 самок) проводили в 2017 г. на Научно-экспериментальной базе “Черноголовка” ИПЭЭ РАН с использованием животных, входящих в ЦКП “Живая коллекция диких видов млекопитающих”. Аутбредных хомячков Кэмпбелла (11 самцов и 8 самок) и джунгарских хомячков (5 самцов 7 самок) исследовали в виварии ИCиЭЖ СО РАН в 2016 г. Все животные были половозрелыми, возраст до двух лет, рождены в вивариях. До тестирования они не встречались с потенциальной добычей и не имели опыта охоты. Зверьков содержали в индивидуальных клетках 26 × 21 × 14 см при постоянном доступе к воде и пище. В рацион входили стандартные гранулы для лабораторных животных, зерносмесь (семена подсолнечника, овса), свежие овощи, крупяная каша, приготовленная на мясном бульоне, вареные яйца и творог. Все эксперименты проведены с соблюдением правил научных исследований с использованием экспериментальных животных, утвержденных распоряжением Президиума АН СССР от 2 апреля 1980 12000-№ 496 и приказом Минвуза СССР от 13 сентября 1984 № 22.
Для исследования спонтанного проявления охотничьего поведения, животным предлагали подвижную добычу – имаго и нимф старших возрастов мраморного таракана (Nauphoeta cinerea Olivier 1789), со средней длиной тела 27.93 ± 0.22 мм. Хомячков Эверсманна, монгольских и джунгарских хомячков тестировали по 3 раза, а хомячков Кэмпбелла – по 7. Для наблюдения за процессом охоты зверьков по одному помещали в прозрачную стеклянную арену размером 40 × 25 × 30 см для эверсманновых хомячков и 30 × 30 × 35 см для мохноногих хомячков. Передерживали на арене в течение 5 мин, после чего предъявляли добычу (аккуратно сбрасывали на арену сверху). Действия животных фиксировались с помощью видеокамеры Sony HDR–AS200V (60 кадров/с). Если животное охотилось, то на арену помещали еще две единицы добычи, каждую после поедания предыдущей. Если хомячок не проявлял интерес к насекомому в течение 10 мин, тест прекращали. Всего проведено 226 тестов.
Для описания поведения в качестве элементарной единицы выделялись элементарные двигательные акты и позы (Tinbergen, 1951) – для краткости, “элементы поведения”. Поведенческой последовательностью обозначали набор последовательно совершаемых элементов поведения. Внутри поведенческих последовательностей мы выделяли поведенческие стереотипы, состоящие из устойчиво повторяющихся “цепочек” элементов поведения (подробно см.: Reznikova et al., 2017). Обработка видеоматериала проводилась при 5-и и 25-и кратном замедлении в программе Noldus Observer XT 12.5. Для анализа стереотипов отбирались только случаи успешной охоты, закончившейся поимкой и поеданием добычи. Используя полученный “алфавит” из 17 элементов поведения, мы преобразовали демонстрируемое поведение в последовательности букв, где каждая буква соответствовала одному элементу поведения. Полученные последовательности символов через пробел перемещались в отдельные для каждого вида “суммарные” текстовые файлы (в формате .txt). Каждый суммарный текстовый файл включал все зафиксированные поведенческие последовательности (т.е. при предъявлении более одной единицы добычи за тест), полученные при первичном и повторных тестированиях одних тех же особей. Для построения схемы стереотипа на основе “суммарных” текстовых файлов рассчитывались матрицы вероятностей перехода от одного поведенческого элемента к другому (Марковский процесс первого порядка) (Casarrubea et al., 2008).
Доли успешных и неуспешных атак попарно сравнивали с помощью точного теста Фишера с поправкой Бонферрони. Сравнение количества элементов в стереотипах и “скорости охоты” (количества элементов поведения в единицу времени), проведен с помощью H-критерия Краскела–Уоллиса с поправкой Бонферрони. При сравнении представлены медиана, первый и третий квартили: Me (Q1–Q3).
РЕЗУЛЬТАТЫ
Элементы охотничьего поведения у всех четырех видов оказались одинаковыми. Мы разделили 17 выделенных элементов поведения на 3 типа. 1) “Ключевые”, связанные с обнаружением, преследованием и захватом добычи, без которых совершение стереотипа невозможно: сближение с добычей бегом (Q) или спокойным шагом (S), укус (W), захват добычи лапами (E). 2) “Дополнительные”, связанные с “приготовлениями” к охоте и с “обработкой” добычи и присутствующие не во всех стереотипах: принюхивание (D), перенос добычи в зубах (G) (у джунгарского хомячка не обнаружен), перехват (R), откусывание конечностей насекомого (H). 3) “Шумовые”, не связанные непосредственно с процессом охоты и не влияющие на совершение стереотипа: замирание (C), поворот корпуса на 90° (V), разворот корпуса 180° (B), поворот головы (F), вертикальная стойка (I), стойка с опорой на арену (Y), движения назад (U), прыжок (J) и чистка (X). Отметим, что у джунгарского хомячка из “шумовых” элементов обнаружены лишь три: “поворот корпуса на 90°”, “разворот корпуса 180°” и “движение назад”.
У обоих видов рода Phodopus охотничий стереотип проявлялся факультативно. В опытах с хомячками Кэмпбелла при первом предъявлении добычи 7 из 19 (36.8%) зверьков проявили охотничье поведение, после семи тестов количество охотящихся животных возросло до 12 (63.1%), а 7 ни разу не приступили к охоте. Все джунгарские хомячки (n = 12) при первом предъявлении добычи не охотились, и только 3 проявили охотничье поведение при повторных тестах. У хомячков Эверсманна и монгольских хомячков охотничий стереотип проявлялся у всех животных при первом же тестировании.
Не все акты охоты заканчивались успешной поимкой добычи. Мы оценивали успешность охоты как соотношение атак, закончившихся поимкой добычи и атак, в которых добыча была упущена. Наиболее успешным охотником на насекомых из всех исследованных видов оказался монгольский хомячок: 80% (39 из 49) случаев охоты в первом же тесте закончились поимкой и поеданием таракана. Это достоверно отличается от успешности охоты хомячков Эверсманна (46%, n = 48) и Кэмпбелла (35%, n = 20) (точный тест Фишера с учетом поправки Бонферрони, p < < 0.0017 для обоих случаев). По итогам повторных тестов успешность охоты у эверсманновых хомячков не изменилась, а у хомячков Кэмпбелла проявилась тенденция к ее увеличению (до 51%). У трех джунгарских хомячков, которые начали охотиться в повторных тестах, доля успешных случаев охоты составила 76% (16 из 21).
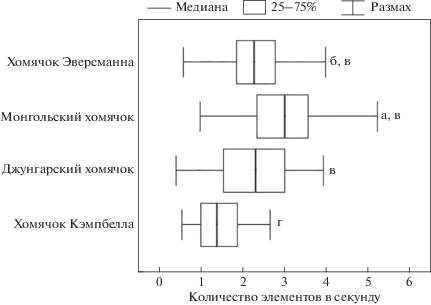
На протяжении всех тестов во время актов успешной охоты наиболее стремительно охотились монгольские хомячки, совершая 3 (2.4–3.6) элемента поведения в секунду (рис. 2). Стоит отметить, что в охотничьих стереотипах монгольского хомячка доля “шумовых” элементов самая низкая (3%, 81 из 2700) по сравнению c хомячком Эверсманна (5.3%, 74 из 1404), Кэмпбелла (4.8%, 80 из 1674) и джунгарским хомячком (6.9%, 18 из 262), то есть они меньше “отвлекаются” во время охоты (точный тест Фишера с поправкой Бонферрони, p < 0.0033 для всех случаев). Несколько ниже скорость охоты у хомячка Эверсманна – 2.3 (1.9–2.8), наименьшая – у хомячков Кэмпбелла – 1.4 (1–1.9). Значения критерия Краскела-Уоллиса в парах составляли: для монгольских и хомячков Эверсманна (H = 20, p < 0.0033), монгольских и джунгарских хомячков, монгольских хомячков и хомячков Кэмпбелла (H = 63, p < 0.0033), для хомячков Эверсманна и Кэмпбелла (H = 27, p < < 0.0033). Скорость охоты у джунгарского хомячка 2.3 (1.6–2.9) не отличалась от таковой у монгольского и хомячка Эверсманна и была выше, чем у хомячков Кэмпбелла (H = 8.4, p < 0.0083).
Рис. 2.
Скорость охоты: количество элементов поведения, совершаемых в секунду. Данные, отмеченные буквами а, б, в и г, достоверно различаются (H-тест с поправкой Бонферрони, p < 0.0033).
Для того чтобы составить представление о том, как разные виды хомячков атакуют добычу и как манипулируют ею, мы сравнили количество элементов поведения, связанных с атакой и обработкой добычи, которые приходятся на один стереотип (табл. 1). Количество элементов “укус” у хомячка Кэмпбелла больше, чем у монгольского (H = 9.3, p < 0.0083) и не отличается от количества “укусов” у хомячка Эверсманна. Джунгарский хомячок ни от одного из видов по этому показателю достоверно не отличался. Количество элементов “захват добычи лапами”, приходящихся на один стереотип, у хомячка Эверсманна, монгольского и джунгарского не различалось, а у хомячка Кэмпбелла этот показатель был достоверно выше, чем у всех остальных (H = 13, p < 0.0016; H = 9.4, p < 0.0016; H = 8.2, p < 0.0083 соответственно). Хомячки Кэмпбелла и джунгарские совершали перехваты добычи реже, чем хомячки Эверсманна и монгольские (p < 0.0016 в обоих случаях), и при этом хомячки Кэмпбелла откусывали конечности добычи чаще, чем особи всех остальных видов: хомячков Эверсманна (H = 33, p < 0.0016), монгольских (H = 54, p < 0.0016) и джунгарских (H = 24, p < 0.0016).
Таблица 1.
Количество элементов поведения и переходов между ними, приходящихся на один охотничий стереотип, у исследованных видов хомячков
| Элементы поведения и переходы между ними | Хомячок Эверсманна | Монгольский хомячок | Хомячок Кэмпбелла | Джунгарский хомячок |
|---|---|---|---|---|
| Укус | 5.5 (3–9)а, б | 6 (3–8)а | 8 (4.5–14)б | 5 (3–6)а, б |
| Захват лапами | 6 (3–9)а | 6 (3–11)а | 10 (5.5–18)б | 5 (4–6.8)а |
| Перехват | 5 (3–7)а | 6 (4–8)а | 1 (0–2)б | 1 (0.8–2)б |
| Откусывание конечностей | 1 (0–2)а | 1 (0–1)а | 3 (2–6)б | 1 (0–1)а |
| Укус – захват лапами | 4 (2–7) | 4 (2–6) | 6 (2.5–11) | 3 (1–4) |
| Захват лапами – укус | 2 (1–3.2) | 2 (1–4) | 2 (1–5) | 1 (0–1.3) |
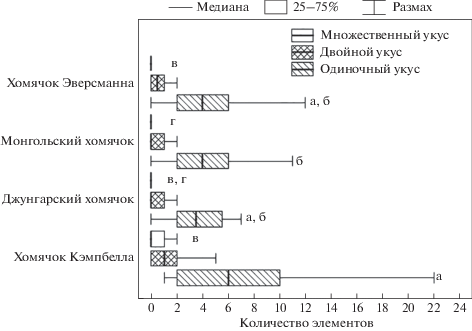
Для того чтобы понять, каким образом охотник повреждает добычу, мы исследовали повторяемость элемента поведения “укус” (W) (рис. 3). Вся совокупность этих элементов была разделена на три группы: одиночные, двойные (два последовательно совершаемых “укуса”) и множественные (три и более “укусов” подряд). Для всех исследованных видов характерны были, главным образом, одиночные укусы. Количество одиночных укусов в охотничьих стереотипах у хомячка Кэмпбелла было выше, чем у монгольского (H = 8.3, p < 0.0083). По содержанию двойных укусов охотничьи стереотипы у всех исследованных видов хомячков не различались. Количество множественных укусов у монгольских хомячков было меньше, чем у хомячков Эверсманна (H = 7.4, p < 0.0083), Кэмпбелла (H = 8.6, p < 0.0083) и не отличалось от этого же показателя у джунгарских хомячков.
Рис. 3.
Количество одиночных, двойных и множественных укусов, приходящихся на один охотничий стереотип, у разных видов. Данные, отмеченные буквами (одиночные укусы: а, б; множественные укусы: в, г), достоверно различаются (H-тест с поправкой Бонферрони, p < 0.013).
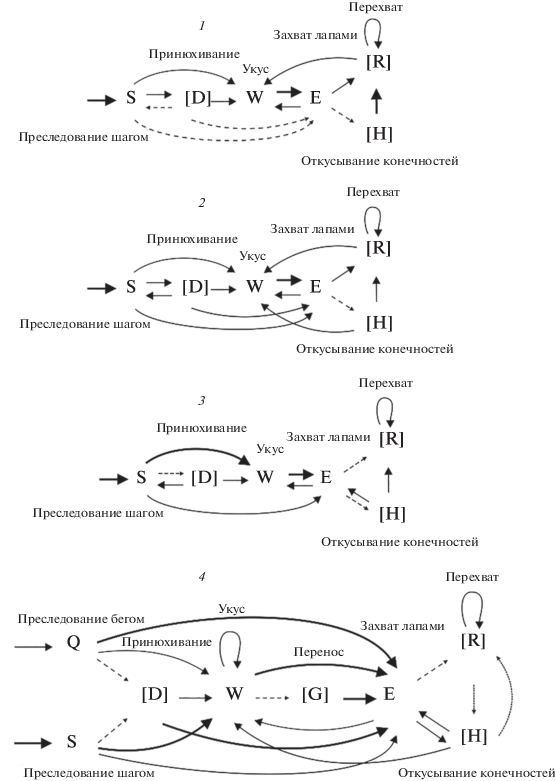
На основании матриц вероятностей переходов между элементами поведения построены схемы охотничьих стереотипов (рис. 4). У всех четырех видов охотничий стереотип начинался со сближения с добычей бегом (Q) или спокойным шагом (S), затем могло следовать принюхивание (D). Атака с большей вероятностью начиналась с захвата зубами (“укус”, W), с меньшей – с захвата лапами (E). Количество переходов от “укуса” к “захвату лапами” у всех видов было достоверно выше, чем “захвата лапами” к “укусу” (Н-тест, p < 0.0016 для всех случаев) (см. табл. 1). После поимки добычи грызуны могли переносить ее в зубах (G) или начинали ею манипулировать. Если атака начиналась с “укуса” (W), то в дальнейшем грызуны захватывали насекомое лапами (Е), и после этого чаще всего следовали перехваты (R), которые включали манипуляции удерживаемой в лапах добычей (повороты, перевороты). У хомячков Кэмпбелла манипуляции лапами с добычей наблюдались реже, чем у других видов, зато они значительно чаще откусывали конечности насекомых (H). У всех видов стереотипы охоты могли оканчиваться “укусом” (W), “захватом лапами” (E), “перехватом” (R) или “откусыванием конечностей добычи” (H). Далее следовало поедание добычи.
Рис. 4.
Схемы охотничьего стереотипа: 1 – хомячок Эверсманна, 2 – монгольский хомячок, 3 – джунгарский хомячок, 4 – хомячок Кэмпбелла. На схемах тонкой пунктирной линией обозначены некоторые неустойчивые связи между элементами (p < 0.2). Простой черной линией обозначены устойчивые связи (0.2 ≤ p < 0.5). Жирной линией обозначены высоко устойчивые связи между элементами поведения (p ≥ 0.5). В скобках указаны дополнительные элементы поведения.
ОБСУЖДЕНИЕ
В наших экспериментах впервые выявлено и детально описано у четырех видов хомячков целенаправленное взаимодействие с подвижной добычей по типу “хищник–жертва”, с демонстрацией полного стереотипа охотничьего поведения. Набор поведенческих актов у них оказался одинаковым и в целом сходным с репертуаром ранее исследованных видов мышевидных грызунов (Левенец, 2017; Reznikova et al., 2017).
Обнаруженные нами различия в скорости, тактике и успешности охоты в целом отражают как видовую специфику питания хомячков, так и их филогенетический статус. Наиболее успешным охотником на насекомых оказался самый “насекомоядный” из исследуемых видов монгольский хомячок. В наших экспериментах с предъявлением таракана у хомячков Эверсманна и Кэмпбелла успешность охоты оказалась почти вдвое меньше, чем у монгольского. Возможно, хомячок Эверсманна проявил бы наибольшую сноровку в охоте на позвоночных.
Тактику охоты двух видов эверсманновых хомячков можно описать как быстрый захват и повреждение добычи одним или несколькими точными укусами. Высокая эффективность укусов достигается за счет манипуляций с добычей лапами. Тактика охоты хомячка Кэмпбелла отличается откусыванием конечностей для обездвиживания добычи. Это выражается в устойчивости связей между элементами поведения “захват лапами” и “откусывание конечностей” (0.2 ≤ p < 0.5). У джунгарского хомячка паттерн самый простой: догнать, схватить и сразу начать есть, насколько это удается почти без манипуляций.
Для обоих видов эверсманновых хомячков (более плотоядных) отмечена высокая манипуляторная активность, что отражается в повторяющихся цепочках поведенческих элементов “перехват добычи – укус”. Поворачивая добычу лапами, хомячки наносят ей один, реже несколько, точных укусов. Манипулирование добычей с помощью передних конечностей было выявлено ранее у серых крыс (Дерягина, 1980; Махмутова, 1983) и полевых мышей (Пантелеева и др., 2011). У эверсманновых хомячков манипуляторное поведение значительно более направлено. В отличие от крыс, которые кусают много и беспорядочно (Левенец и др., 2016а), хомячки, перехватывая лапами, подставляют насекомое под точные укусы. Менее хищные хомячки Кэмпбелла действуют иначе: захватив насекомое в передние лапы, они обездвиживают его, удерживая в лапах и откусывая конечности. Интересно отметить, что и в естественных условиях прямокрылые и жуки, найденные нами вблизи нор хомячков Кэмпбелла, были лишены конечностей и голов. Джунгарские хомячки добычей почти не манипулируют.
Для формирования представлений об эволюции охотничьего поведения у исследованных видов важно проанализировать начало атаки на добычу. На примере специализированных хищных кузнечиковых хомячков было показано, что начало атаки с захвата добычи лапами является одним из показателей хищнической специализации и, соответственно, эволюционной продвинутости охотничьего поведения (Langley, 1994). Неспециализированные охотники – сирийские хомяки (M. auratus) (Polsky, 1977; Langley, 1987), калифорнийские хомячки (P. сalifornicus Gambel 1848) и каирские мыши (A. cahirinus Geoffroy 1803) (Timberlake, Washburne, 1989) в большинстве случаев начинают атаку с захвата добычи зубами, что считается более примитивным признаком в филогенетическом плане (Eisenberg, Leyhausen, 1972; Langley, 1994). У всех исследованных нами видов хомячков атака на добычу начиналась с захвата лапами (E) реже, чем атака с захвата зубами (W). Вероятность перехода “W–E” у них около 50%, а “E–W” – около 25%. Тем не менее, мы считаем, что и 25% – это весьма устойчивое поведение. Стоит отметить, что и у кузнечиковых хомячков начало охоты с захвата добычи лапами происходит не всегда. Проанализировав видео охоты южного кузнечикового хомячка, полученное в исследованиях Rowe с соавторами (Rowe et al., 2013) (URL: https://youtu.be/s-YZwYHrD24), мы выявили один случай начала атаки с захвата зубами из семи. Таким образом, охотничьи стереотипы у всех исследованных нами видов и у специализированных хищных хомячков рода Onychomys обладают чертами сходства. Можно высказать предположение, что паттерн охоты джунгарского хомячка близок к “исходному”: он самый простой и лишен таких черт специализации, как специфические манипуляции добычей, наблюдавшиеся нами в разных формах у других видов.
В целом же схемы стереотипов охоты на подвижных насекомых оказались у всех четырех видов весьма сходными по порядку ключевых элементов, а существенно различаются у них такие показатели, как частота проявления охотничьего поведения в популяциях (в той мере, в какой этот показатель выражен у животных, рожденных в вивариях), эффективность охоты и роль индивидуального опыта в формировании стереотипа. У более плотоядных эверсманновых хомячков охотничье поведение проявляется облигатно, как и у хищных хомячков рода Onychomys (Langley, 1987), а у мохноногих хомячков – факультативно. Нам кажется особенно интересным, что эффективность охотничьего стереотипа не зависит от частоты его проявления в популяциях разных видов. Так, у джунгарских хомячков самая низкая доля “охотников”, однако паттерн у них полный, а успешность и скорость охоты сходны с соответствующими показателями у монгольского хомячка – лучшего охотника на насекомых из исследованных нами видов. Это говорит о цельности стереотипа охотничьего поведения у разных видов хомячков. Характерно, что у более “хищных” эверсманновых хомячков стереотип охотничьего поведения проявляется по принципу “все и сразу”, как и у специализированных кузнечиковых хомячков (Langley, 1986), а менее плотоядные мохноногие хомячки, по-видимому, нуждаются в некотором опыте для совершенствования охотничьего стереотипа, как это было показано ранее на сирийских хомяках (Polsky, 1977) и белоногих хомячках (Kreiter, Timberlake, 1988).
В целом, выявленное нами хищническое поведение по отношению к насекомым отражает видовую специфику питания разных видов хомячков, а паттерны атаки на подвижную добычу у них универсальны и сравнимы с таковыми у специализированных хищных видов. Самый простой стереотип охотничьего поведения выявлен у джунгарского хомячка, у остальных видов отмечаются черты специализации в виде манипуляторной активности разных форм. Особенно оригинальную форму принимает эта активность у хомячка Кэмпбелла. Наиболее стремительным и эффективным охотником оказался монгольский хомячок. Облигатное и полное (не нуждающееся в предварительном опыте) проявление паттерна у двух видов эверсманновых хомячков может говорить о большей специализации хищнического поведения у этой эволюционно более молодой группы, по сравнению с мохноногими хомячками, у которых атаки на добычу проявляются факультативно и совершенствуются с приобретением опыта.
Список литературы
Алимбаев Р.А., 1965. К экологии хомячка Эверсманна в Кустанайской области // Вестник сельскохозяйственной науки. № 7. С. 46–52.
Алимбаев Р.А., 1968. Влияние распашки целинных и залежных земель на распределение и экологи сусликов, хомяков и тушканчиков Кустанайской области. Автореф. дис. … канд. биол. наук. Алма-Ата. 18 с.
Башенина Н.В., 1977. Пути адаптации мышевидных грызунов. М.: Наука. 354 с.
Воронцов Н.Н., 1982. Фауна СССР. Млекопитающие. Низшие хомякообразные (Cricetidae) мировой фауны. Л.: Наука. 449 с.
Дерягина М.А., 1980. Особенности манипуляционной активности грызунов // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 85. Вып. 4. С. 11–19.
Зорина З.А., Полетаева И.И., Резникова Ж.И., 2013. Основы этологии и генетики поведения: Учебник. 3-е изд., испр. и доп. М.: Изд-во МГУ. 384 с.
Крыльцов А.И., Шубин И.Г., 1964. Об экологии хомячков Эверсманна (Cricetulus eversmanni) и джунгарского хомячка (Phodopus sungorus) // Зоологический журнал. Т. 43. № 7. С. 29–36.
Левенец Я.В., 2017. Оценка изменчивости и сложности поведенческих стереотипов на примере охотничьего поведения мелких млекопитающих. Автореф. дис. … канд. биол. наук. Новосибирск: Институт систематики и экологии животных СО РАН. 22 с.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., 2016. Сравнительный анализ стереотипного поведения на примере процесса охоты у мелких млекопитающих // Экспериментальная психология. Т. 9. № 4. С. 68–78.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., 2016а. Экспериментальное исследование питания насекомыми у грызунов // Евразиатский энтомологический журнал. Т. 6. № 6. С. 550–554.
Махмутова Е.Н., 1983. Анализ манипуляционной активности животных. Автореф. дис. … канд. психол. наук. М. 16 с.
Павлинов И.Я., 2006. Систематика современных млекопитающих. М.: Изд-во МГУ. 287 с.
Пантелеева С.Н., Выгоняйлова О.Б., Резникова Ж.И., 2011. Рыжие лесные муравьи как потенциальная массовая добыча полевых мышей: результаты лабораторных экспериментов // Евразиатский энтомологический журнал. Т. 10. С. 99–103.
Попов В.А., 1960. Млекопитающие Волжско-Камского края: насекомоядные, рукокрылые, грызуны. Казань: Изд-во АН СССР. 68 с.
Резникова Ж. И., 2016. Экология, этология, эволюция. Межвидовые отношения животных в 2-х частях. Учебник для вузов. М.: Изд-во “Юрайт”. 262 с.
Рюриков Г.Б., Суров А.В., Тихонов И.А., 2003. Хомячок Эверсманна (Allocricetulus eversmanni) в Саратовском Заволжье: экология и поведение в природе // Поволжский экологический журнал. № 3. С. 251–258.
Феоктистова Н.Ю., 2008. Хомячки рода Phodopus. Систематика, филогеография, физиология, поведение, химическая коммуникация. М.: Товарищество научных изданий КМК. 414 с.
Флинт В.Е., Головкин А.Н., 1961. Очерк сравнительной экологии хомячков Тувы // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 66. Вып. 5. С. 57–75.
Babinska-Werka J., Garbarczyk H., 1981. Animal components of the diet of the striped field mouse under urban conditions // Acta theriologica. V. 26. № 18. P. 301–318.
Caro T.M., 1980. The effects of experience on the predatory patterns of cats // Behavioral and Neural Biology. V. 29. № 1. P. 1–28.
Casarrubea M., Sorbera F., Crescimanno G., 2008. Multivariate analysis of the modifications induced by an environmental acoustic cue on rat exploratory behavior // Physiology & Behavior. V. 93. № 4. P. 687–696.
Eisenberg J.F., Leyhausen P., 1972. The phylogenesis of predatory behavior in mammals // Zeitschrift für Tierpsychologie. V. 30. № 1. P. 59–93.
Konczal M., Koteja P., Orlowska-Feuer P., Radwan J., Sadowska E.T., Babik W., 2016. Genomic response to selection for predatory behavior in a mammalian model of adaptive radiation // Molecular Biology and Evolution. V. 33. № 9. P. 2429–2440.
Kreiter N., Timberlake W., 1988. Form and development of predation on crickets in adults of Peromyscus maniculatus bairdi and P. leucopus noveboracensis // Journal of Comparative Psychology. V. 102. № 3. P. 269–278.
Landry S.O., 1970. The Rodentia as omnivores // Quarterly Review of Biology. V. 45. P. 351–372.
Langley W.M., 1986. Development of predatory behavioral in the southern grasshopper mouse (Onychomys torridus) // Behaviour. V. 99. P. 275–295.
Langley W.M., 1987. Specializations in the predatory behavior of grasshopper mice (Onychomys leucogaster and O. torridus): a comparison with the golden hamster (Mesocricetus auratus) // Journal of Comparative Physiology and Psychology. V. 101. P. 322–327.
Langley W.M., 1994. Comparison of predatory behaviors of deer mice (Peromyscus maniculatus) and grasshopper mice (Onychomys leucogaster) // Journal of Comparative Psychology. V. 108. № 4. P. 394–400.
Lebedev V.S., Bannikova A.A., Neumann K., Ushakova M.V., Ivanova N.V., Surov A.V., 2018. Molecular phylogenetics and taxonomy of dwarf hamsters Cricetulus Milne-Edwards, 1867 (Cricetidae, Rodentia): description of a new genus and reinstatement of another // Zootaxa. V. 4387. № 2. P. 331–349.
Neumann K., Michaux J., Lebedev V., Yigit N., Colak E., et al., 2006. Molecular phylogeny of the Cricetinae subfamily based on the mitochondrial cytochrome b and 12S rRNA genes and the nuclear vWF gene // Molecular Phylogenetics and Evolution. V. 39. № 1. P. 135–148.
Panteleeva S., Reznikova Z., Vygonyailova O., 2013. Quantity judgments in the context of risk/reward decision making in striped field mice: first “count,” then hunt // Frontiers in Psychology. V. 4. P. 45–53.
Polsky R.H., 1977. The ontogeny of predatory behaviour in the golden hamster (Mesocricetus a. auratus): I. The influence of age and experience // Behaviour. V. 61. P. 26–57.
Reznikova Z., Levenets J., Panteleeva S., Ryabko B., 2017. Studying hunting behaviour in the striped field mouse using data compression // Acta ethologica. V. 20(2). P. 165–173.
Rowe A.H., Rowe M.P., 2008. Physiological resistance of grasshopper mice (Onychomys spp.) to Arizona bark scorpion (Centruroides exilicauda) venom // Toxicon. V. 52. № 5. P. 597–605.
Rowe A.H., Xiao Y., Rowe M.P., Cummins T.R., Zakon H.H., 2013. Voltage-gated sodium channel in grasshopper mice defends against bark scorpion toxin // Science. V. 342. № 6157. P. 441–446.
Ryurikov G.B., Tolstov I.V., Surov A.V., 2004. Sexual and aggressive patterns in behavior of 7 hamster species in captivity // Advances in Ethology. V. 38. P. 77.
Sadowska E.T., Baliga-Klimczyk K., Chrzascik K.M., Koteja P., 2008. Laboratory model of adaptive radiation: a selection experiment in the bank vole // Physiological and Biochemical Zoology. V. 81. № 5. P. 627–640.
Sarko D.K., Leitch D.B., Girard I., Sikes R.S., Catania K.C., 2011. Organization of somatosensory cortex in the Northern grasshopper mouse (Onychomys leucogaster), a predatory rodent // Journal of Comparative Neurology. V. 519. № 1. P. 64–74.
Timberlake W., Washburne D.L., 1989. Feeding ecology and laboratory predatory behavior toward live and artificial moving prey in seven rodent species // Animal Learning & Behavior. V. 17. P. 2–11.
Tinbergen N., 1951. The Study of Instinct. London: Oxford University Press. 228 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал