

Зоологический журнал, 2020, T. 99, № 4, стр. 394-404
Новые данные о креветках семейства Crangonidae (Crustacea, Decapoda)
Н. А. Заренков *
Биологический факультет Московского государственного университета
им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: zarenkovn@yandex.ru
Поступила в редакцию 16.12.2018
После доработки 23.06.2019
Принята к публикации 26.06.2019
Аннотация
Сообщается о находках 9 видов креветок Crangonidae, уточняются их географическое распространение и батиметрия. Обсуждается комбинаторика структур сегмента VI абдомена и плеопода II самцов.
Морские креветки семейства Crangonidae характеризуются всесветным распространением и широкой эврибатностью (Bruce, 1988; Chan, 1996; Komai, 1995, 1997, 2001, 2008; Komai, Takeda, 1989; Kim, Hayashi, 2003; Komai, Amaoka, 1991; Komai, Chan, 2009).
Большинство видов семейства обитает на дне, на малых глубинах и только немногие встречаются на глубине больше 3 тыс. м, т.е. проникают в абиссаль: Neocrangon abyssorum (Rathbun 1902) (887‒3640 м), Sclerocrangon zenkevitchi Birstein et Vinogradov 1953 (2995‒4070 м), Parapontophilus occidentalis (Faxon 1893) (1760‒4018 м), P. talismani Crosnier et Forest 1973 (2100‒3459 м), а также группа видов, близких к P. gracilis (Smith 1882) (1800‒5852 м). Этот список может быть пополнен находками Рlacopsicrangon formosa Komai et Chan 2009 (4412‒4824 м) (Komai, Chan, 2009).
При построении системы семейства, выполненном Кристоферсеном (Christoffersen, 1988) с помощью кладистического метода, использованы 74 признака (“characters”). Предложенная им система состоит из 5 подсемейств и отличается большой информативностью.
Тем не менее, ревизия системы семейства остается актуальной, потому что в последние десятилетия описаны десятки новых видов и уточнены диагнозы уже известных видов. Некоторые виды семейства известны по немногим находкам, и их географическое распространение нуждается в уточнении. В связи с этим могут представить интерес публикуемые здесь новые находки уже известных видов.
Изученные автором коллекции (Заренков, 1965), включая вновь публикуемые материалы, позволяют пополнить и детализировать информацию, которая заключена в таксономических диагнозах. В настоящей публикации с этой целью систематизированы доступные автору данные о строении сегмента VI абдомена и передних плеоподов самца. У разных видов эти придатки нередко различаются относительными размерами эндо- и экзоподита, несущих оперенные щетинки, и appendix masculina, вооруженного шипами. Показано, что строение плеоподов самца осуществилось по меньшей мере в двух вариантах, тогда как строение VI сегмента абдомена представлено не менее чем тремя морфотипами. При этом таксономическое разнообразие остается комбинацией структур, подобных “characters” Кристоферсена (Christoffersen, 1988).
Благодаря любезности проф. Чейса (Chace), автор имел возможность ознакомиться с материалом по Crangon (Neocrangon) resima Rathbun 1902, определение Шмидта (Schmitt). Длина креветок измерялась от края орбиты до середины заднего края карапакса. Изученная коллекция хранится в Зоологическом музее Московского университета им. М.В. Ломоносова.
НОВЫЕ НАХОДКИ КРЕВЕТОК СЕМЕЙСТВА CRANGONIDAE
Placopsicrangon formosa Komai et Chan 2009
Материал. “Витязь”. Ст. 4320, 10.02.1959, 20°01′9 с.ш., 161°03′0 з.д., глубина 4718 м, самец дефектный 18.0 мм.
Замечания. Вновь обнаруженный экземпляр этого вида удовлетворительно отвечает первоописанию (Komai, Chan, 2009).
Важной особенностью P. formosa служит латерально уплощенный ножевидный рострум. У паратипов он заострен и вооружен зубцами числом 2–5. У нашего экземпляра конец рострума притуплен и имеется единственный зубец.
Короткий, уплощенный c боков рострум отличает Рlacopsicrangon от других представителей семейства, которые имеют длинный вздернутый вверх рострум, как например, у Sclerocrangon zenkevitchi или S. unidentata Komai et Takeda 1989, а также отличает от Vercoia Baker 1904 и Sabinea Ross 1835. Два последних рода имеют широкий рострум, уплощенный в спинно-брюшном направлении (Duris, 1992).
Отсутствием клешни на переоподе II вновь обнаруженный Рlacopsicrangon сходен с родами Sabinea Owen и Vercoia Baker. Общей особенностью Рlacopsicrangon и Sabinea является слабое развитие переоподов II. Далее, только у Рlacopsicrangon глаза лишены стебельчатой части и превратились в массивные придатки, полностью заполняющие орбиты.
Распространение. Наша находка в Центральной Пацифике у Гавайских о-вов, глубина 4718 м, значительно расширяет ареал вида, который был известен только восточнее Тайваня (Формоза) на глубине 4412‒4824 м (Komai, Chan, 2009).
Prionocrangon curvicaulis Yaldwyn 1960 (рис. 1a‒1f)
Рис. 1.
Prionocrangon curvicaulis Yaldwyn, самец 8.2 мм: a ‒ основание антенны II, b ‒ тельсон и правый уропод, c ‒ переопод I, d ‒ переопод II, e ‒ переопод III, f ‒ переопод IV.
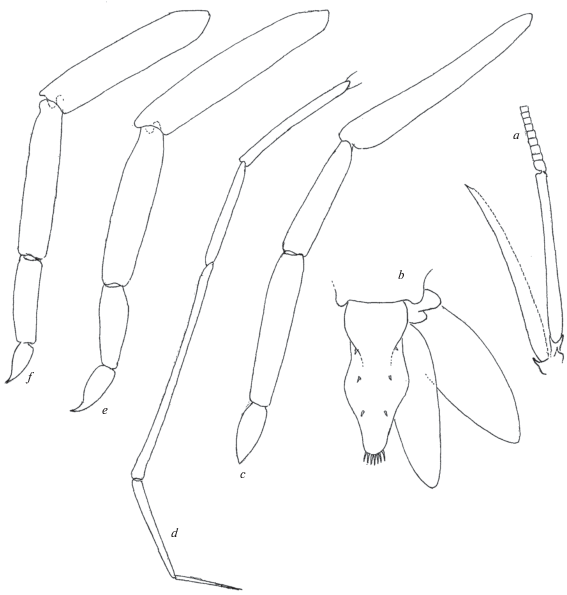
Материал. “Дмитрий Менделеев”. Ст. 1277, 15.01.1976. 51°00′8 ю.ш., 169°59′5 в.д., глубина 570 м, 2 самца 7.3, 8.2 мм.
Замечания. Нижеследующие уточнения и дополнения к первоописанию, тем не менее, не служат поводом для сомнений в принадлежности наших экземпляров к данному виду.
Cкафоцерит изогнут, узкий, саблевидный. На срединном гребне карапакса насчитывается 8‒10 шипов (в первоописании 7‒11). Дистальный конец уроподов плавно закруглен, тогда как согласно первоописанию скорее заострен. На переоподах II, IV, V имеются оперенные щетинки. Пальцы переоподов II широко ланцетовидные, пальцы переоподов IV, V широкие когтевидные. Переоподы III тонкие, длиннее прочих, оканчиваются длинным тонким пальцем. Тельсон с поперечным пережимом посредине.
Распространение. Подводное поднятие Чатам, Новая Зеландия и воды о-ва Самар, Филиппины, глубина 403‒700 м (Yaldwyn, 1960; Chace, 1984). Новая находка – в Большом Австралийском заливе, глубина 570 м.
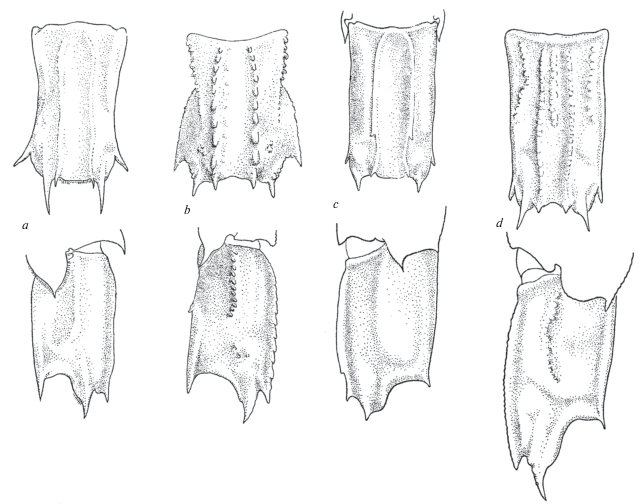
Neocrangon abyssorum Rathbun 1902 (рис. 2a)
Рис. 2.
Строение сегмента VI абдомена со спины и сбоку: a ‒ Neocrangon abyssorum (Rathbun), b ‒ Меsoсrangon intermedia (Stimpson), c ‒ Neocrangon communis (Rathbun).
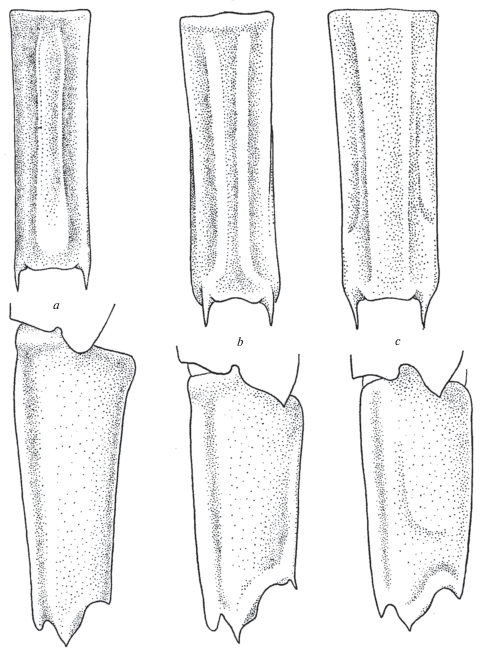
Материал. “Витязь”. Ст. 3353, 05.06.1955, 53°53'5 с.ш., 169°15'5 в.д., глубина 1600 м, самка 9.8 мм, самец 7.5 мм. Ст. 7502, 22.06.1976, 37°36′15 с.ш., 143°03′2 в.д., глубина 3620‒3640 м, 2 самки 6.5, 6.5 мм, 2 экз. дефектные 6.5, 6.7 мм.
Замечания. Уже опубликованные рисунки этого вида (Komai, 1991) представляется целесообразным дополнить изображением сегмента VI абдомена. Он имеет цилиндрическую форму, несет пару низких дорсальных килей, которые не достигают заднего края сегмента. Последний представляет собой неглубокую выемку и с боков ограничен парой заостренных выступов. Сходное строение сегмента VI наблюдается у Neocrangоn communis Rathbun 1899 и Mesocrangon intermedia (Stimpson 1860) (рис. 2b‒2c).
Распространение. В настоящее время уже известно не менее 29 находок этого преимущественно батиального вида (Бирштейн, Виноградов, 1951; 1953; Бирштейн, Заренков, 1970; Rathbun, 1902; 1904; Schmitt, 1921; Komai, 1991), с учетом вновь публикуемых. Они позволяют очертить ареал вида следующим образом: от тихоокеанских вод островов Хонсю и Хоккайдо до западной и восточной частей Берингова моря и Южной Калифорнии, глубина 887‒3640 м.
Neocrangon resima Rathbun 1902 (рис. 3a‒3n)
Рис. 3.
Neocrangon resima (Rathbun), самка 11.3 мм: a ‒ головогрудь со спины, сбоку и абдомен; b ‒ скафоцерит; c ‒ антенна I; d ‒ основание антенны II; e ‒ максилла I; f ‒ максилла II; g ‒ мандибула; h ‒ ногочелюсть II; i ‒ ногочелюсть III; j ‒ переопод I; k ‒ переопод II; l ‒ переопод III; m – переопод IV; n ‒ переопод V.
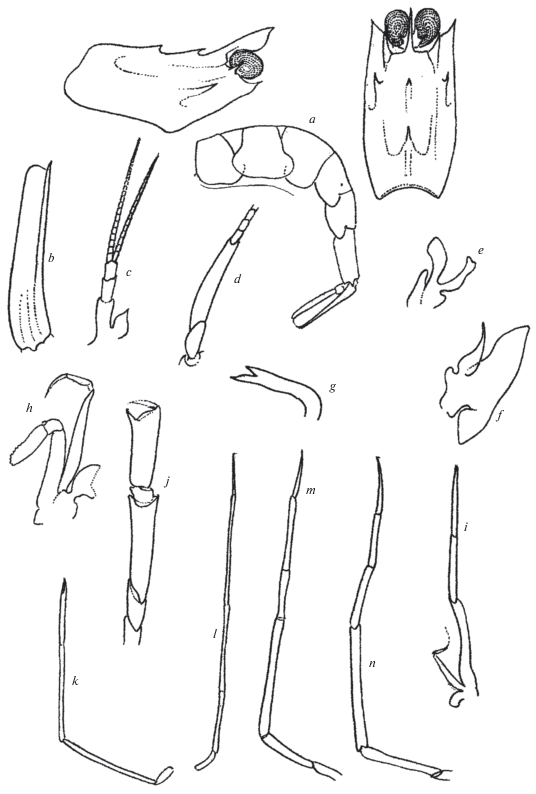
Материал. “Витязь”. Ст. 3540, 01.09.1955, 30°45′8 с.ш., 128°04′5 в.д., глубина 500 м, самка c яйцами, дефектная 11.3 мм.
Замечания. При общем соответствии краткому первоописанию (Rathbun, 1902) и последующему переописанию (Schmitt, 1921) наш экземпляр имеет ряд особенностей.
Первый медианный зубец на карапаксе лишь немного ниже заднего. На карапаксе имеются сильные печеночный и внешнеорбитальный зубцы. Рострум пластинчатый. вздернут вверх и заострен. Скафоцерит с вогнутым наружным краем и шипом, не выступающим за край пластинчатой части. Глаза темно коричневого цвета, крупные. Антенны I с заостренным стилоцеритом, наружный жгут немного длиннее внутреннего. Дистальный членик стебелька антенны II более чем вдвое длиннее базального. Мандибула крючковидная, двузубая. Ногочелюсти II и III с эпиподитом. Переопод II с клешней. Переопод III тонкий, слабее прочих. Наиболее сильным остается переопод V.
Поверхность сегментов абдомена гладкая, нижние края плевр ровные. Поверхность сегмента VI с парой низких продольных килей, не достигающих заднего края. Тельсон цилиндрической формы. Плеопод II самца имеет обычное для рода Neocrangon строение. Размеры яиц из яйцекладки самки 0.50‒0.70 мм.
Распространение. До настоящего времени был известен только в акватории от Сан-Франциско до мыса Сан Доминго, Калифорния, глубина 274‒487 м (Schmitt, 1921). Впервые найден в Западной Пацифике, в Восточно-Китайском море, глубина 500 м. Это второй после Neocrangon abyssorum пример широкого северо-пацифического распространения.
Metacrangon knoxi (Yaldwyn 1960) (рис. 4a)
Рис. 4.
Строение сегмента VI абдомена у видов рода Metacrangon (М-морфотип): a ‒ M. knoxi (Yaldwyn), b ‒ M. spinirostris (Rathbun), c ‒ M. variabilis (Rathbun), d ‒ M. robusta (Kobjakova).
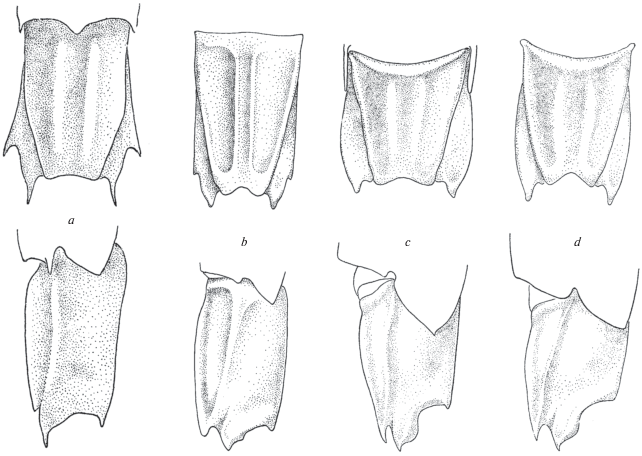
Материал. ”Дмитрий Менделеев”. Ст. 1277, 15.01.1976, 51°00′8 с.ш., 169°59′5 в.д., глубина 570 м, самка c яйцами, 11.5 мм.
Замечания. Удовлетворительно соответствует первоописанию голотипа, а также паратипа, яйценосной самки 11 мм, в яйцекладке которых насчитывается 45 яиц размером 1.5‒1.8 × × 1.2‒1.4 мм (Yaldwyn, 1960). У вновь обнаруженной яйценосной самки насчитывается 44 яйца, размером 1.5 × 1.8 мм.
Сегмент VI абдомена иного строения, нежели у N. abyssorum и N. resima. Он несет пару низких дорcальных килей и имеет неглубокую широкую выемку на заднем крае. Вентро-латеральный угол сегмента VI преобразован в широкую лопасть, которая заканчивается двумя заостренными шипами. Сходным образом устроен сегмент VI у некоторых других видов рода (рис. 4b–4d).
Среди прочих видов рода Metacrangon Zarenkov 1965 изучаемый вид выделяется массивным задним шипом на карапаксе, вершина которого изогнута и направлена вперед.
Распространение. Ранее был известен только на подводном поднятии Чатам, глубина 396‒638 м (Yaldwyn, 1960; Komai, 1997). Новая находка – в Большом Австралийском заливе, глубина 570 м.
Metacrangon spinirostris (Rathbun 1902) (рис. 4b)
Материал. “Витязь”. Ст. 2078, 05.1953, 44°29′0 с.ш., 148°38′0 в.д., глубина 1060 м, самец 9.8 мм.
Замечания. Так же как у M. knoxi, у данного вида имеется характерная особенность ‒ обособленная вентро-латеральная поверхность абдоминального сегмента VI. Однако она вооружена не двумя парами шипов, как у M. knoxi, а одной парой широких приостренных выступов. Плеопод II самца имеет обычное для видов этого рода строение.
Распространение. Берингово море, акватория Алеутских и Командорских островов, глубина 49‒1125 м (Макаров, 1941; Rathbun, 1902), а также воды Тихого океана у южных Курильских о-вов, глубина 780‒1540 м (Бирштейн, Заренков, 1970). Впервые найден в Охотском море у южных Курильских о-вов, глубина 1060 м.
Metacrangon variabilis (Rathbun 1902) (рис. 4c)
Материал. “Витязь”. Ст. 33, 18.08.1949, 59°50′0 с.ш., 155°22′0 в.д., глубина 84 м, 1 экз. Ст. 88, 08.1949, 59°11′6 с.ш., 148°57′8 в.д., глубина 52 м, 1 самка, 1 juv. Ст. 584, 11.09.1950, 61°58′2 с.ш., 179°44′2 в.д., глубина 136 м. Ст. 1316, 05.1952, 50°28′4 с.ш., 157°23′7 в.д., глубина 360‒380 м, 1 экз.
Замечания. Строение сегмента VI абдомена M. variabilis такое же, как у M. robusta (Kobjakova 1935) (рис. 4d).
Распространение. Воды Командорских и Алеутских островов, а также от Прибыловых о-вов до о-ва Св. Николая, Калифорния, глубина 90–1215 м (Макаров, 1941; Rathbun, 1904; Schmitt, 1921). Кроме того, воды Курильских о-вов (Шикотан, Итуруп) (Кобякова, 1958) и тихоокеанские воды у о-ва Парамушир, глубина 445‒780 м (Бирштейн, Заренков, 1970). Новые находки значительно расширяют ареал вида: Олюторский залив, северная часть Охотского моря (у полуостровов Пьягина и Хмитиевского), воды юго-восточной Камчатки, глубина 52‒380 м.
Rhynocrangon sharpi (Ortmann 1902) (рис. 5a)
Рис. 5.
Строение сегмента VI абдомена (Z-морфотип): a ‒ Rhynocrangon sharpi (Ortann), b ‒ Argis lar (Оwen), c ‒ Sclerocrangon alata Rathbun.
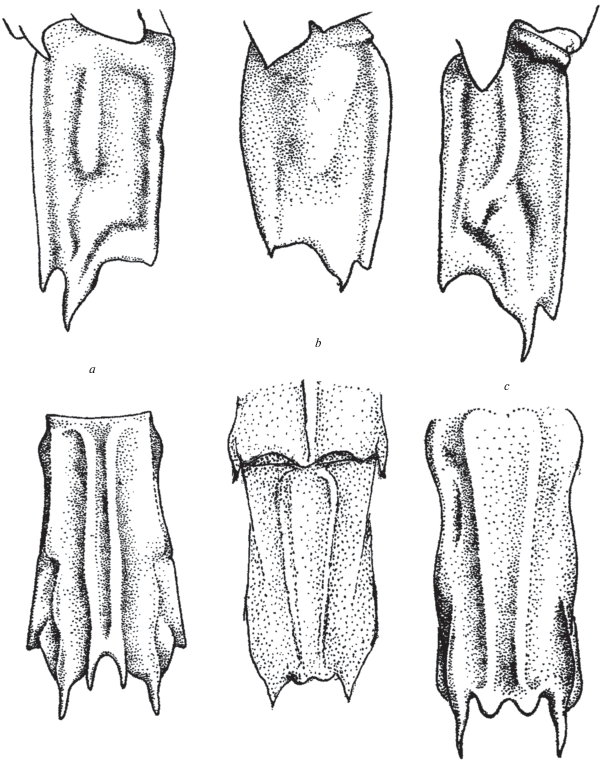
Материал. “Витязь”. Ст. 523, 16.08.1950, 56°10′8 с.ш., 176°32′1 в.д., глубина 106 м. Ст. 526, 18.08.1950, 55°32'7 с.ш., 165°08'5 в.д., глубина 125 м. Ст. 529, 11.09.1950, 61°58′2 с.ш., 179°44′2 в.д., глубина 110 м. Ст. 596, 15.09.1950, 59°50′9 с.ш., 170°30′5 в.д., глубина 75 м. Ст. 962, 30.09.1951, 54°28″3 с.ш., 168°18′6 в.д., глубина 123 м. Ст. 1331, 16.05.1952, 51°50′9 с.ш., 158°13′1 в.д., глубина 83 м. Ст. 1372, 22.05.1952, 54°59'4 с.ш., 162°09′0 в.д., глубина 122 м. Ст. 1385, 24.05.1952, 55°55′0 с.ш., 162°56′0 в.д., глубина 50 м. Ст. 1454, 05.05.1952, 60°07′8 с.ш., 171°48′0 в.д., глубина 141 м.
Замечания. Перечисленные находки цитируются по архивным материалам Я.А. Бирштейна и Л.Г. Виноградова. Количество экземпляров, найденных на станциях, и их размеры остались не известны.
Выделяясь среди видов семейства относительно крупными размерами, настоящий вид отличается и сильным развитием сложной скульптуры. Это распространяется на Argis lar (Owen 1839) (рис. 5b) и Sclerocrangon alata (Rathun 1902) (рис. 5c). Больше того, свойственный этим видам сложный скульптурированный сегмент VI абдомена наблюдается и у других видов Sclerocrangon (рис. 6). Разнообразие скульптуры этого сегмента могло бы служить предметом классификации.
Рис. 6.
Строение сегмента VI абдомена (Z-морфотип): a ‒ Sclerocrangon derjugini Kobjakova, b ‒ S. salebrosa Owen, c ‒ S. ferox G.O. Sars, d ‒ S. boreas Phipps.
Распространение. Берингово море: на востоке примерно от 58°05′ с.ш. до Аляски и Алеутских о-вов, на западе от 57°19′ с.ш. до юго-восточной Камчатки, глубина 32‒275 м (Макаров, 1941; Rathbun, 1904). Новые находки свидетельствуют о широком распространении этого вида в западной части Берингова моря – вплоть до Анадырского залива, глубина 50‒141 м.
Sclerocrangon zenkevitchi Birstein et Winogradow 1953 (рис. 7a–7b)
Рис. 7.
Sclerocrangon zenkevitchi Birstein et Winogradow: a ‒ сегмент VI абдомена со спины и сбоку (Z-морфотип). b ‒ двуветвистый плеопод II самца, ветви неравной длины. Sclerocrangon derjugini Кobjakova: c ‒ одноветвистый плеопод II самца, одна из ветвей почти полностью редуцирована.
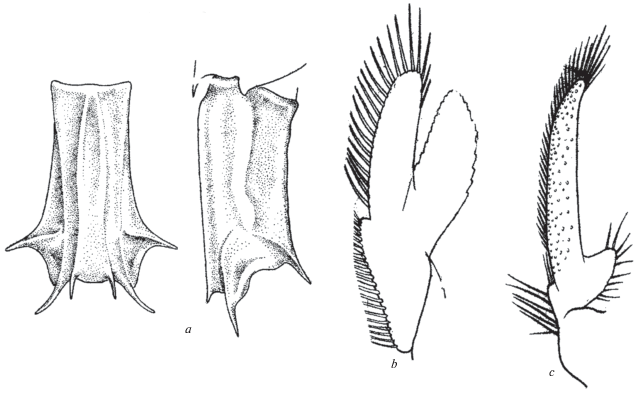
Материал. ”Витязь”. Ст. 1604, 28.06.1952, 57°43′ с.ш., 167°23′0 в.д., глубина 3661 м, 10 экз. Ст. 7502, 22.06.1976, 37°36′1 с.ш., 143°03′2 в.д., глубина 3620‒3640 м, 10 экз. самцы 16.0‒22.2 мм, 10 экз. пол не определен 6.5‒11.0 мм.
Замечания. Вместе с некоторыми другими видами рода, например S. boreas (Phipps 1774), S. salebrosa (Owen 1839), S. derjugini Kobjakova 1936, настоящий вид характеризуется своеобразной скульптурой сегмента VI абдомена: дорсальные и боковые гребни переходят в искривленные шипы. Плеопод II самца двуветвистый, с длинными щетинками, appendix masculinum короче эндоподита. Ветви плеопода мало отличаются по длине, однако у большинства изученных экземпляров эти различия довольно значительны.
Распространение. Обнаружен на склонах Командорской и Алеутской котловин Берингова моря, в северной части Курило-Камчатского желоба, глубина 2995‒4070 м, а также в Тихом океане у о-ва Хонсю, глубина 3620 м (Бирштейн, Виноградов, 1953; Бирштейн, Заренков, 1970). Упоминание о находке в Японском море (Заренков, 1993) является недоразумением: находка состоялась в Командорской котловине Берингова моря, глубина 3661 м.
ОБЩИЕ ВОПРОСЫ СИСТЕМЫ СЕМЕЙСТВА CRANGONIDAE НА ПРИМЕРЕ НЕКОТОРЫХ ВИДОВ СЕМЕЙСТВА
Изучаемые креветки различаются размерами. В имеющейся у автора коллекции длина карапакса креветок колеблется от 20 (Neocrangon abyssorum – пример мелких слабо скульптурированных креветок) до 170 мм (S. salebrosa – пример крупных сильно скульптурированных креветок). При этом диаметр яиц, которые вынашиваются самкой, составляет соответственно 0.45 и 4.5 мм. Следовательно, в некоторой мере размеры яиц скоррелированы с размерами организма.
Сходным образом с размерами креветок скоррелирована и их скульптура, в частности строение сегмента VI абдомена. У крупных особей она отличается сложностью, тогда как у более мелких остается относительно простой. Эти различия могли бы считаться результатом неравномерного аллометрического роста. Он мог осуществиться не только в эволюции, но и в индивидуальном развитии. На эту мысль наводит и сравнение просто устроенных ювенильных особей со зрелыми S. zenkevitchi (Бирштейн, Виноградов, 1953).
В уже упоминавшейся работе Кристоферсена (Christoffersen, 1988) фактически рассматривается комбинаторика 74 признаков (“characters”). Некоторыми из них служат структуры сегмента VI абдомена, частично использованные ранее с применением комбинаторного подхода (Заренков, 1965). В связи с этим у изученных креветок выделяются, по крайней мере, три морфотипа абдоминального сегмента VI.
Первый морфотип, условно называемый морфотипом Z, характерен для относительно крупных сильно скульптурированных креветок рода Sclerocrangon G.O. Sars 1883, например S. zenkevitchi, S. derjugini, S. igarashii Komai et Amaoka 1991, S. salebrosa, S. alata, а также Rhynocrangon sharpi, Argis lar. Морфотипу Z свойственны длинные шипы, которые служат продолжениями продольных килей абдоминального сегмента VI (рис. 5–6, 7a).
Второй морфотип обозначен буквой M. Он характеризуется несколько обособленными боковыми поверхностями абдоминального сегмента VI, которые несут одну или две пары краевых шипов (рис. 4). Морфотип M свойствен, например, видам рода Metacrangon: М. knoxi, M. robusta (Kobjakova 1935), M. spinirostris, M. variabilis и др.
Форма третьего морфотипа, С, приближается к цилиндру, который несет пару дорсальных слабо выраженных продольных килей и вооружен парой шипов на заднем крае сегмента (рис. 2). Примером служат N. abyssorum, N. communis, N. resima и Mesocrangon intermedia. Возможно, что к С-морфотипу относятся и другие виды этих родов (Заренков, 1965). Следует допускать и возможность существования иных морфотипов, отличных от морфотипов Z, M и С.
В определенной мере разнообразие морфотипов VI сегмента абдомена представляется комбинациями мелких структур – гребней и шипов. Полная комбинация структур осуществилась у морфотипа Z, отсутствие некоторых структур – у морфотипов М и С. Можно допускать, что комбинаторика структур будет разрастаться по мере изучения таксономического разнообразия.
Это замечание относится и к структурам плеопода II самцов. Едва ли не у всех видов изученной коллекции они состоят из двух придатков, покрытых длинными щетинками. У некоторых видов Sclerocrangon одна из ветвей плеопода редуцирована. Соответственно различаются два варианта строения плеопода: вариант 1 ‒ придатки сопоставимы по длине (рис. 7b), и реже встречающийся вариант 2 ‒ наружный придаток, возможно являющийся appendix masculinum, по длине значительно превосходит редуцированный эндоподит (рис. 7c). Вариант 1 свойствен многим видам родов Crangon Fabricius 1798 и Мetacrangon.
У Notocrangon antarcticus (Pfeffer 1887) придатки плеопода II самца редуцированы полностью за исключением удлиненного appendix masculinum, терминально вооруженного шипами (Заренков, 1968). В настоящее время известно, по крайней мере, два случая полной редукции внутреннего придатка плеопода ‒ Sclerocrangon derjugini и S. igarashii (Komai, Amaoка, 1991). Два альтернативных суждения об этом признаке ‒ ”внутренний придаток имеется или отсутствует” ‒ могли бы сочетаться с суждениями о структурах (characters) в общей комбинаторике, как у Кристоферсена (Christoffersen, 1988).
Честь открытия принципа комбинаторного разнообразия органов принадлежит Кювье (Cuvier, 1835, p. 64). В случае осуществления всех комбинаций структур разнообразие получает выразительное название “континуум” (Заренков, 1976; Silvestri, Hill, 1976) и представляется “пространством логических возможностей” (Заварзин, 1974).
О широком распространении комбинаторики в определенной мере свидетельствуют кладограммы и обычные дихотомические ключи для определения. Фактически они служат формой представления комбинаторной структуры разнообразия, наделенного статистической структурой (Заренков, 1984).
Список литературы
Бирштейн Я.А., Виноградов Л.Г., 1951. Новые и редкие десятиногие ракообразные Охотского моря и Курильских вод // Доклады АН СССР. Т. 79. № 2. С. 357‒360.
Бирштейн Я.А., Виноградов Л.Г., 1953. Новые данные по фауне десятиногих ракообразных (Decapoda) Берингова моря // Зоологический журнал. Т. 32. № 2. С. 215‒228.
Бирштейн Я.А., Заренков Н.А., 1970. О донных десятиногих ракообразных (Crustacea, Decapoda) района Курило-Камчатского желоба // Труды Института океанологии. Т. 86. С. 420‒426.
Заварзин Г.А., 1974. Фенотипическая систематика бактерий. М.: Наука. 143 с.
Заренков Н.А., 1965. Ревизия родов Crangon Fabricius и Sclerocrangon G.O. Sars (Decapoda, Crustacea) // Зоологический журнал. Т. 44. № 12. С. 1761‒1774.
Заренков Н.А., 1968. Десятиногие ракообразные (Crustacea, Decapoda), собранные Советскими антарктическими экспедициями в Антарктической и Антибореальной областях // Исследования фауны морей. Т. 6. № 14. С. 153‒195.
Заренков Н.А., 1976. Лекции по теории систематики. М.: 140 с.
Заренков Н.А., 1984. Общебиологические принципы зоологической систематики. Автореф. дис. … докт. биол. наук. М.: Московский государственный университет. 33 с.
Заренков Н.А., 1993. О новой находке глубоководной креветки Sclerocrangon zenkevitchi Birstein & Winogradow (Crustacea, Decapoda) в Курило-Камчатской впадине и Японском море // Arthropoda Selecta. Т. 4. № 2. С.17.
Кобякова З.И., 1958. Десятиногие раки (Decapoda) района южных Курильских островов // Исследования дальневосточных морей СССР. № 5. С. 220‒248.
Макаров В.В., 1941. Фауна Decapoda Берингова и Чукотского морей // Исследования дальневосточных морей CCCР. № 1. С. 111‒163.
Bruce A.J., 1988. A new crangonid shrimp, Pontocheras arafurae gen. et sp. n., from Arafura Sea // Zoologica Scripta. V. 17. № 2. P. 213‒221.
Chace F.A., Jr., 1984. The caridean shrimps (Crustacea: Decapoda) of the Albatross Philippine Expedition, 1907-1910. Part 2: Families Glyphocrangonidae and Crangonidae // Smithsonian Contributions to Zoology. № 397. 63 p.
Chan T.-Y., 1996. Crustacea Decapoda Crangonidae; Revision of the three related genera Aegeon Agassiz, 1846, Pontocaris Bate, 1888 and Parapontocaris Alcock, 1901 / Resultats des campagnes MUSORSTOM. V. 15. Memoirs de Museum National d’Histoire naturelle. № 168. P. 269‒336.
Christoffersen M.L., 1988. Genealogy and phylogenetic classification of the world Crangonidae (Crustacea Caridea), with a new species and new records for the South Western Atlantic // Revista Nordestina de Biologia. V. 6. № 1. P. 43‒59.
Cuvier G., 1835. [Lettre à J.G. Metrud] Leçons d’Anatomie comparée. Paris.P.XIII.‒P.XXXIIJ.
Duris Z., 1992. Revision of Vercoia Baker (Crustacea: Decapoda: Crangonida Invertebrate Taxonomy. № 6. P. 1437‒1457.
Kim J.N., Hayashi K.-I., 2003. Syncrangon, a new crangonid genus, with rediscriptions of S. angusticauda (de Haan) and S. dentata (Balss) (Crustacea, Decapoda, Caridea) from East Asian Waters // Zoological Science. V. 20. P. 669‒682.
Komai T., 1991. Deep-Sea Decapod Crustaceans from the Pacific Coast of Eastern Hokkaido, Northern Japan (Crustacea, Decapoda, Penaeidea and Caridea) // Report of North Japan Sub-Committee for Bottom Fish, Results of Committee for Fish. Resources. № 24. P. 55‒96.
Komai T., 1995. Vercoia japonica, a new species of crangonid shrimp (Crustacea: Decapoda: Caridea) from Japan // Natural History Research. V. 3. № 2. P. 123‒132.
Komai T., 1997. Revision of Argis dentata and related species (Crustacea, Decapoda, Crangonidae, with description) of a new species from the Okhotsk sea // Journal of Crustacean Biology. V. 17. № 1. P. 135‒161.
Komai T., 2001. New record of a crangonid shrimp, Aegeon rathbuni (Crustacea: Decapoda: Caridea) from Japon, with notes on its tegumental scales // Natural History Research. V. 6. № 2. C. 67‒75.
Komai T., 2008. A world-wide review of species of the deep-water crangonid genus Parapontophilus Christoffersen, 1988 (Crustacea, Decapoda, Caridea), with descriptions of ten new species // Zoosystema. V. 30. № 2. P. 261‒332.
Komai T., Amaoka, 1991. A new species of the genus Sclerocrangon from Urup Island, Kurile Islands and east of Hokkaido (Crustacea, Decapoda, Crangonidae) // Proceedings of the Japanese Society of Systematic Zoology. № 44. P. 26‒37.
Komai T., Chan Tin-Yam., 2009. New genus and species of Crangonidae (Decapoda Caridea) with a large plate eye from the abyssal zone of Taiwan, Northwestern Pacific // Journal of Crustacean Biology. T. 29. № 2. P. 254‒265.
Komai T., Takeda M., 1989. Sclerocrangon unidentata, a new crangonid shrimp from the Pacific Coast of Honshu (Crustacea: Decapoda) // Bulletin of the Biogeographical Society of Japan. V. 44. P. 77‒84.
Rathbun M. J., 1902. Description of new decapod crustaceans of the northwest coast of North America // Proceeding of U.S. National Museum. V. 24. P. 885‒905.
Rathbun M.J., 1904. Decapods crustaceans of the Northwest coast of North America // Harriman Alaska Expedition. Ser. 10. P. 1‒210.
Schmitt W.L., 1921. The marine Decapoda Crustacea of California // University of Californian Publication in Zoology. 3. P. 1‒470.
Silvestri L.G., Hill L.R., 1964. Some problems of the taxonomic approach // Phenetic and phylogenetic classification. Systematic association. Publ. № 6. P. 87‒103.
Yaldwyn J.C., 1960. Crustacea Decapoda Natantia from the Chatham Rise: A deep water bottom fauna from New Zealand // New Zeeland Department of Scientific and Industrial. Research. Bulletin. 139 (1). Biological Results of Chatham Islands 1954 Expedition. Pt. I. P. 14‒56.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал