

Зоологический журнал, 2022, T. 101, № 1, стр. 94-100
Находки туруханской пищухи (Ochotona turuchanensis Naumov 1934) на Приморском хребте (Иркутская область)
Н. Г. Борисова a, *, А. А. Никулин a, Н. А. Никулина b, С. В. Попов a, А. И. Старков a, С. Ю. Ленхобоева a
a Институт общей и экспериментальной биологии СО РАН
670047 Улан-Удэ, Россия
b ФГБОУ ВО Иркутский ГАУ имени А.А. Ежевского
664038 Иркутск, Россия
* E-mail: nboris@list.ru
Поступила в редакцию 27.08.2020
После доработки 27.08.2021
Принята к публикации 28.08.2021
- EDN: DFQNIV
- DOI: 10.31857/S0044513422010020
Аннотация
В июне 2018 г. и июле 2019 г. были обследованы 14 локалитетов на Приморском хребте в Иркутской области, где согласно актуальным картам ареалов обитает только северная пищуха. По результатам сонографического анализа записанных звуковых реакций было установлено, что данные локалитеты населяет лишь туруханская пищуха (Ochotona turuchanensis Naumov 1934). Таксономическая принадлежность зверьков подтверждена с помощью анализа короткой последовательности цитохрома b, рисунков швов между предчелюстными и верхнечелюстными, между теменными и лобными костями черепа.
Ареал пищух группы alpina–hyperborea охватывает обширную территорию Северной Азии (рис. 1). В систематике этой группы до сих пор остается довольно много нерешенных вопросов как в силу специфики морфологической вариабельности внутри семейства, так и труднодоступности многих местонахождений зверьков. В 2002 г. Лисовский (Lissovsky, 2003), проведя многомерное шкалирование отдельно морфологических и совместно морфологических и биоакустических характеристик по большой выборке экземпляров алтайской и северной пищух, показал, что выборка распадается на 3 группы в обоих случаях. Одна группа по входившим в ее топотипам и типам была отнесена автором к северной пищухе (O. hyperborea Pallas 1811), другая – к алтайской (O. alpina Pallas 1773). Третья группа включала голотип и часть паратипов туруханской пищухи – таксона, впервые описанного Н.П. Наумовым в 1934 г. в качестве подвида северной пищухи на зверьках, отловленных в Эвенкии и отличающихся от других подвидов ярко-рыжей окраской шерсти по всему телу (Наумов, 1934). Зверьки этой группы по краниометрическим характеристикам перекрывались с северными пищухами, но имели количество хромосом (Формозов и др., 1999) и рисунок шва срастания верхнечелюстных и предчелюстных костей как у алтайских пищух и отличались от представителей обоих видов по признакам окраски шерсти и параметрам сигналов предупреждения об опасности. На основании полученных результатов Лисовский предложил выделить данную группу в новый вид – туруханскую пищуху (Ochotona turuchanensis) (Лисовский, 2002; Lissovsky, 2003). Позже было установлено, что по исследованным генам митохондриальной ДНК (COI и cytb) особи туруханской пищухи объединяются в кладу с алтайской пищухой, сестринскую по отношению к другим видам подрода Pika (Lissovsky et al., 2007); по комбинированным последовательностям 12 ядерных локусов туруханская также объединяется в одну кладу с алтайской, сестринскую по отношению к кладе, включающей северную, маньчжурскую и хэнтэйскую пищух, и к палласовой пищухе (Melo-Ferreira et al., 2015). Таксон имеет звуковой репертуар, элементы которого отличны от северной и алтайской пищух, но ближе к звуковой сигнализации алтайской пищухи (Лисовский, 2005). На сегодняшний день наряду с представлениями о видовом статусе туруханской пищухи (Lissovsky, 2003; Handbook of the Mammals …, 2016) существуют данные в пользу того, что это хорошо дифференцированный подвид или группа подвидов алтайской пищухи (Формозов и др., 1999; Формозов, Яхонтов, 2003; Формозов и др., 2006).
Рис. 1.
Ареалы северной, туруханской и алтайской пищух (с уточнениями). Вопросительными знаками отмечены местонахождения пищух группы видов alpina–hyperborea (Тавровский и др., 1971; Соколов и др., 1994) в зоне предполагаемого разрыва между ареалами туруханской и северной пищух (Lissovsky et al., 2007; Handbook of the Mammals …, 2016). Черная точка – территория, на которой расположены обследованные в работе локалитеты и которая в бо́льшем масштабе представлена на рис. 2.
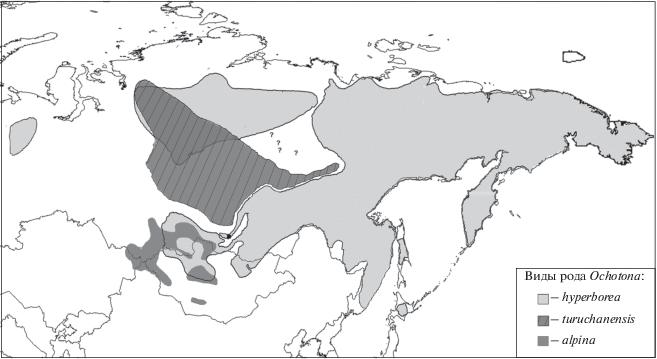
На рис. 1 показаны ареалы трех таксонов: алтайской, северной и туруханской пищух в соответствии с современными представлениями (Lissovsky et al., 2007; Handbook of the Mammals …, 2016). Ареал туруханской изолирован от ареала алтайской, на большей части территории Северной Азии соседствует с ареалом северной и перекрывается с ним в Эвенкии и на плато Путорана. Наше сообщение содержит факт обнаружения туруханской пищухи за пределами ранее установленного ареала, возможно, в зоне симпатрии с северной пищухой.
МАТЕРИАЛ И МЕТОДЫ
В период с 1 по 7 июня 2018 г. мы проводили отлов металлическими сплошными трапиковыми живоловками (7 × 7 × 25 см) на осыпи близ с. Новый Кочергат (рис. 2), где согласно картам ареалов (рис. 1) обитает северная пищуха. Одновременно записывали крики пищух в двух стационарных точках на осыпи и проводили наблюдения за зверьками утром (4:30–9:00) и вечером (18:00–22:00). На этом этапе были получены записи 120 сигналов, в том числе 48 песен, принадлежавших не менее чем 5 зверькам. Анализируя эти и более ранние записи, произведенные одним из авторов (А. Никулиным) на этой же осыпи в 2016–2017 гг., мы предположили, что имеем дело не с северной, а с туруханской пищухой. В 2019 г. провели обследование ряда локалитетов (рис. 2) в Голоустненском р-не Иркутской обл., где было известно обитание пищух (личные сообщения И.И. Медведева и Д.Ф. Леонтьева). Искали следы присутствия пищух (экскременты, стожки), слушали и записывали звуковые сигналы. Поскольку не по всем сигналам можно однозначно идентифицировать видовую принадлежность исполнителей, то в каждом локалитете слушали вокализации до тех пор, пока не удавалось записать или услышать звуки быстрой серии песни (рис. 3), которые позволяют четко отличать туруханскую от северной пищухи. С 1 по 7 июля 2019 г. проводили отлов живоловками Щипанова (Щипанов и др., 2008) размерами 10 × 10 × 25 см, наблюдения и регистрацию звуковых сигналов пищух на осыпях в 5 км к северу от стационара ИрГСХА Большие Мольты. В период с 7 по 10 июля 2019 г. были обследованы биотопы по левому берегу р. Нижний Кочергат и в ряде отдельных локалитетов. В общей сложности были проанализированы 112 песен от не менее чем 21 зверька из следующих локалитетов: местность Бурхай 52.1313° N 105.3263° E (1); 52.1305° N 105.3256° E (3); низовья р. Большие Мольты: правый берег 52.0583° N 105.2191° E (2); 52.0534° N 105.2223° E (6); 52.0534° N 105.2229° E (8); 52.0661° N 105.2033° E (14); среднее течение р. Нижний Кочергат: 52.0348° N 105.1318° E (7); 52.0513° N 105.2218° E (9); 52.0510° N 105.1221° E (11); правобережье р. Голоустная: ниже устья р. Колесма Морская 52.3720° N 105.3853° E (4); 52.3767° N 105.3876° E (5); окрестности села Нижний Кочергат 52.1580° N 105.2873° E (10); 52.0918° N 105.2559° E (12); 52.0827° N 105.2487° E (13).
Рис. 2.
Обследованные локалитеты (1–13, см. “Материал и Методы”) в Голоустненском р-не Иркутской обл.
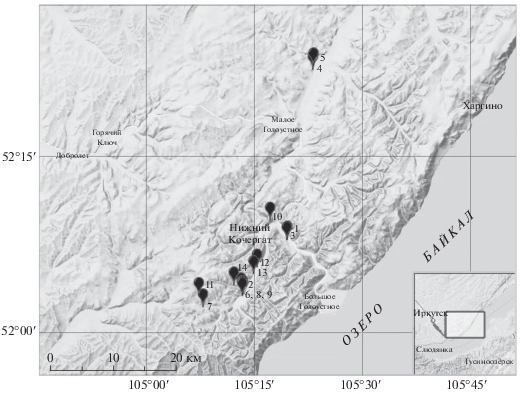
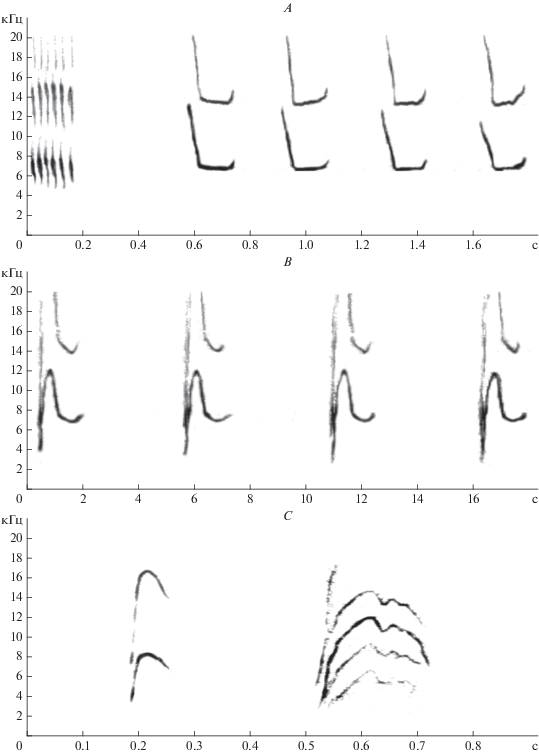
Рис. 3.
Звуковые сигналы пищух, записанные на осыпи в с. Нижний Кочергат: A – песня: трель и быстрая серия; B – песня: звуки медленной серии; C – сигналы “цик”, издаваемые самкой: первый – на канюка, второй – контекст неясен.
Туруханская пищуха, кроме как на осыпях, была обнаружена на сырых берегах речек и ручьев – в местах выхода камней и/или с наличием хорошо развитого мохового покрова. Одна особь обитала под полом деревянного гаража.
В 2018 г. была отловлена самка (локалитет 13), в 2019 г. – молодой зверек, пол которого не был установлен (локалитет 11). У обоих после обработки ушей лидокаином хирургическими ножницами отрезали небольшой фрагмент ушной раковины и помещали его в 95% спирт, в другое ухо вставляли пластиковую метку (Dalton Rototag) для идентификации. ДНК из фрагментов ушей выделяли стандартным хлороформно-фенольным методом. Амплификация коротких фрагментов cytb (650 и 470 п. н.) проведена с использованием праймеров и режимов ПЦР, указанных в работе Лисовского с коллегами (Lissovsky et al., 2007). Секвенирование проведено ЗАО “Евроген” (Москва, Россия), последовательности помещены в Генбанк под номерами MT513190 (молодой зверек из локалитета 11) и MT513191 (взрослая самка из локалитета 13). Для определения таксономической принадлежности отловленных зверьков построено филогенетическое дерево коротких фрагментов гена cytb (348 п. н.), полученных от отловленных особей и извлеченных из репозиториев NCBI (https://www.ncbi.nlm.nih.gov/) и Dryad (https://datadryad.org/), в MEGA 7 (Kumar et al., 2016), использован алгоритм neighbor-joining, модель Juke-Cantor, 1000 повторностей.
Спектрограммы звуковых сигналов получены на цифровом сонографе Raven Pro 1.6.1 (Center for Conservation Bioacoustics, 2019) с использованием следующих параметров: частота дискретизации 44.1 кГц, окно Kaiser, перекрывание по времени 50%, временно́е разрешение 5.8 мс, частотное разрешение 86.1 Гц.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Нами было записано несколько типов звуковых сигналов, один, согласно классификации Лисовского (Лисовский, 2005), представлял собой песни, состоящие в наиболее полном варианте из 4 компонентов: трели, затем звуки быстрой серии (рис. 3A), снова трели и звуки медленной серии (рис. 3B). Каждый элемент повторялся несколько раз. Быстрая и медленная серии звуков в отличие от трелей издавались и по отдельности. Следует отметить, что в период наблюдений меченые зверьки издавали только сигналы, названные исследователями пищух “предупреждающие об опасности сигналы” (Формозов, Никольский, 1979; Никольский, 1984; Формозов, 1991), далее по тексту называемые нами звукоподражательно “цик”. Сонограмма сигнала, поданного самкой на пролет и крики канюка поблизости от осыпи, приведена первой на рис. 3C. Второй сигнал на рис. 3C был подан самкой в отсутствие видимых причин для беспокойства. Все локалитеты, обследованные нами в 2018–2019 гг., судя по присутствию звуков быстрых серий песен, заселены туруханской пищухой. Мы ни разу не слышали песни северной пищухи.
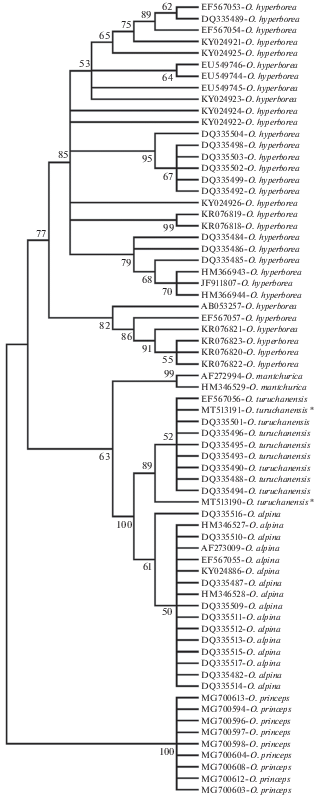
Данные анализа коротких фрагментов гена cytb двух отловленных особей подтверждают принадлежность пойманных зверьков к туруханской пищухе (рис. 4).
Рис. 4.
Топология филогенетического дерева пищух, построенного методом ближайшего соседа (NJ), на основании короткой последовательности гена cytb. На ветвях указаны значения индекса бутстрепа (1000 повторностей). Показаны только узлы, имеющие более 50% поддержки. Особи, отловленные на Приморском хребте, обозначены звездочкой.
Анализ рисунков швов между лобными и теменными костями (венечный шов), а также между верхнечелюстными и предчелюстными костями на четырех черепах пищух, добытых на осыпи близ с. Нижний Кочергат в 2016 г., показал, что они соответствуют описанным для туруханской пищухи (Lissovsky, Lissovskaya, 2002; Lissovsky, 2003): линия венечного шва неровная, U-образная; предчелюстные кости не образуют заметных парных выпячиваний к боковой стенке небного отверстия. Обследованные осыпи располагаются вблизи стационаров ИрГСХА, и в течение почти 50 лет проведения здесь полевых практик для студентов факультета охотоведения на этих осыпях постоянно регистрируется присутствие пищух. К сожалению, коллекция черепов зверьков из этих точек была уничтожена пожаром в музее ИрГСХА. Мы обследовали черепа, собранные в 50–60 гг. XX в. из ряда локалитетов долины р. Нижний Кочергат и хранящиеся в коллекции музея ИСиЭЖ (Новосибирск). Все они в соответствии с принятой во время коллектирования систематикой пищух были отнесены к северной пищухе. Согласно определительной таблице Лисовского (2002) по краниометрическим промерам и рисунку шва между верхнечелюстными и предчелюстными костями их следует отнести к туруханской пищухе. Исходя из этого, можно полагать, что данный таксон населяет локалитеты в долине р. Нижний Кочергат, по крайней мере, с 50-х годов прошлого столетия. Следует отметить, что в работе Лисовского (2002) использованы только промеры четырех черепов из локалитета Большие Мольты (указываемого в работе как пос. Мальта, Приморский хребет, по написанию локалитета в этикетках), но не оценен рисунок шва между верхнечелюстными и предчелюстными костями. Поскольку по размерным характеристикам невозможно различить туруханскую и северную пищух (Lissovsky, 2003), то, видимо, именно поэтому возникло представление, что на Приморском хребте обитает северная пищуха (Lissovsky et al., 2007).
Таким образом, в результате нашего исследования установлено обитание туруханской пищухи на Приморском хребте, в 300 км к югу от поселка городского типа Жигалово, возле которого расположена ближайшая точка подтвержденного обитания туруханской пищухи (Lissovsky, 2003). Обследованные нами точки находятся внутри участка, который ранее рассматривался как входящий в ареал северной пищухи. Обитают ли на данной территории северные пищухи и где расположены зоны контакта или симпатрии двух видов, если они есть, – остается открытым вопросом. Мы предполагаем, что вся территория к северу вплоть до Жигалово может быть заселена туруханской пищухой, так как с точки зрения ландшафтов и растительности она сходна с обследованной территорией (Д.Ф. Леонтьев, личное сообщение). То, что до сих пор данная территория рассматривалась как лишенная населения пищух (рис. 1), скорее всего, является следствием ее необследованности. Требуют уточнения юго-западная и восточная границы ареала туруханской пищухи.
В целом картина ареалов трех таксонов внутри рассматриваемой группы выглядит следующим образом: ареал северной пищухи охватывает подковой ареал подгруппы alpina–turuchanensis, при этом перекрывается в ряде локалитетов с ареалами обоих таксонов этой подгруппы. В то же время пока не найдено зон контактов ареалов алтайской и туруханской пищух. Такая картина предполагает более раннее расхождение между северной пищухой и общим предком alpinа–turuchanensis, более позднее – между alpinа и turuchanensis, что соответствует результатам анализа молекулярно-генетических данных (Melo-Ferreira et al., 2015): расхождение между предковой формой северной пищухи (давшей начало трем видам: hyperborea, mantchurica, hoffmanni) и предковой формой подгруппы alpina–turuchanensis датируется от 3 до 1.5 млн лет назад (в зависимости от разных дат калибровки), между алтайской и туруханской пищухами – от 0.6 до 0.3 млн лет назад. Группа рассматриваемых таксонов представляет собой интересную модель для выявления механизмов дивергенции на разных уровнях, для исследования процессов эволюции экологической ниши, поведения и т.п. Точная картина их ареалов необходима для разработки дизайна пространственно эксплицитного сбора молекулярно-генетических данных, поскольку только с помощью последних можно установить время образования тех или иных линий, центры их происхождения и пути расселения – параметры, базовые для понимания эволюции данной группы животных.
Список литературы
Лисовский А.А., 2002. Систематика пищух (Ochotona, Mammalia) группы alpina-hyperborea. Дис. … канд. биол. наук. М., МГУ. 134 с.
Лисовский А.А., 2005. Сравнительный анализ акустического репертуара и изменчивости звуковых сигналов пищух (Ochotona, Mammalia) группы alpina-hyperborea // Бюллетень Московского общества испытателей природы: Отдел биологический. Т. 110. Вып. 6. С. 12–26.
Наумов Н.П., 1934. Млекопитающие Тунгусского округа // Труды Полярной комиссии. Л.: Изд-во АН СССР. Вып. 17. С. 5–82.
Никольский А.А., 1984. Звуковые сигналы млекопитающих в эволюционном аспекте. М.: Наука. 200 с.
Соколов В.Е., Иваницкая Е.Ю., Груздев В.В., Гептнер В.Г., 1994. Млекопитающие России и сопредельных регионов: зайцеобразные. М.: Наука. 272 с.
Тавровский В.А., Егоров О.В., Кривошеев В.Г., Попов М.В., Лабутин Ю.В., 1971. Млекопитающие Якутии. М.: Наука. 659 с.
Формозов Н.А., 1991. Звуковая сигнализация пищух (Ochotona, Lagomorpha): географический, систематический и экологический аспекты. Автореф. канд. … биол. наук. М. 24 с.
Формозов Н.А., Никольский А.А., 1979. Положение уральской пищухи в группе alpina (биоакустический анализ) // Млекопитающие Уральских гор. Информ. материалы. Свердловск: УНЦ АН СССР. С. 80–82.
Формозов H.А., Лисовский А.А., Баклушинская И.Ю., 1999. Кариологическая диагностика пищух (Ochotona, Lagomorpha) плато Путорана // Зоологический журнал. Т. 78. № 5. С. 606–612.
Формозов H.А., Яхонтов Е.Л., 2003. Зона симпатрии алтайской (Ochotona alpina) и северной (O. hyperborea) пищух на плато Путорана с описанием Ochotona hyperborea naumovi ssp. // Зоологический журнал. Т. 82. № 4. С. 485–496.
Формозов H.А., Григорьева Т.В., Сурин В.Л., 2006. Молекулярная систематика пищух подрода Pika (Ochotona, Lagomorpha) // Зоологический журнал. Т. 85. № 2. С. 1465–1473.
Щипанов Н.А., Литвинов Ю.Н., Шефтель Б.И., 2008. Экспресс-метод оценки локального биологического разнообразия сообщества мелких млекопитающих // Сибирский экологический журнал. Т. 5 (10). С. 783–791.
Center for Conservation Bioacoustics, 2019. Raven Pro: Interactive Sound Analysis Software (Version 1.6.1) Ithaca, NY: The Cornell Lab of Ornithology [Электронный ресурс].
Handbook of the Mammals of the World. Vol. 6: Lagomorphs and Rodents I, 2016. Wilson D.E., Lacher T.E., Mittermeier R.A. (Eds). Barcelona: Lynx Edicions. 987 p.
Kumar S., Stecher G., Tamura K., 2016. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets // Molecular Biology and Evolution. V. 33. P. 1870–1874.
Lissovsky A.A., 2003. Geographical variation of skull characters in pikas (Ochotona, Lagomorpha) of the alpina-hyperborea group // Acta Theriologica. V. 48. № 1. P. 11–24.
Lissovsky A.A., Ivanova N.V., Borisenko A.V., 2007. Molecular phylogenetics and taxonomy of the subgenus Pika (Ochotona, Lagomorpha) // Journal of Mammalogy. V. 88. № 5. P. 1195–1204.
Lissovsky A.A., Lissovskaya E.V., 2002. Diagnostics of pikas (Lagomorpha, Ochotonidae, Ochotona) from the Putorana Plateau, Eastern Siberia // Russian Journal of Theriology. V. 1. № 1. P. 37–42.
Melo-Ferreira J., Lemos de Matos A., Areal H., Lissovsky A.A., Carneiro M., Esteves P.J., 2015. The phylogeny of pikas (Ochotona) inferred from a multilocus coalescent approach // Molecular Phylogenetics and Evolution. V. 84. P. 240–244.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал