

Зоологический журнал, 2020, T. 99, № 9, стр. 1014-1022
Холодостойкость и зимовка обыкновенной гадюки (Vipera berus, Reptilia, Viperidae) на острове Кижи, Карелия
Д. И. Берман a, Н. А. Булахова a, b, *, А. В. Коросов c, Н. Д. Ганюшина c
a Институт биологических проблем Севера ДВО РАН
685000 Магадан, Россия
b Научно-исследовательский институт биологии и биофизики
Томского государственного университета
634050 Томск, Россия
c Петрозаводский государственный университет
185035 Петрозаводск, Россия
* E-mail: sigma44@mail.ru
Поступила в редакцию 12.07.2019
После доработки 06.03.2020
Принята к публикации 03.04.2020
Аннотация
Ареал обыкновенной гадюки (Vipera berus (L. 1758)) ограничен на севере и северо-востоке, как считается, недостатком мест зимовки с температурами выше 0°C, поскольку она не переносит замораживания. Для подтверждения справедливости этого утверждения в лаборатории измерена холодостойкость гадюк, обитающих на острове Кижи (Карелия), и сопоставлена с температурами в зимовочных убежищах и грунтах названного региона. Замерзание взрослых змей происходило при средней температуре –3.0°C (диапазон –2…–3.9°C). Длительного замерзания V. berus, действительно, не выносит, но кратковременное (1–8 ч) – благополучно переживает. При малых отрицательных температурах (до –2…–2.2°C) змеи находились в переохлажденном состоянии, в котором пребывали от 23 до 121 сут. Вплоть до –1.5°C гадюки сохраняли подвижность – перемещались в экспериментальных контейнерах, что, вероятно, происходит и в зимовочных убежищах. Проведенная работа позволяет считать способность длительно находиться в состоянии переохлаждения (до 4 мес.) одной из важных составляющих адаптивной стратегии обыкновенной гадюки в холодных регионах. Критичной для вида температурой в горизонтах зимовки следует считать не 0°C, а –2°C. Названный порог может служить фактором, лимитирующим границу ареала гадюки. Температурные условия убежищ (каменных гряд) на острове Кижи и, вероятно, грунтов территории Карелии по этому критерию благоприятны для зимовки змей, что и отражается в высоком их обилии.
Обыкновенная гадюка (Vipera berus (L. 1758)) – одна из наиболее широко распространенных и массовых змей Северной Евразии; ее огромный ареал простирается от Атлантического океана до Якутии включительно. В большинстве работ, посвященных экологии вида, указывается, что гадюка зимует ниже уровня промерзания грунтов в полостях, расположенных в различных частях ареала от приповерхностных горизонтов до глубины 2 м. На севере и востоке распространение ограничено, вероятно, недостатком мест для зимовки. Их главная особенность – положительные температуры в течение всей зимы, поскольку эта змея, как считается, не переносит замораживания (Калецкая, 1956; Ларионов, 1961; Viitanen, 1967; Банников и др., 1977; Дробенков, 2001; Lenders, 2003).
Между тем, существуют лишь две публикации, в которых сделана попытка экспериментально оценить способность V. berus в Финляндии и Швеции выдерживать отрицательные температуры (Viitanen, 1967; Andersson, Johansson, 2001). Автор первой работы в кратких экспериментах показал, что минимальная температура воздуха, измеренная в террариуме и переносимая гадюками в течение нескольких часов, находится между –3.5 и –5°C и что криорезистентность молодых животных выше, чем взрослых. Andersson и Johansson (2001), напротив, считают, что новорожденные гадюки мало холодостойки, поскольку переносят лишь непродолжительное (2–3 ч) воздействие температур не ниже примерно –4°C. Кроме указанных работ, существует краткое замечание (без указания применявшихся методов), что взрослые гадюки могут благополучно переживать в течение 16 ч понижение температуры до –1.5°C, а при охлаждении до –5°C погибают, в первую очередь, мелкие особи (Павлов и др., 2004). Эти разночтения требуют проверки идентичности величины холодостойкости новорожденных гадючат и взрослых животных.
В Карелии, на Заонежском п-ове и прилегающих островах (около 62° с.ш.) гадюка обычна, а на о-ве Кижи особенно многочисленна. Здесь она успешно перезимовывает в сложенных на поверхности почвы грядах валунов со щебнем, собранных в прошлые века крестьянами при очистке полей (Коросов, 2010). Изобилие подходящих зимних убежищ – один из факторов поддержания высокой численности гадюк на острове. Однако температурные условия зимой в подобных убежищах не исследованы.
В настоящей работе решаются четыре сопряженные задачи:
1) оценка предельных величин отрицательных температур, переносимых взрослыми гадюками во время экспериментальной зимовки;
2) выяснение способности взрослых гадюк выживать после замораживания;
3) оценка возможной длительности пребывания взрослых гадюк при отрицательных температурах в состоянии переохлаждения;
4) сопоставление динамики температур в зимних убежищах гадюк на о-ве Кижи с холодостойкостью животных и температурами грунтов в регионе по данным метеостанций.
МАТЕРИАЛ И МЕТОДЫ
Животные для экспериментов собраны на о-ве Кижи, расположенном в северной части Онежского оз. (Республика Карелия, Российская Федерация, 62.08° с.ш., 35.21° в.д.) осенью 2016 (n = 14) и 2017 гг. (n = 21). Лабораторные исследования проведены в ИБПС ДВО РАН (г. Магадан).
Доставленных животных взвешивали и рассаживали поодиночке или попарно в пластмассовые прозрачные контейнеры (12 × 9 × 17 см) со слоем влажного мха на дне. В каждый контейнер помещали логгер DS1922L (iButton temperature loggers) для контроля температур. При проведении всех экспериментальных процедур влажный мох из контейнеров не удаляли.
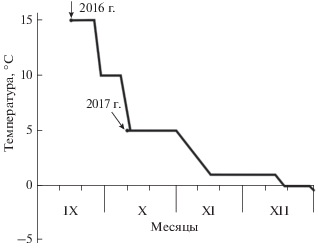
Акклимацию гадюк начинали в модифицированных охлаждающих термостатах ТСО–1/80 СПУ (от 15 до 3°С) и продолжали в морозильных шкафах Whirlpool AFB601AP (от 3 до 1°C). Последний этап акклимации (от 1 до 0°C) и дальнейшее охлаждение проводили в испытательной камере WTL 34/40. Акклимация состояла в постепенном понижении температур контейнеров с гадюками по схеме (рис. 1). Она представляла собой несколько сокращенный (в положительной области) аналог последовательности изменения температуры, наблюдаемой осенью при выхолаживании грунтов в природе в местах зимовки. Общая продолжительность акклимации (от 15 до 0°C) перед определением температуры максимального переохлаждения составила 94 суток (2016 г.), перед экспериментами по определению способности длительно находиться в переохлажденном состоянии (от 5 до 0°C) – 80 сут (2017 г.).
Рис. 1.
Режим акклимации гадюки в лаборатории перед экспериментами (стрелками показано начало акклимации в 2016 и 2017 гг.).
Холодостойкость оценивали по температуре максимального переохлаждения (Тп). Напомним, что Тп – самая низкая отрицательная температура, которую может выдержать не переносящее замерзания тканей животное. До достижения Тп холоднокровные животные в состоянии переохлаждения могут находиться разное время (от часов до месяцев), зависящее от множества факторов (Storey, Storey, 2012; Berman et al., 2016; Bulakhova et al., 2017 и др.).
По нашим предположениям, в отличие от мнения предыдущих исследователей (Viitanen, 1967; Andersson, Johansson, 2001; Павлов и др., 2004), обыкновенные гадюки способны на зимовке весьма значительное время пребывать в состоянии переохлаждения. Поэтому задача настоящего исследования – оценить его возможную продолжительность.
Определение Тп проведено стандартным методом с точностью ±0.1°C (Costanzo et al., 1995; Berman et al., 2016) на 8 гадюках массой 48–92 г. Термопары приклеивали с помощью медицинского пластыря к коже передней части тела змей, помещали животных в контейнерах в программируемую испытательную камеру WTL 34/40 и охлаждали со скоростью 0.02°C/ч (около 0.5°C/сут) до появления пика тепла кристаллизации на термограммах. Подобные скорости используются в исследованиях холодоустойчивости рептилий (см. ссылки выше). Через 1–8 ч после регистрации Тп часть животных (не вынимая из контейнеров) перемещали в холодильник с температурой 0.5–1°C, в котором они находились в течение нескольких суток (до появления признаков жизни или гибели), часть змей оставляли в камере с термопарами до замораживания. Весь процесс записывали с помощью компьютера.
У выживших при частичном замораживании животных после оттаивания и выяснения жизнеспособности через 6–10 сут на втором этапе эксперимента повторно определяли Тп для оценки динамики ее изменения. Процедура была полностью аналогична первичному измерению Тп.
Способность длительно пребывать при отрицательных температурах в состоянии переохлаждения оценивали для 17 взрослых гадюк массой 40–108 г, помещенных по 1–2 особи в описанных выше пластиковых контейнерах в модифицированные морозильные камеры Whirlpool AFB601AP или в испытательную камеру WTL 34/40.
Во всех использованных термостатах и камерах в разной степени выражены температурные градиенты, поэтому температуру змей фиксировали 24 раза в сутки поверенными логгерами DS1922L, прикрепленными медицинским пластырем в передней трети тела. Поверка логгеров осуществлялась перед началом и после завершения каждого эксперимента. Индивидуальная погрешность логгеров относительно декларированной производителем для всей серии приборов этой марки (±0.5°C) составила при 0°C для 10 логгеров +0.3°C, для 6 логгеров +0.2°C и для одного логгера +0.1°C. Данные о температурах в разделе “Холодостойкость” приведены с учетом этих погрешностей.
Полученные с помощью логгеров записи температуры тела змей в течение эксперимента индивидуальны. Максимальные их значения не превышали 1°C, а минимальные варьировали от –0.9 до –2.2°C. Время начала эксперимента для разных животных было разным, и продолжительность выживания рассчитывали с момента перехода температур в отрицательную область на период не менее 8–10 ч. Состояние змей (по подвижности) проверяли 1 раз в 2 недели, не вынимая контейнеры из холодильников. Один раз в месяц (7 февраля, 7 марта, 6 апреля и 7 мая) осторожно, чтобы не спровоцировать сотрясением кристаллизацию жидкостей тела переохлажденных животных, контейнеры извлекали из холодильников и быстро (чтобы не нагреть) осматривали через прозрачные стенки, отмечая неподвижных или замерзших гадюк (т.е. погибших). Учитывая вынужденную редкость детальных осмотров, за дату гибели принимали дату последней проверки, при которой животное было живо. Например, если при осмотре 7 февраля животное в контейнере двигалось, а при проверке через месяц (7 марта) – мертво, датой смерти считали 8 февраля. Такой подход, с одной стороны, значительно занижает оценку продолжительности выживания животных в эксперименте, но с другой, позволяет избежать ошибок в случаях, когда мы не могли гарантированно судить о состоянии животных на промежуточных осмотрах в термостатах.
Температурные условия зимовки гадюк в Карелии измерены в одной из каменных “гряд” (№ 980, см. ниже) в течение пяти зим: 2013/2014–2017/2018 гг. В качестве термометров использовали логгеры DS1921 (http://alfa-prom.ru/e_besprovodniye_registratory.html), которые устанавливали парами на грунте в центре гряды (глубина 1.3 м). Логгеры работали с октября по май с периодичностью отсчетов 3 и 4 ч; предварительно их не калибровали, заводская точность ±0.5°C, что позволяет получить оценку температуры в зимних убежищах гадюк в пределах ±1°C.
РЕЗУЛЬТАТЫ
Холодостойкость
Смертность до начала экспериментов. В 2016 г. во время транспортировки погибла одна гадюка, за 15 сут акклимации (при 10°C) – еще три. В 2017 г. за 25 сут акклимации (при 5°C) отход составил 4 гадюки. Эта смертность, очевидно, не связана с содержанием в лаборатории, в частности с температурой или режимом влажности, а, вероятно, с состоянием животных на момент отлова.
Температуры максимально переохлаждения. Средняя Тп при первичном тестировании составила –3.0 ± 0.3°C (табл. 1). Продолжительность пребывания гадюк при отрицательных температурах до достижения температуры максимального переохлаждения коррелировала с их массой (rs = 0.8, p ≤ 0.05).
Таблица 1.
Температура максимального переохлаждения (Тп) и выживание гадюк после первого и повторного (для выживших) этапов охлаждения
| Первый этап | Второй этап | Масса гадюки, г | ||||
|---|---|---|---|---|---|---|
| Тп, °C | Время после Тп, ч | Результат | Тп, °C | Время после Тп, ч | Результат | |
| –2.0 | 8.0 | ✓ | –5.0 | 20.3 | † | 48 |
| –2.5 | 14.0 | † | – | – | – | 55 |
| –2.0 | 72.0 | † | – | – | – | 70 |
| –3.0 | 1.5 | ✓ | –2.5 | 42.0 | † | 76 |
| –3.0 | 42.0 | † | – | – | – | 77 |
| –3.3 | 72.0 | † | – | – | – | 78 |
| –3.9 | 1.0 | ✓ | –4.2 | 0.4 | † | 88 |
| –3.9 | 2.5 | ✓ | –3.0 | 15.0 | † | 92 |
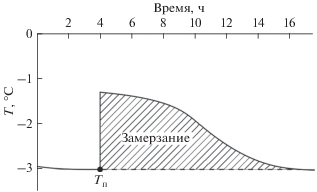
Замораживание особей массой 55–78 г отмечено через 12–15 ч после регистрации выброса тепла кристаллизации (рис. 2). Ни одна из исследованных гадюк не перенесла длительного замораживания, но несколько особей выжили после кратковременного замораживания (табл. 1). Выживание после начала кристаллизации жидкостей тела, вероятно, ограничено временем, проведенным при отрицательных температурах после прохождения Тп (rs = 0.88, p ≤ 0.05): выжили и самые мелкие, и наиболее крупные гадюки, с начала замерзания которых прошло не более 8 ч.
Рис. 2.
График охлаждения при определении температуры максимального переохлаждения (Тп) и последующего замерзания гадюки массой 77 г. Время замораживания животного составило около 13 ч.
При повторном охлаждении Тп выживших гадюк (n = 4) незначительно уменьшилась – до –3.7 ± ± 0.6°C (U-test 9.5, p = 0.3). У двух особей она оказалась ниже, чем у каждой из особей, исследованных при первичном определении Тп: –4.2 и –5°C. Из-за малой величины выборки установить зависимости величины повторной Тп от режима эксперимента и характеристик животных не представляется возможным.
Важной особенностью гадюк в состоянии переохлаждения следует считать сохранение подвижности при малых отрицательных температурах: вплоть до –1.5°C они не только медленно двигались в контейнерах, но и, реагируя на освещение, поднимали головы и шипели. Их способность к перемещению в более теплые горизонты зимовочных убежищ ранее наблюдалась при малых положительных температурах Ларионовым (1961) и Viitanen (1967).
Выживание при длительном пребывании в малых отрицательных температурах. Длительность нахождения гадюк в переменных режимах (возникающих за счет “плавания” температуры в камерах от –2.2 до 0.9°C) показана в табл. 2. В течение первого месяца эксперимента погибла одна особь, которую тестировали в щадящем режиме – от 0 до –0.9°C. Для еще одной особи известна лишь продолжительность выживания в эксперименте (912 ч), но нет данных о длительности воздействия и величине отрицательных температур (логгер перестал работать на вторые сутки). Эти данные не включены в табл. 2. Четырнадцать гадюк перенесли пребывание в термостатах с различными температурными режимами от 38 до 66 сут, в т.ч. они находились непрерывно в отрицательной области от 22 до 66 сут. Одна из гадюк выжила в эксперименте продолжительностью 128 сут (табл. 2), из них в течение 121 сут – при отрицательных температурах в интервале от 0 до ‒1.9°C.
Таблица 2.
Характеристики экспериментов по выяснению способности гадюк длительно существовать в состоянии переохлаждения
| № | Температура режима, °C | Продолжительность эксперимента, сут | Масса гадюки, г | |||
|---|---|---|---|---|---|---|
| min | max | общая | суммарная при Т < 0°C/ (из них при Т < –1°C) | непрерывная при Т < 0°C | ||
| 1 | –0.9 | 0.9 | 38.4 | 22.8/(0) | 22.0 | 72 |
| 2 | –1.0 | 0.9 | 37.8 | 29.2/(0) | 22.3 | 40 |
| 3 | –1.1 | 0.5 | 66.3 | 39.0/(12.3) | 24.6 | 102 |
| 4 | –1.2 | 0.1 | 61.5 | 58.8/(9.7) | 58.3 | 80 |
| 5 | –1.3 | 0 | 61.6 | 61.6/(21.6) | 61.6 | 72 |
| 6 | –1.3 | 0.8 | 66.5 | 64.5/(47.3) | 38.8 | 76 |
| 7 | –1.4 | 0.6 | 65.0 | 61.2/(5.3) | 39.7 | 52 |
| 8 | –1.4 | 0.4 | 66.4 | 64.8/(14.9) | 64.5 | 60 |
| 9 | –1.4 | 0.7 | 66.5 | 63.9/(47.2) | 38.7 | 78 |
| 10 | –1.4 | 0.3 | 64.8 | 64.5/(1.9) | 64.3 | 70 |
| 11 | –1.5 | 0.3 | 47.0 | 44.7/(33.9) | 40.9 | 60 |
| 12 | –1.8 | 0.7 | 66.4 | 52.4/(37.5) | 27.8 | 108 |
| 13 | –1.8 | 0 | 66.3 | 66.3/(6.3) | 66.3 | 76 |
| 14 | –1.9 | 0.9 | 128.0 | 121.0/(54.6) | 47.3 | 90 |
| 15 | –2.2 | 0 | 65.5 | 65.5/(28.4) | 65.5 | 52 |
Примечания. Общая продолжительность эксперимента – время, в течение которого гадюки были живы. Суммарная при Т < < 0°C и Т < –1°C – совокупная продолжительность периодов с температурой ниже 0 и с температурой ниже –1°C соответственно. Непрерывная при Т < 0°C – максимальная продолжительность пребывания гадюк при отрицательных температурах без возврата этих особей в положительную область.
Животные в эксперименте выдержали переменные режимы – с многократным возвратом из отрицательной области в положительную и обратно (что можно рассматривать в качестве модели приповерхностного расположения зимовочных убежищ осенью, весной и во время оттепелей). Они также успешно пережили постоянное и длительное пребывание в отрицательных температурах (модель более глубокого расположения места зимовки), находясь в состоянии переохлаждения, что свидетельствует об его устойчивости (табл. 2).
Возможная продолжительность благополучного выживания при температуре –1.5°C оказалась существенно больше, чем ранее наблюдали Павлов с соавторами (2004). Так, при определении Тп гадюки находились в условиях постоянного понижения температур после прохождения ‒1.5°C в течение по меньшей мере 25 ч, максимально – 160 ч. В экспериментах 2017 г. четырех гадюк содержали при температуре ниже –1.5°C от 21 до 1208 ч (суммарно) или 15 ч, 87 ч, 120 ч и 632 ч – непрерывно.
Температурные условия зимовки
На крупных островах Кижского архипелага (в т.ч. и на о-ве Кижи) гадюка многочисленна из-за благоприятных условий обитания: высокого обилия в болотах лягушек, служащих кормом, множества летних и зимних убежищ. Плотность животных здесь примерно на два порядка выше, чем в среднем по региону (Коросов, 2009). Основными убежищами для гадюк служат каменные “ровницы”, или “заборья” – сложенные по окраинам полей на суходольных лугах аккуратные гряды валунов со щебнем, собранных при очистке земли, начиная с XV в (Коросов, 2008). Гадюки успешно перезимовывают в них. На о-ве Кижи около 1000 таких гряд; на нескольких десятках из них гадюк встречали ранней весной на разогретых камнях склонов, где только и сходит к этому времени снежный покров.
Гряда № 980, в глубине которой изучали температурные условия зимовки, сложена из валунов разных размеров (3–40 см в поперечнике, в основном 10–15 см). Она имеет неправильную овальную форму (5 × 6 м при наибольшей высоте 1.3 м); южный склон около 60°. Пустоты между камнями снаружи заполнены лиственным опадом, не мешающим гадюкам уходить в глубину на зимовку. На гряде разреженный травяной покров, на поверхности камней – накипные лишайники, подножье со мхами и травой. Зимой она целиком покрыта снегом: по краям мощность снежного покрова около 1 м, по верху – 10–30 см. Гряда удалена от других подобных сооружений на 300 м, что дополнительно гарантирует отсутствие весенней иммиграции змей.
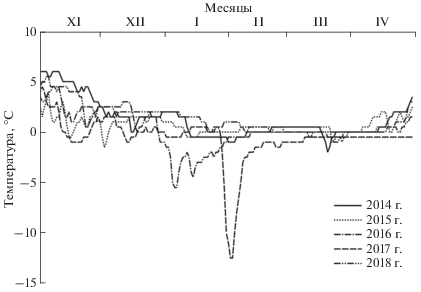
В центре гряды на уровне земли падение температур с летних значений (около 15°С) до близких к нулю отметок происходит в течение октября–декабря. Скорость охлаждения составляет от 0.1 до 0.2°C/сут в разные годы. Зимой температуры большую часть времени держались на названном уровне и лишь кратковременно (обычно в январе) опускались в отрицательную область (но не ниже –2°C) (рис. 3). В зимние сезоны двух лет из пяти (в 2016 и 2017 гг.) температура падала существенно ниже обычной: до –5.5 и до –12.5°C, соответственно (рис. 3). Последний случай связан с оттепелями, согнавшими снег с верхней части гряды, и с наступившими затем морозами, выстудившими ее. Это единственный пример значительного остывания центральной части зимовальной гряды. Несмотря на столь существенное падение температуры, змеи, тем не менее, выжили: в начале мая 2017 г. здесь была обнаружена одна самка (обычно после зимовки отмечались 3–6 гадюк). Таким образом, есть основания полагать, что по периферии гряды под неповрежденным снежным покровом температура была не столь низка, и время ее воздействия было не столь продолжительным, как в центре.
ОБСУЖДЕНИЕ
Как показано выше, взрослые гадюки выдерживают отрицательные температуры в состоянии переохлаждения вплоть до –5°C, что несущественно ниже минимальной Тп (–4.6°C), зарегистрированной Andersson и Johansson (2001) у новорожденных животных. Выявленные диапазоны Тп также близки: у змей из Карелии –2.0…–5.0°C, у змей из Швеции –2.8…–4.6°C. Важно, что эти данные получены на разительно различающихся размерами животных и скоростях их охлаждения. Andersson и Johansson (2001) использовали гадюк средней массой 4.1 г и со средней длиной тела 18.1 см. Скорости охлаждения в двух из трех серий их экспериментов достигали 5°C/ч, что весьма далеко и от наблюдающихся в природе, и от принятых в практике исследования холодоустойчивости пойкилотермных позвоночных (Costanzo et al., 1995; Berman et al., 2016). Впрочем, третий эксперимент со значительно меньшими скоростями охлаждения (0.2°C/ч) не изменил характера результатов Andersson и Johansson (2001).
Выделим два важных обстоятельства. Во-первых, температуры максимального переохлаждения шведских и карельских гадюк очень близки; во-вторых, они не зависят от возраста, размеров животных и скорости охлаждения (U-test, p ≥ 0.05). Все это позволяет объединить значения из работы Andersson и Johansson (2001) и полученные нами в единую выборку (рис. 4), что увеличивает надежность средней Тп= –3.1 ± 0.1 (–2… –4.6)°C обыкновенной гадюки.
Рис. 4.
Распределение Тп гадюк по классам температур в объединенной выборке (наши данные и данные Andersson, Johansson, 2001).
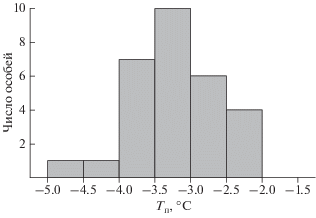
Как следует из анализа объединенной выборки Тп (рис. 4, табл. 1), и ювенильные, и взрослые гадюки способны без ущерба кратковременно (в течение нескольких часов или суток) переносить постепенное понижение температур в интервале 0…–1.9°C; замерзания животных при этих температурах не отмечено ни Andersson и Johansson (2001), ни нами. У значительной части особей в объединенной выборке (почти у 2/3) оно начиналось лишь при температурах, которые были ниже или равны –3°C (рис. 4).
Результаты экспериментов 2017 г. позволяют заключить, что благополучное пребывание гадюк как при малых отрицательных температурах, так и в условиях переменного режима (при возврате в положительную область и обратно), может измеряться не часами и сутками, а неделями и месяцами (табл. 2).
Хотя оценка продолжительности выживания из-за редкости проверок весьма приблизительна, между этим параметром и массой тела животных прослеживается связь (rs = 0.53, p ≤ 0.05), т.е. чем меньше масса, тем быстрее змея погибала в эксперименте. Эти результаты требуют специальных исследований, поскольку причина смерти гадюк остается не ясной.
Полученные данные позволяют заключить, что температурным критерием благополучной зимовки обыкновенной гадюки следует считать не 0°C, а –1.9…–2.0°C.
Длительное замораживание, как и в экспериментах Viitanen (1967), а также Andersson и Johansson (2001), в наших опытах гадюки не выдерживали. Тем не менее, они без ущерба переносили частичное замораживание, происходившее в течение 1–8 ч с момента начала кристаллизации жидкостей тела.
На о-ве Кижи, как и на других островах архипелага, представляющих собой озы (особый вид песчано-гравийно-галечных с валунами ледниковых отложений), нет скальных выходов, по расщелинам которых гадюки могли бы проникать в глубокие и потому непромерзающие горизонты грунтов. Поэтому природа очага высокой численности гадюк на Кижах, несомненно, антропогенная. На некоторых других островах (например, Северный Олений) скалы есть, и здесь формируются природные очаги.
Грунты в Карелии при ненарушенном снежном покрове промерзают до 40–80 см на разных почвах (см. табл. 3). Анализ данных метеостанций свидетельствует о том, что район Петрозаводска не уникален по зимним температурным условиям в почве, а грунты островов-озов, вероятно, еще теплее. На показанном температурном фоне в Карелии подходящие для зимовки гадюк места, по-видимому, не являются редкостью. Однако зафиксированные под ровницами на Кижах минимумы на поверхности почвы в холодные и малоснежные зимы могут быть существенно ниже переносимых гадюками температур (–2°C) – до ‒12°C, что видимо, влечет за собой гибель части популяции.
Таблица 3.
Минимальные из среднемесячных температур почв в Карелии на некоторых метеостанциях (Справочник…, 1965)
| Метеостанция | Почва | Глубина, см | Мин. из средн. мес. Т°С |
|---|---|---|---|
| Лоухи | Торфяная | 20 | –0.7 |
| 40 | 0.2 | ||
| 80 | 1.1 | ||
| Песчаная | 20 | –0.9 | |
| 40 | –0.5 | ||
| 80 | 0.0 | ||
| Реболы | Супесчаная | 20 | – |
| 40 | 0.0 | ||
| 80 | 0.7 | ||
| Петрозаводск | Супесчаная | 20 | –1.8 |
| 40 | –1.1 | ||
| 80 | –0.4 | ||
| Медвежьегорск | Песчаная | 20 | –3.4 |
| 40 | –2.2 | ||
| 80 | –0.6 | ||
| Сортавала | Песчаная | 20 | –1.2 |
| 40 | 0.0 | ||
| 80 | 0.6 |
ЗАКЛЮЧЕНИЕ
Полученные данные по холодостойкости обыкновенной гадюки позволяют заключить, что на зимовке эти змеи, будучи переохлажденными, могут в течение длительного времени переносить первые градусы ниже нуля, многие особи – в течение нескольких месяцев.
Важной выявленной в эксперименте особенностью V. berus следует считать сохранение подвижности в состоянии переохлаждения до ‒1.5°C. Весьма вероятно, что при малых отрицательных температурах гадюки перемещаются и в зимних укрытиях. При таких температурах они проявляют оборонительные реакции – отчетливо шипят.
Длительного замораживания змеи не переносят (после частичного – восстанавливаются). Поэтому температуры ниже –2°C (вплоть до –5°C кратковременно) представляют собой область риска. Проведенная работа позволяет считать не 0°C, а –1.9…–2°C температурной областью, критичной для зимовки этого вида. Названный порог может служить фактором, лимитирующим границу ареала гадюки.
Температурные условия грунтов Карелии по этому критерию весьма благоприятны для зимовки змей, что и отражается в высоком их обилии. Роль гряд из валунов в поддержании высокой численности обыкновенной гадюки, вероятно, состоит в создании многочисленных убежищ, расположенных рядом с летними ее биотопами. Животные свободно перемещаются между грядами, не образуя массовых зимовок (Коросов, 2008). Последнее обстоятельство подтверждает мнение Калецкой (1956) о том, что “…крупные скопления гадюк наблюдаются там, где имеется мало пригодных для зимовки мест…” (стр. 102).
В связи с обсуждением холодостойкости нельзя не упомянуть возможное расширение области распространения и увеличение численности гадюки в холодных регионах за счет потепления и антропогенных преобразований ландшафтов, наблюдающееся, по мнению Емцева с коллегами (2012), на севере Западной Сибири. Важнейшим критерием оценки пригодности новых мест для зимовки этих змей может служить выявленная нами критическая для гадюк во время зимовки температурная область, которую можно кратко назвать “не ниже –2°C”.
Список литературы
Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н., 1977. Определитель земноводных и пресмыкающихся фауны СССР. М.: Просвещение. 414 с.
Дробенков С.М., 2001. Экология и социальная организация популяции гадюки обыкновенной (Vipera berus) в зимний период // Актуальные проблемы герпетологии и токсинологии. Вып. 5. С. 45–50.
Емцев А.А., Берников К.А., Акопян Э.К., 2012. О расширении границ ареалов некоторых видов животных в северной части Западной Сибири // Мир науки, культуры, образования. № 6 (37). С. 471–477.
Калецкая М.Л., 1956. К биологии гадюки // Природа. № 5. С. 101–102.
Коросов А.В., 2008. Динамика численности островной популяции обыкновенной гадюки (Vipera berus) // Зоологический журнал. Т. 87. № 10. С. 1235–1249.
Коросов А.В., 2009. Распространение обыкновенной гадюки на островах Кижского архипелага // Труды Карельского НЦ РАН. № 4. С. 102–108.
Коросов А.В., 2010. Экология обыкновенной гадюки (Vipera berus L.) на Севере (факты и модели). Петрозаводск: Издательство Петрозаводского госуниверситета. 264 с.
Ларионов П.Д., 1961. Об условиях зимовки гадюк на северной границе их распространения в долине Лены // Зоологический журнал. Т. 40. № 2. С. 289–290.
Павлов А.В., Гаранин В.И., Бакиев А.Г., 2004. Обыкновенная гадюка Vipera berus (Linnaeus, 1758) // Змеи Волжско-Камского края. Самара: Издательство Самарского НЦ РАН. С. 49–61.
Справочник по климату СССР, 1965. Вып. 3. Ч. 2. Л.: Гидрометеоиздат. 343 с.
Andersson S., Johansson L., 2001. Cold hardiness in the boreal adder, Vipera berus // Cryo Letters. V. 22. P. 151–156.
Berman D.I., Bulakhova N.A., Alfimov A.V., Meshcheryakova E.N., 2016. How the most northern lizard, Zootoca vivipara, overwinters in Siberia // Polar Biology. V. 39. P. 2411–2425. https://doi.org/10.1007/s00300-016-1916-z
Bulakhova N.A., Meshcheryakova E.N., Korosov A.V., Ganjushina N.D., Berman D.I., 2017. Cold tolerance of the common adder, Vipera berus in Karelia // The 5th Biology of the Vipers Conference. Morocco. P. 46.
Costanzo J.P., Grenot C., Lee R.E.Jr., 1995. Supercooling, ice inoculation and freeze tolerance in the European common lizard, Lacerta vivipara // Journal of Comparative Physiology B. V. 165. № 3. P. 238–244.
http://alfa-prom.ru/e_besprovodniye_registratory.html Безбумажные беспроводные регистраторы (логгеры).
Lenders A.J.W., 2003. Overwinteringsplekken en voorjaarzonplekken van de adder in Nationaal Park De Meinweg // Natuurhistorisch Maandblad. V. 92. № 7. P. 181–189.
Storey K.B., Storey J.M., 2012. Insect cold hardiness: metabolic, gene, and protein adaptation // Canadian Journal of Zoology. V. 90. P. 456–475.
Viitanen P., 1967. Hibernation and seasonal movement of the viper, Vipera berus berus (L.) in Southern Finland // Annales Zoologici Fennici. V. 4. № 4. P. 472–546.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал