

Зоологический журнал, 2020, T. 99, № 6, стр. 698-705
Хоминг лесной сони (Dryomys nitedula, Rodentia, Gliridae)
А. В. Андрейчев a, *, О. С. Кияйкина a
a Национальный исследовательский Мордовский государственный университет
430000 Саранск, Россия
* E-mail: andreychev1@rambler.ru
Поступила в редакцию 29.01.2019
После доработки 06.02.2019
Принята к публикации 16.01.2020
Аннотация
Изучен хоминг лесной сони (Dryomys nitedula). Дистанция опытного перемещения отловленных и меченых сонь изменялась от минимальной (50 м) до максимальной (10 км). Проведено 57 экспериментальных выпусков. На основании распределения одиннадцати возвратов зверьков выявлено, что низкая вероятность возврата сохраняется с дистанции до 5 км. Расчет ожидаемой вероятности возврата сделан по формуле Фюррера. Время отсутствия на месте изъятия составляло до 4 сут при переносе на дистанцию до 100 м, от 4 до 7 сут – при переносе на дистанцию до 1000 м и начинало экспоненциально возрастать после переноса на дистанцию свыше 1 км. Время возврата нелинейно увеличивается с увеличением дистанции переноса. Это позволяет предполагать, что перемещенные зверьки обнаруживают домашний участок в результате случайных блужданий.
Стремление и способность животных возвращаться на свой участок обитания принято называть хомингом (англ. homing, от home – дом). Ключевым моментом для проявления хоминга является значимость для животного определенного места в пространстве, куда оно стремиться вернуться после насильственного перемещения. В качестве возможных механизмов при ориентированном хоминге приводится непосредственное “чувство дома”, при котором используются визуальные ориентиры (Collett et al., 1986; Hodara, Busch, 2006) или геомагнитный физиологический компас (Kimchi et al., 2004). Ориентационными механизмами могут служить различные системы восприятия “родных мест” опосредованные разными органами чувств.
Хотя основное внимание исследователей было уделено видам, перемещающимся и мигрирующим на большие расстояния, изучение хоминга наземных мелких млекопитающих также вызывает значительный интерес (Bovet, 1995; Ostfeld, Manson, 1996; Hodara, Busch, 2006; Prevedello et al., 2009; Купцов, 2004, 2013; Купцов, Щипанов, 2004 и др.).
Существуют три основные гипотезы для объяснения возможностей успеха млекопитающих в поиске “дома”. Успех поиска может быть обусловлен: (1) случайным поиском “дома” при постепенном проникновении на “свой” участок обитания после выпуска, (2) ориентированием на знакомых местностях между местом выпуска и “домом” и (3) врожденной способностью животных находить свои “дома”, руководствуясь навигацией (Bovet, 1992; Rogers, 1992). Однако для обнаружения дома млекопитающие преимущественно используют одну из двух систем поиска: “навигацию” и “случайное блуждание”. Выявить систему поиска, которой воспользовалась та или иная особь, можно при сравнении результатов эксперимента с результатами модели, пересказывающей успех хоминга при одной из систем поиска. Если в ходе возврата можно ожидать линейного увеличения времени отсутствия при увеличении дальности переноса, то при случайном поиске время должно увеличиваться экспоненциально, так как зависит от прироста обследованной площади. Кроме того, пользуясь формулой Фюрера (Furrer, 1973), можно рассчитать ожидаемый успех хоминга в зависимости от дистанции переноса и радиуса известного пространства.
В системе “случайного блуждания” хоминг имеет смысл рассматривать, начиная с дистанций, выходящих за пределы регулярно посещаемой территории участка обитания животного (Щипанов, Купцов, 2004; Щипанов и др., 2008). Следует учитывать, что размеры знакомого пространства – важная характеристика для расчета ожидаемого успеха возвратов насильственно перемещенных мелких млекопитающих, в этой системе могут оказаться значительно больше размеров участка (Griffo, 1961; Andrzejewski et al., 2000; Щипанов и др., 2011).
Окрестности участка обитания становятся известны зверьками в результате их нерезидентной активности: собственно расселения с натального участка и в ходе “экскурсий”. При этом с одной стороны особи формируют, а с другой используют “сигнальное” поле, которое может способствовать их возвратам (Щипанов, Купцов, 2004). В частности, в экспериментах с обыкновенными бурозубками выявлены два разных типа активности. Активность, которая обеспечивает животное необходимой пищей и может быть рассмотрена как фуражировочная, и эпизодическая активность за пределами участка, которая дает информацию об окружающем пространстве. Активность за пределами участка почти в 10 раз увеличивает радиус знакомого особи пространства (Щипанов и др., 2011).
Целью нашей работы было изучение хоминга лесной сони (Dryomys nitedula Pallas 1778) на территории биологической станции Мордовского университета. На территории биостанции длительное время существует устойчивая популяция лесной сони (Андрейчев, Кузнецов, 2012).
Известно, что радиус индивидуального участка обитания для разных видов сонь составляет от 50 до 300 м (Россолимо и др., 2001). У сони-полчка отмечены наибольшие расстояния ухода зверьков от мест кольцевания в 2.8 км, у сони орешниковой – в 1.6 км (Schulze, 1970). Средний диаметр участка обитания сони-полчка оценивается в 200 м (Gaisler et al., 1977). Средний диаметр участка обитания орешниковой сони оценивается в 100 м. По результатам кольцевания 75% случаев повторных выловов ограничено 150 м от мест выпуска, остальные – в пределах 350 м (Schulze, 1970). У лесной сони по результатам кольцевания установлено, что их радиус участка обитания в Беловежской Пуще составляет 150–300 м. Лишь один зверек был отловлен с 900 м от места кольцевания (Голодушко, 1959). Таким образом, имеющиеся в литературе сведения по хомингу сонь ограничиваются главным образом информацией по орешниковой соне и соне-полчку.
Рассмотрен успех обнаружения домашнего участка лесными сонями, перенесенными на различные расстояния. Данные по этому виду полностью отсутствуют. Поэтому в нашу задачу входит представить первые полученные данные, без углубленного статистического анализа.
МАТЕРИАЛ И МЕТОДЫ
Отлов производили с использованием живоловок конструкции Щипанова (Щипанов, 1987) в июле–августе 2018 г. Ловушки настораживали в светлое время суток в известных нам ранее местах обитания сонь (Андрейчев, Кузнецов, 2012; Grigoryeva et al., 2015), а также в новых местах, выявленных опытным путем. Ловушки устанавливали в дубравах на наклоненных ветвях деревьев на высоте от 1.5 до 2.2 м. В ловушках использовали комбинированную приманку из кусочков яблок (груш), хлеба, смоченного нерафинированным подсолнечным маслом, и семечек.
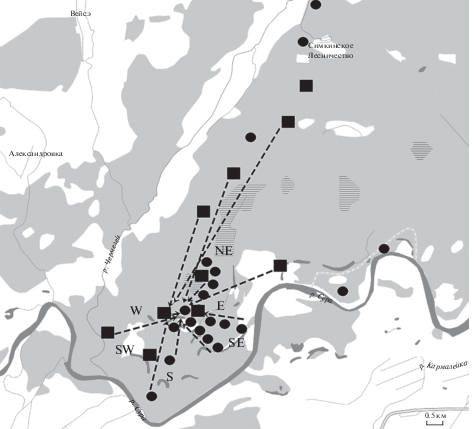
Как показали результаты наших предыдущих работ (Андрейчев и др., 2013) и рекогносцировочные отловы в данном исследовании на участке обитания сонь, меченные оседлые особи регистрируются в живоловках в среднем через сутки или даже каждые сутки. Т.е. наличие зверька на участке определяется фактически сразу. Таким образом, мы предполагали, что время возврата для лесной сони на первоначальный участок поимки после искусственного переселения на разные дистанции будет практически идентично времени отсутствия на месте изначального отлова с отклонением в одни сутки. Биотопы, в которые проводили выпуски сонь, представляют собой смешанные леса, сосняки-зеленомошники, сосняки-черничники, дубравы, березняки, осинники, ольшайники близ заливных пойменных лугов (табл. 1). Отловленных и помеченных зверьков перемещали к местам выпуска в непрозрачных контейнерах. Живоловки, в которые попадались изначально сони, после изъятия зверьков, оставались настороженными на этих же местах. На этих участках создавался избыток ловушек с интервалом между живоловками в среднем в 20 м, чтобы обеспечить быстрое обнаружение возврата перемещенных ранее сонь. Особей выпускали на разные расстояния от места отлова (“дома”). Дистанцию выпуска изменяли от минимальной (50 м) до максимальной (10 км). Интервал расстояния между отдельными выпусками изменяли через каждые 100 м до 1 км и через 1 км до 10 км. Таким образом, были выбраны следующие отметки: 50 м, 100 м, 200 м, 300 м, 500 м, 1 км, 2 км, 3 км, 4 км, 5 км, 6 км, 7 км, 8 км, 9 км, 10 км. Выпуски осуществляли в разных направлениях (рис. 1), зверьков не переносили за реку Сура со средней шириной русла 110 м в районе работ.
Таблица 1.
Распределение выпусков искусственно перемещенных особей лесной сони по биотопам, направлению и дистанции
| Биотоп | Выпуски (направление/дистанция перемещения, км | Всего | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Дубрава | W/0.05 | NE/4 | E/3 | E/3 | E/2 | E/2 | E/2 | E/0.4 | E/0.2 | S/0.2 | 10 | |
| Смешанный лес | NE/10 | NE/10 | NE/9 | NE/9 | NE/0.8 | NE/0.8 | NE/0.8 | NE/0.2 | NE/0.3 | NE/0.3 | 10 | |
| Сосняк- зеленомошник | NE/1 | NE/7 | NE/7 | NE/7 | NE/8 | NE/8 | 6 | |||||
| Сосняк-черничник | NE/4 | NE/4 | NE/2 | NE/2 | NE/2 | NE/2 | NE/0.1 | 7 | ||||
| Березняк | NE/5 | NE/5 | NE/5 | NE/5 | NE/5 | NE/3 | NE/3 | NE/0.9 | NE/0.9 | 9 | ||
| Осинник | SW/1 | S/1 | S/1 | NE/5 | SE/0.7 | SE/0.6 | S/0.5 | SE/0.5 | SE/0.5 | E/0.1 | E/0.1 | 11 |
| Ольшаник | NE/6 | NE/6 | NE/6 | E/0.3 | 4 | |||||||
| Всего | 37 выпусков NE; 1 выпуск W; 1 выпуск SW; 4 выпуска S; 4 выпуска SE; 10 выпусков E | |||||||||||
Рис. 1.
Карта-схема опытов по изучению хоминга лесной сони. Точки первичных (⚫) и повторных (◼) выпусков. Стрелками обозначены возвраты зверьков к “дому” после искусственных перемещений на разные дистанции, стрелками показаны возвраты зверьков.
На участках, где были отловлены зверьки, живоловки настораживали и проверяли каждые сутки, чтобы в последующем зарегистрировать возврат зверьков. Вернувшихся на свои участки зверьков повторно использовали в эксперименте. С каждым новым возвратом все больше увеличивалось расстояние между местом отлова и местом выпуска. Экспериментаторы перемещали зверьков на место выпуска, передвигаясь либо пешком, либо на автомобиле. Всего отловлено 47 особей: 23 самца (из них 6 неполовозрелых), 24 самки (из них 7 неполовозрелых). Проведено 57 экспериментальных выпусков.
Расчет ожидаемой вероятности возврата для случайного поиска выполнен по формуле Фюррера (Furrer, 1973): 2a sin(R/d)/π, где R – радиус знакомого пространства, d – дистанция перемещения. Для построения графиков на рис. 2 считали объединенные данные для дистанций до R = = 300 м, до R = 800 м по табл. 2.
Рис. 2.
Результативность хоминга (а) и среднее время отсутствия лесной сони на начальном пункте отлова после искусственных переселений на разные дистанции (б). Для б: кривая – экспоненциальный тренд для времени отсутствия на дистанциях свыше 0.8 км.
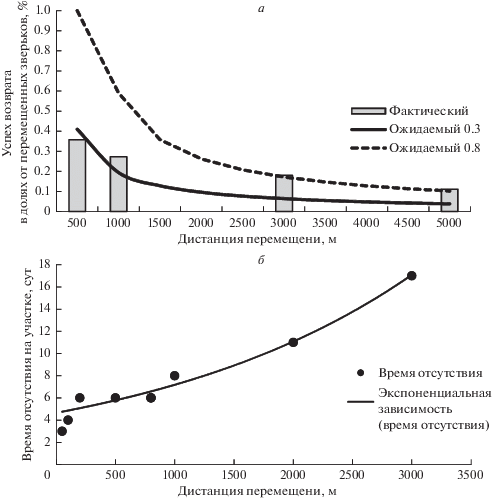
Таблица 2.
Объем и распределение данных по лесной соне в экспериментах в условиях биологической станции Мордовского университета
| Дистанция перемещения, м | Количество выпущенных особей | Количество вернувшихся особей | Среднее время отсутствия на участке, суток | ||
|---|---|---|---|---|---|
| первично | повторно | всего | |||
| 50 | 1 | – | 1 | 1 | 3 |
| 100 | 3 | – | 3 | 1 | 4 |
| 200 | 3 | – | 3 | 1 | 6 |
| 300 | 2 | 1 | 3 | – | – |
| 400 | – | 1 | 1 | 1 | 6 |
| 500 | 3 | – | 3 | 1 | 7 |
| 600 | 1 | – | 1 | – | – |
| 700 | – | 1 | 1 | – | – |
| 800 | 2 | 1 | 3 | 1 | 6 |
| 900 | 2 | – | 2 | – | – |
| 1000 | 2 | 2 | 4 | 2 | 8 |
| 2000 | 6 | 1 | 7 | 1 | 11 |
| 3000 | 3 | 1 | 4 | 1 | 17 |
| 4000 | 3 | – | 3 | – | – |
| 5000 | 5 | 1 | 6 | 1 | 16 |
| 6000 | 2 | 1 | 3 | – | – |
| 7000 | 3 | – | 3 | – | – |
| 8000 | 2 | – | 2 | – | – |
| 9000 | 2 | – | 2 | – | – |
| 10 000 | 2 | – | 2 | – | – |
РЕЗУЛЬТАТЫ
В результате работ получено 11 возвратов сонь, причем некоторые возвращались неоднократно после выпусков. Выявлено, что низкая вероятность возврата сохраняется с дистанции до 5 км (табл. 2). С дистанции свыше 5 км возвратов сонь не получено. Время отсутствия на месте изъятия составляло до 4 суток при переносе на дистанцию до 100 м, от 4 до 7 суток – на дистанцию до 1000 м, и начинало экспоненциально возрастать после переноса на дистанцию свыше 1 км. Снижение успеха хоминга с увеличением дистанции соответствовало ожидаемому для случайного перемещения. Однако если для коротких дистанций это соответствует ожиданию для R = 300 м, то для дальних переносов – для R = 800 м (рис. 2а).
Самки и самцы возвращались примерно с равной частотой. Из первично выпущенных особей (эксперимент 1) получено три возврата самок и три возврата самцов. Из повторных выпусков вернувшихся зверьков (эксперименты 2, 3, 4) получено 2 возврата от самки и три возврата от самца (табл. 3). В отношении неполовозрелых особей дистанция возврата очевидно существенно меньше, чем у взрослых особей, т.к. лишь одна особь вернулась “домой” с дистанции в 500 м. Повторный выпуск на дистанцию 700 м не дал возврата.
Таблица 3.
Характеристика и условия эксперимента по возвратившимся особям лесной сони, отловленных в дубраве
| № особи | Пол | Эксперимент 1 | Эксперимент 2 | Эксперимент 3 | Эксперимент 4 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Направление/ дистанция выпуска, км | Время возврата, суток | Направление/ дистанция выпуска, км | Время возврата, суток | Направление/ дистанция выпуска, км | Время возврата, суток | Направление/ дистанция выпуска, км | Время возврата, суток | ||
| 4 | Самец | W/0.05 | 3 | – | – | – | – | – | – |
| 8 | Самка | S/0.5 | 7 | – | – | – | – | – | – |
| 10 | Самка | SW/1 | 8 | – | – | – | – | – | – |
| 14 | Самец | S/0.2 | 6 | E/0.4 | 6 | NE/2 | 11 | NE/3 | 17 |
| 16 | Самец | NE/0.8 | 6 | – | – | – | – | – | – |
| 37 | Самка | NE/0.1 | 4 | NE/1 | 8 | NE/5 | 16 | – | – |
Взвешивания повторно отловленных меченых сонь не показали в 10 случаях из 11 изменения массы тела, даже после отсутствия (в результате странствий в поисках “дома”) в течение 3 недель. Единственным исключением является самец № 2, который вернулся с дистанции в 3 км за 17 суток и сбавил вес с 40 г (первоначальный отлов) до 37 г.
Большинство сонь возвращались “домой” в течение 0.5–2.5 недель (табл. 3). Рекордсменами по числу возвратов и пройденному расстоянию стали самец № 14 и самка № 37, которые возвращались 4 и 3 раза, соответственно. Примечательно, что эти особи возвращались “домой” и отлавливались повторно в те же ловушки, в которые были пойманы изначально. Самец в общей сложности преодолел более 5 км (0.2 км – первый возврат, 0.4 км – второй возврат, 2 км – третий возврат, 3 км – четвертый возврат). Самка же в общей сложности преодолела 6.1 км (0.1 км – первый возврат, 1 км – второй возврат, 5 км – третий возврат).
Мы наблюдали, как ведут себя зверьки после того, как мы их выпускали в биотопы, где сони не регистрировались нами. Соня изначально совершала рывок, с целью взобраться на ветку дерева, а затем затаивалась и осматривалась.
ОБСУЖДЕНИЕ
У лесной сони, среди известных для мелких млекопитающих отмечена наибольшая дистанция успешного возврата – 5 км. До сих пор самыми длительными зарегистрированными дистанциями по успешности возврата отмечены у флоридского кролика (Sylvilagus floridanus) – 4.8 км (Bowers, 1954; Rogers, 1992), серой белки (Sciurus carolinensis) – 4.5 км (Hungerford, Wilder, 1941), оленьего хомячка (Peromyscus maniculatus) – 3.2 км (Murie, Murie, 1931). Успех возврата лесной сони с дальней дистанции (5 км) низкий и составляет 0.16 (1 особь из 6 выпущенных). Что касается успеха возврата лесной сони на коротких дистанциях (до 300 м), то он составляет 0.3 (3 особи из 10 выпущенных). По успеху возврата с коротких и дальних дистанций лесная соня значительно уступает другим видам. По результатам работ Анджеевского с соавторами (Andrzejewski et al., 2000) по рыжим полевкам было выявлено, что зверьки имели высокий процент возвращений с дистанций более 600 м. Успешность хоминга составила 69.2%. Средний период времени между выпуском и повторным отловом был от 3.2 ± 1.6 дней до 5.2 ± 2.8 дней, в зависимости от применяемой приманки. Ими установлено, что знание пространства за пределами регулярно используемой территории является определяющим фактором возвращения мелких млекопитающих с дистанций, сравнимых с длиной обычных ориентировочных выходов с участка (Andrzejewski et al., 2000). Такие перемещения мелких млекопитающих за границы своих индивидуальных участков обитания, т.е. выходы за пределы “дома”, являются нормальным поведением (Griffo, 1961; Crawley, 1969; Lidicker, 1985; Andrzejewski, Babinska-Werka, 1986; Liro, Szacki, 1987; Clark et al., 1988; Liro, Szacki, 1994; Szacki, 1999; Lira et al., 2007; Щипанов, Купцов, 2004; Щипанов и др., 2011).
Распределение величин времени отсутствия лесных сонь в зависимости от дистанций перемещений выглядит на графике как экспонента и, скорее всего, свидетельствует о ненаправленном поиске, связанном с последовательным обследованием пространства (рис. 2б). Небольшое время отсутствия при переносах на дистанции 50–100 м вероятно связано с перемещением в пределах домашнего участка. При переносах на дистанции свыше 300 м время отсутствия сразу увеличивается на треть и до 800 м практически не меняется. Это, скорее всего, радиус известного пространства – R. После дистанции 0.8 км прирост времени при увеличении дистанции близок к ожидаемому для экспоненциального тренда.
Распределение успеха хоминга на коротких дистанциях визуально соответствует ожидаемому распределению для R = 0.3 км, а на дальних – для R = 0.8 км. Возвраты с дальних дистанции были отмечены у повторно перемещенных сонь. Заметим, что на большие дистанции переносили уже вернувшихся зверьков. Таким образом, увеличение радиуса знакомого пространства могло произойти в ходе предыдущих возвратов.
По-видимому, возвраты у лесных сонь, в основном, обусловлены случайными перемещениями. Доказано, что успех хоминга ряда других видов мелких млекопитающих является результатом случайного перемещения, а не определяется “чувством направления”. Так, например, наблюдения за 51 особью лесной мыши (Apodemus sylvaticus), помеченной хемолюминесцентными лампочками, показали, что зверьки не направились прямо к дому, но блуждали по зоне выпуска и преодолели большие дистанции (Jamon, Bovet, 1987). В ходе перемещений они ориентировались преимущественно на ландшафты, которые напоминали их обычную среду обитания, независимо от того, в каком направление находился их “дом”. Несмотря на хаотичность перемещений, успешность хоминга была хорошей, но время возврата зависело от первоначального направления, в котором они исчезали после выпуска. Радиус знакомого пространства может определяться как наличием эпизодических выходов, так и памятью о поисках участка при расселении от места рождения. Это один из предметов дальнейшего исследования.
Положение площадки, где проводились отловы сонь на биостанции относительно р. Сура, имеет большое значение, так как река может служить ориентиром, ограничивающим пространство “случайного” перемещения, т.е. в этом случае выступает обстоятельством, повышающим вероятность возврата.
Успешность хоминга зависит от расстояния перемещения, и дистанция возвращения мелких млекопитающих составляет обычно менее 2 км (Bovet, 1978). Виласенор (Villasenor et al., 2013) выделяет пороговые расстояния, после которых успешность хоминга сильно уменьшается. Этим автором на хомячках плоскогорий (Abrothrix longipilis) показано, что выпущенные недалеко от места отлова (≤100 м) особи остались живы, в то время как смертность достигла 22% на более длинных дистанциях перемещения, главным образом в результате борьбы между пришельцами и оседлыми грызунами того же вида. Таким образом, большие дистанции перемещения предотвращают быстрое возвращение (Villasenor et al., 2013). Мы склонны также считать, что уменьшение успеха хоминга у лесных сонь связано с гибелью нерезидентных зверьков. Взвешивания повторно отловленных сонь указывают на то, что особи не теряли вес в ходе своих блужданий в поисках “дома” и нормально питались. Предположительно факторами, которые могут увеличивать смертность сонь, выступают хищники из семейства куньих.
Лесные сони с каждым последующим выпуском приобретали опыт в ориентировании на незнакомой местности и использовали это для скорого возвращения “домой”. Об этой способности мелких млекопитающих известно из опытов Тефери и Милара (Teferi, Millar, 1993) на белоногом хомячке. Однако не следует исключать, что неопытные молодые зверьки при первых переносах могли погибнуть, а опытные взрослые выжить. Поэтому возможно здесь правомерно употребить формулировку “опыт выживания вне участка”.
Таким образом, можно сделать заключение, что лесная соня в значительной степени оседла на территории биологической станции, домашний участок в радиусе составляет около 50–100 м, а знакомое пространство особи не менее 300 м. По результатам работ можно предполагать, что успех хоминга у лесной сони является следствием случайных перемещений в поисках своего участка. Выяснение причины низкого успеха возвратов с коротких дистанций требует дальнейших специальных исследований.
Список литературы
Андрейчев А.В., Кузнецов В.А., 2012. Млекопитающие Мордовии: учеб. пособие. Саранск: Изд-во Мордовского университета. 100 с.
Андрейчев А.В., Орлов В.Н., Кривоногов Д.М., Григорьева О.О., Ютукова С.А., 2013. Соня лесная – редкий вид Мордовии // Редкие животные Республики Мордовия: материалы ведения Красной книги Республики Мордовия за 2013 год. Саранск: Изд-во Мордовского университета. С. 4–6.
Голодушко Б.З., 1959. Материалы по кольцеванию лесной сони в заповеднике “Беловежская Пуща” // Миграции животных. М. С. 186–189.
Купцов А.В., 2004. Нерезидентность у землероек-бурозубок 4 видов: Sorex araneus, S. caecutiens, S. minutus, S. isodon (Insectivora, Soricidae). Дис. … канд. биол. наук. М. 151 с.
Купцов А.В., 2013. Хоминг сеголеток обыкновенной (Sorex araneus) и средней (Sorex caecutiens) землероек-бурозубок (Insectivora, Soricidae) // Зоологический журнал. Т. 92. № 8. С. 941–954.
Купцов А.В., Щипанов Н.А., 2004. Хоминг у землероек-бурозубок Sorex araneus, S. caecutiens, S. minutus, S. isodon (Insectivora Soricidae) // Зоологический журнал. Т. 83. № 2. С. 213–219.
Россолимо О.Л., Потапова Е.Г., Павлинов И.Я., Крускоп С.В., Волцит О.В., 2001. Сони (Myoxidae) мировой фауны. М.: Изд-во Московского университета. 229 с.
Щипанов Н.А., 1987. Универсальная живоловка для мелких млекопитающих // Зоологический журнал. Т. 66. № 5. С. 759–761.
Щипанов Н.А., Купцов А.В., 2004. Нерезидентность у мелких млекопитающих и ее роль в функционировании популяции // Успехи современной биологии. Т. 124. № 1. С. 28–43.
Щипанов Н.А., Купцов А.В., Демидова Т.Б., Калинин А.А., Александров Д.Ю., Павлова С.В., 2008. Нерезидентность и расселение у обыкновенных бурозубок (Sorex araneus, Insectivora) // Зоологический журнал. Т. 87. № 3. С. 331–343.
Щипанов Н.А., Тумасьян Ф.А., Распопова А.А., Купцов А.В., 2011. Два типа использования пространства оседлыми обыкновенными бурозубками (Sorex araneus L.) // Известия РАН. Серия биологическая. № 1. С. 107–112.
Andrzejewski R., Babińska-Werka J., 1986. Bank vole population: are their densities really high and individual home range small? // Acta Theriologica. V. 31. P. 409–422.
Andrzejewski R., Babinska-Werka J., Liro A., Owadowska E., Szacki J., 2000. Homing and space activity in bank voles Clethrionomys glareolus // Acta Theriologica. V. 45. № 2. P. 155–165.
Bovet J., 1978. Homing in wild myomorph rodents: current problems, (in: Animal migration, navigation and homing. K. Schmidt-Koenig and W. T. Keeton, eds). Springer, Berlin. P. 405–412.
Bovet J., 1992. Mammals. (In: Animal homing. F. Papi, ed.). London: Chapman & Hall, P. 321–361.
Bovet J., 1995. Homing in red squirrels (Tamiasciurus hudsonicus): the importance of going straight // Ethology. V. 101. P. 1–9.
Bowers G.L., 1954. An evaluation of cottontail rabbit management in Pennsylvania. Trans. North Am. Wildl. Conf. V. 19. P. 358–367.
Clark B.K., Kaufman G.A., Fine E.J., Hand S.S., 1988. Long-distance movements by Reithrodontomys megalolis in tallgrass prairie // American Midland Naturalist. V. 120. P. 276–281.
Collett T.S., Cartwright B.A., Smith B.A., 1986. Landmark learning and visuo-spatial memories in gerbils. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology. V. 158. P. 835–851.
Crawley M.C., 1969. Movements and home ranges of Clethrionomys glareolus Schreber and Apodemus silvaticus L. in north-eastern England // Oikos. V. 20. P. 276–420.
Furrer R.K., 1973. Homing of Peromyscus maniculatus in the Channelled Scablands of East–Central Washington // Journal of Mammalogy. V. 54. № 2. P. 466–482.
Gaisler J., Holas V., Homolka M., 1977. Ecology and reproduction of Gliridae (Mammalia) in Northern Moravia // Folia Zoologica. V. 26. № 2. P. 213–228.
Griffo J.V., Jr., 1961. A study of homing in the cotton mouse, Peromyscus gossypinus // American Midland Naturalist. V. 65. P. 257–289.
Grigoryeva O., Balakirev A., Orlov V., Krivonogov D., Stakheev V., Andreychev A., 2015. Philogeography of the forest dormouse Dryomys nitedula (Gliridae, Rodentia) in Russian Plain and the Caucasus // Folia Zoologica. T. 64. № 4. C. 361–364.
Hungerford K.E., Wilder N.G., 1941. Observations on the homing behavior of the gray squirrel (Sciurus carolinensis) // Journal of Wildlife Management. V. 5. P. 458–460.
Hodara K., Busch M., 2006. Return to preferred habitats (edges) as a function of distance in Akodon azarae (Rodentia, Muridae) in cropfield-edge systems of central Argentina // Journal of Ethology. V. 24. P. 141–145.
Jamon M., Bovet P., 1987. Possible use of environmental gradients in orientation by homing wood mice, Apodemus sylvaticus // Behavioural Processes. V. 15. I. 1. P. 93–107.
Kahmann H., Ostermann K., 1951. Wahrnehmen und Hevorbringen honer Tome bei Kleinen Saugetioren // Experientia. V. 7. P. 268–269.
Kimchi T, Etienne A.S., Terkel J., 2004. A subterranean mammal uses the magnetic compass for path integration. Proceedings of the National Academy of Sciences. V. 101. P. 1105–1109.
Lidicker W.Z., Jr., 1985. Dispersal. The American Society of Mammalogists. Special publication. № 8. P. 420–454.
Lira P.K., Fernandez F.A.S., Carlos H.A.S., Curzio P.L., 2007. Use of a fragmented landscape by three species of opossum in southeastern Brazil // Journal of Tropical Ecology. V. 23. P. 427–435.
Liro A., Szacki J., 1987. Movements of field mice Apodemus agrarius (Pallas) in a suburban mosaic of habitats // Oecologia. V. 74. P. 438–440.
Liro A., Szacki J., 1994. Movements of small mammals along two ecological corridors in suburban Warsaw // Polish Ecological Studies. V. 20. P. 223–227.
Murie O.J., Murie A., 1931. Travels of Peromyscus // Journal of Mammalogy. № 12. P. 200–209.
Ostfeld R.S., Manson R.H., 1996. Long–Distance Homing in Meadow Voles, Microtus pennsylvanicus // Journal of Mammalogy. V. 77. № 3. P. 870–873.
Prevedello J.A., Delciellos A.C., Vieira M.V., 2009. Homing behavior of Philander frenatus (Didelphimorphia, Didelphidae) across a fragmented landscape in the atlantic forest of Brazil // Mastozoología neotropical. V. 16. № 2. P. 475–480.
Rogers L.L., 1992. Homing tendencies of large mammals: A review. St. Paul, Minnesota. P. 76–92.
Schulze W., 1970. Beitrage zum Vorkommen und Zur Biologie der Haselmaus (Muscardinus avellanarius L.) und des Siebenschlafers (Glis glis L.) im Sudharz // Hercynia. V. 7. №4. P. 355–371.
Szacki J., 1999. Spatially structured populations: how much do they match the classic metapopulation concept? // Landscape Ecology. V. 14. P. 369–379.
Teferi T., Millar J.S., 1993. Long distance homing by the Deer Mouse, Peromyscus maniculatus // Canadian Field Naturalist. V. 107. № l. P. 109–111.
Villasenor N.R., Escobar M.A.H., Estades C.F., 2013. There is no place like home: high homing rate and increased mortality after translocation of a small mammal // European Journal of Wildlife Research. V. 59. I. 5. P. 749–760.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал