

Зоологический журнал, 2022, T. 101, № 1, стр. 101-116
Структурные особенности кожи, волос и специфических кожных желез обыкновенного хомяка (Cricetus cricetus, Cricetidae, Rodentia)
О. Ф. Чернова a, *, Р. М. Хацаева a, **, В. П. Куприянов a, ***, Н. Ю. Феоктистова a, ****, А. В. Суров a, *****
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: chernova@sevin.ru
** E-mail: r.khatsaeva@mail.ru
*** E-mail: meahn@mail.ru
**** E-mail: feoktistovanyu@gmail.com
***** E-mail: allocricetulus@gmail.com
Поступила в редакцию 29.05.2021
После доработки 16.08.2021
Принята к публикации 17.08.2021
- EDN: DXALLZ
- DOI: 10.31857/S0044513422010044
Аннотация
Впервые с использованием комплексного методического подхода (морфометрия, гистология, световая и сканирующая электронная микроскопия) исследовали строение кожно-волосяного покрова обыкновенного хомяка (Cricetus cricetus), вида, отнесенного к категории CR (Critically Endangered, IUCN) Красной книги МСОП. Показано, что пестрый окрас типичной трехцветной формы этого вида определяется разнообразием цвета волос разных категорий – от черных и белых направляющих до зонарно окрашенных, однотонных или бесцветных остевых и пуховых. Волосы на дорсальной стороне тела более густые, располагаются неровными рядами небольших “ложных” пучков – фолликулы разных волос залегают на разной глубине разрозненно, а стержни собираются в общий пучок только в волосяной воронке. Рассмотрены особенности строения парных боковых (БЖ) и непарной среднебрюшной (СБЖ) желез. БЖ представляют собой железистое поле, а СБЖ – мешотчатый орган. Волосы на железах обоих типов деформированы, что предполагает постоянное их травмирование в процессе маркировочного поведения, лизис, выпадение и включение в состав секрета. Мелкие темные волоски центральной полости СБЖ имеют кутикулярные пазухи, в которых скапливается пахучий секрет железы. В обоих типах желез впервые у этого вида обнаружены мерокриновые гепатоидные железы, которые добавляют в сальный секрет белковую составляющую. Голо-мерокриновая секреция, соединительнотканная и мышечная строма БЖ и СБЖ различаются в деталях, что позволяет предполагать различия химического состава железистой паренхимы и производимых ею секретов. Обсуждаются структурные особенности волос и специфических кожных желез, а также их роль в химической коммуникации обыкновенного хомяка.
В настоящее время изучение морфофизиологических адаптаций, в том числе кожно-волосяного покрова редких и исчезающих видов, чрезвычайно актуально. К таким видам относится и объект нашего исследования – обыкновенный хомяк (Cricetus cricetus Linnaeus 1758) – единственный представитель рода Cricetus Leske 1779 (Wilson, Reeder, 2005). До 1970-х годов он был многочислен и серьезно вредил сельскому хозяйству практически на всем своем огромном ареале (около 6 млн км2). В отдельных регионах плотность его поселений достигала 80 особей/га (Ružić, 1977; Nechay, 2000). Однако в настоящее время численность обыкновенного хомяка резко сократилась, вплоть до полного исчезновения в некоторых европейских странах (Surov et al., 2016). В 2020 г. эксперты Международного союза охраны природы (IUCN), оценив состояние различных популяций обыкновенного хомяка, подняли его природоохранный статус сразу на четыре ступени, до “находящийся под угрозой исчезновения” (CR – “Critically Endangered”) (Banaszek et al., 2020). Поэтому любые знания о биологии вида приобретают особую значимость.
Сведения о морфологии кожно-волосяного покрова и специфических кожных желез C. cricetus отражены в ряде работ прошлого века (Vrtiš, 1930, 1930а; Воронцов, Гуртовой, 1959; Kourist, 1959; Reznik et al., 1979; Соколов и др., 1994). Ранее нами было детально изучено строение кожи, волос и желез у представителей ряда других родов подсемейства Cricetinae: Phodopus Miller 1910 (Феоктистова, 2008), Cricetulus Milne-Edwards 1867 и Allocricetulus Argiropulo 1932 (Чернова и др., 2021), что дает возможность провести сравнительно-морфологический анализ этих структур.
Обыкновенный хомяк – самый крупный представитель подсемейства Cricetinae. Характерной чертой, отличающей типичных Cricetus cricetus от других хомяков, да и вообще грызунов России, является выраженная трехцветность. Спина охристо-коричневого цвета, на боках – два больших светлых пятна, обычно разделенных участком черного меха, по одному большому светлому пятну имеется также на каждой боковой стороне головы и по одному маленькому пятнышку – за каждым ухом. У некоторых особей есть белые пятна и в лопаточной области, и на бедрах. Брюхо полностью окрашено в черный цвет. Хомяк отлично использует это при оборонительном поведении: встаёт на задние лапы, разводит передние, издаёт громкие резкие звуки и демонстрирует черное брюхо, пугая соперника. Однако помимо стандартной трехцветной окраски у обыкновенного хомяка в природе встречается множество цветовых вариаций: белые, альбиносы, желтые, красноватые, песочные, серые, двухцветные (без светло-желтого), но чаще – почти целиком черные особи (белыми остаются только горло, нос, уши и кончики лап (Kryštufek et al., 2020; Крускоп, 2002). В некоторых частях ареала такие частичные меланисты могут доминировать или даже полностью замещать типично окрашенных зверьков (Кириков, 1934).
Волосяной покров у C. cricetus густой и мягкий, укороченный, но с сохранением ворса, направленного спереди назад. Подпушь – голубовато-серая. Несмотря на многочисленные сведения о пестром окрасе C. cricetus, микроскопические особенности пигментации волос разных категорий у этого вида остаются мало изученными.
У обыкновенного хомяка на спине хорошо развиты грубые остевые волосы длиной 25–30 мм и толщиной 60–85 мкм, которые характеризуются цилиндрическими или уплощенными стержнями с лестничной сердцевиной. Подшерсток состоит из тонких (10–25 мкм) и более толстых (25–55 мкм) волосков, стержни которых уплощены в дорсо-вентральном направлении и на срезе имеют форму эллипса (Galatík et al., 2011).
Вибриссы прямые, жесткие и длинные (32− 39 мм), коричневые, толщиной 166−191 мкм, располагаются по 30 штук с каждой стороны морды четырьмя или пятью рядами, а светлые вибриссы короче (7.5–25.1 мм) (Kourist, 1957; Reznik et al., 1979).
Таким образом, данные о структуре волос C. cricetus все еще фрагментарны, особенно это касается типологии, микроструктуры и внутреннего строения волос на разных участках тела. Можно предположить, что разнообразие волос обеспечивает ярусность шерсти, а сами волосы специфичны на разных участках тела зверька по размерам и, возможно, по внутреннему строению, особенно, если они ассоциированы со специфическими железами, выполняющими разнообразные жизненно важные функции.
У С. cricetus имеются многочисленные специфические кожные железы: Гардеровы, мейбомиевы, железы наружного слухового прохода, углов рта, парные боковые железы (БЖ), непарная среднебрюшная железа (СБЖ), препуциальные, циркуманальные, железы крайней плоти, клитора, подошвенные. Апокриновые потовые железы отсутствуют (Schaffer, 1940; Brown, Williams, 1972; Бодяк, 1994; Соколов, Чернова, 2001).
Наиболее значимыми являются БЖ и СБЖ, выделяющие пахучий секрет (см. Schaffer, 1940; Воронцов, 1982; Соколов, Чернова, 2001). Причем среди Cricetinae железы обоих типов хорошо развиты еще только у представителей Allocricetulus, что подтверждает филогенетическую близость родов Allocricetulus и Cricetus (Neumann et al., 2005, 2006; Lebedev et al., 2018). У изученных видов других родов подсемейства присутствует либо один, либо другой тип желез. Так, у хомяков рода Mesocricetus Nehring 1898 описаны только БЖ (Märkel, 1952), тогда как у представителей рода Phodopus – CБЖ (Феоктистова, 2008), что не согласуется с утверждением Воронцова и Гуртового (1959) об одновременном присутствии СБЖ и БЖ у всех представителей подсемейства.
БЖ у C. cricetus были впервые обнаружены и описаны, по-видимому, Зульцером (Sultzer, 1774, цит. по: Vrtiš, 1930). Позднее Вртиш (Vrtiš, 1930, 1930a) более детально описал расположение, строение, возрастные и сезонные особенности развития и функционирования этих желез. Обнаружено, что БЖ особенно активны у взрослых самцов, которые метят этим секретом свою территорию, маркируют объекты и субстрат (Schaffer, 1940; Eibl-Eibesfeldt, 1953). Тем не менее до настоящего времени не было проведено комплексного описания БЖ на разных уровнях структурной организации, не описана микроструктура эпидермиса, дермы и волос, ассоциированных с этими железами.
СБЖ (или “пупочный орган”) у обыкновенного хомяка также хорошо развита. Внешне она представляет собой оголенный участок кожи размером 5 × 5 мм, в центре которого располагается выводное отверстие. Это отверстие сверху прикрыто кожей и ведет в резервуар длиной примерно 5−10 и шириной 3−5 мм. В середине возвышается гребень, с обеих сторон которого растут группы черных жестких волос длиной 2 мм, направленные каудально. По структуре СБЖ это – многодольчатое сальное образование, снабженное системой ветвящихся выводных протоков, оболочкой из поперечнополосатой мускулатуры и пучками гладкой мускулатуры, способствующей выведению секрета (Vrtiš, 1930 − цит. по: Schaffer, 1940, p. 212).
У взрослых особей под эпидермисом и между ацинусами БЖ и СБЖ имеются скопления меланина, что отличает их от неполовозрелых особей (Reznik et al., 1978, 1979).
Принимая во внимание природоохранный статус С. cricetus, очевидным представляется важность изучения разных аспектов биологии этого вида и, в частности, строения его кожно-волосяного покрова и специфических кожных желез.
Цель работы состоит в сравнительно-морфологическом и морфофункциональном анализе кожного покрова и специфических кожных желез у C. cricetus.
МАТЕРИАЛ И МЕТОДЫ
Объектом наших исследований послужили половозрелые (полуторагодовалые) самец № 1 и самка C. cricetus, добытые соответственно в Воронежской обл. (24.06.2019 г.) и Крыму (12.07.2019 г.), а также самец № 2, добытый в Пензенской обл. в июле 2021 г. (табл. 1). Исследование проведено на зверьках с типичным трехцветным окрасом. Зверьков подвергли эвтаназии на аппарате Плинаркон-5 (ООО “ПрофМТ”, Санкт-Петербург) и взяли пробы кожи и волос с холки, крестца, бока, брюха, провели экстирпацию БЖ и СБЖ. Кроме того, были проведены промеры БЖ и СБЖ у живых обыкновенных хомяков обоих полов разного возраста (от 2−4 мес до годовалых) при их отлове на экспериментальной площадке в г. Симферополе.
Таблица 1.
Стандартные промеры особей обыкновенного хомяка (Cricetus cricetus), использованных в исследовании
| Пол, возраст, № | Масса тела, г | Длина, мм | Длина, мм | |||||
|---|---|---|---|---|---|---|---|---|
| L | Cd | Pl | Au | Семенники | БЖ | СБЖ | ||
| Самец, ad. № 1 | 520 | 350 | 50 | 35 | 30 | 30 | 26.0 | 7.5 |
| Самец, ad. № 2 | 501 | 349 | 49 | 36 | 29 | 25 | 23.0 | 6.9 |
| Самка, ad. | 390 | 300 | 55 | 38 | 22 | – | 21.0 | 6.0 |
Работа выполнена на оборудовании Центра коллективного пользования “Инструментальные методы в экологии” ИПЭЭ РАН, с использованием новейших методов гистологии, морфометрии, световой микроскопии и сканирующей электронной микроскопии (СЭМ).
Морфологическая терминология описания волос и их структур соответствует общепринятой (Соколов, 1973; Чернова, Целикова, 2004). Выделенные категории волос включают: направляющие (самые длинные и толстые, цилиндрические, образующие ворс шерсти), остевые и пуховые волосы нескольких размерных порядков в зависимости от размеров и конфигурации стержня. Термин “дифференциация волосяного покрова” обозначает подразделение массы шерсти на несколько ярусов и наличие волос разных категорий и порядков.
Для изучения в СЭМ пробы волос дорсальной и латеральной поверхности кожи, БЖ и СБЖ фиксировали в 10% нейтральном формалине (Соколов и др., 1988): очищали в шампуне, промывали в дистиллированной воде, проводили через спирты возрастающей концентрации (от 30 до 100%) и ацетон, высушивали в критической точке на установке Hitachi Critical Point Dryer HCP-1 (“Hitachi”, Япония). Далее напыляли золотом в вакууме на установке S150A Sputter Coater (“Edwards”, Великобритания). Препараты изучали в растровых электронных микроскопах Vega TS5130MM (“Tescan”, Чехия) и JSM 840A (“JEOL”, Япония), получая электронограммы орнамента кутикулы и внутреннего дизайна (архитектоники) волос.
Для гистологического изучения пробы кожи фиксировали в 10% нейтральном формалине, получали парафиновые срезы толщиной 5−15 мкм, окрашивали гематоксилином Эрлиха и эозином (Соколов и др., 1988) с использованием полуавтоматического специализированного гистологического оборудования: гистопроцессор TPC-15, заливочная станция TES-99, микротом Meditome M 530 (“Medite”, ФРГ). Препараты изучали на цифровом флуоресцентном микроскопе Keyence Biorevo BZ-9000 (“Eurotek Inc.”, США, Германия, Россия, Польша).
Микроструктуру анализировали в программе “ATLAS” (Tescan, Чехия) и “ImageJ” (Wayne Rasband, США). Полученные морфометрические данные статистически обрабатывали в программе “STATISTICA 10” (США).
Микрофотографии редактировали в программе “Adobe Photoshop Element 11” (“Adobe”, США), но изменения касались лишь компоновки, яркости и контраста изображений.
РЕЗУЛЬТАТЫ
Волосяной покров. Пестрый окрас шерсти на дорсальной и боковой поверхностях тела обыкновенных хомяков обеспечивается за счет зонарного окраса остевых волос, у которых нижняя часть стержня светло-коричневая, срединная – темно-коричневая, а апикальный кончик – светло-коричневый (или бурый). Черные направляющие волосы незначительно выступают над ярусом остевых волос и пуха. Пуховые волосы темно-коричневые по всему стержню или бесцветные. Область расположения БЖ хорошо различима, благодаря более темному цвету волос, окружающих железистое поле. Шерсть на брюхе выглядит черной. Ости без зонарного окраса, темно-коричневые, а пуховые волосы темно-серые или бесцветные. СБЖ также прикрыта темными волосами и хорошо заметна, благодаря раскрытому выводному протоку.
Поверхность рогового слоя эпидермиса шероховатая или разрыхленная (особенно на боку), большей частью гладкая или слегка волнистая и даже складчатая (на боку складки вытянуты перпендикулярно росту волос) с хорошо выраженными многочисленными отслаивающимися роговыми чешуйками и их обломками, границы между которыми различимы (рис. 1A). Округлые чешуйки с волнистыми приподнятыми краями не цельные, а разделены складками на треугольные сектора, число которых может достигать шести (рис. 1A, врезка). Плотные пучки волос выходят из волосяных воронок, каждый пучок может содержать до 20 волосков (рис. 1B). Надо отметить, что волосы всех категорий имеют тонкое и длинное основание – “ножку”, поэтому различить их по морфометрии на уровне нижней трети стержня затруднительно.
Рис. 1.
Общий вид поверхности кожи и растущих на ней пучками волос на разных участках тела взрослого самца обыкновенного хомяка (Cricetus cricetus): A – кожа и волосы холки, врезка – разделенная на сектора чешуйка рогового слоя; B − пучок волос на боку; С − железистое поле левой боковой железы, врезка − свободный от волос проток железы с гранулами секрета, указан стрелкой; D − кутикула волос боковой железы; E − среднебрюшная железа; F − кутикула волос среднебрюшной железы. CЭМ. Масштаб: A − 10 мм, врезка − 200 мкм; B, F – 100 мкм; C − 2 мм, врезка − 200 мкм; D − 20 мкм; E − 1 мм.
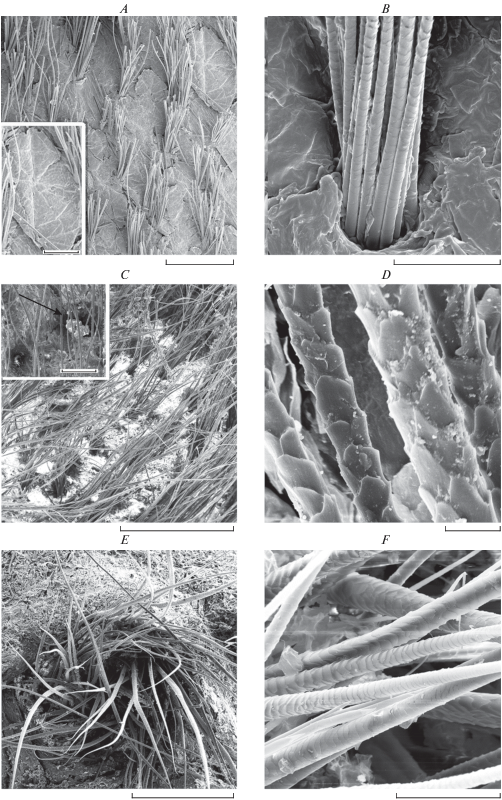
Дифференциация шерсти относительно слабая, сходная на разных участках тела, незначительно отличается в паховой области брюха: волосы растут пучками, имеются относительно длинные и толстые темные направляющие волосы, остевые волосы нескольких размерных порядков и многочисленные пуховые волосы (табл. 2, рис. 2A, 2B). Шерсть на спине и боках гуще, чем на брюхе. На 1 см2 кожи на холке, спине и боках в среднем располагается около 100 пучков волос, а на брюхе – около 85 пучков. На спине такой пучок включает один толстый направляющий волос, один или два остевых волоса и 13–16 пуховых волос (табл. 2). Пучок на брюхе отличается меньшим количеством пуховых волос.
Таблица 2.
Морфометрия волос из разных топографических участков тела взрослого самца обыкновенного хомяка (Cricetus cricetus)
| Показатель | Участок тела | |||||
|---|---|---|---|---|---|---|
| холка | спина | бок | брюхо | БЖ | СБЖ | |
| Число пучков волос на 1 см2 кожи | 107 | 105 | 103 | 85 | 55 | Растут поодиночке |
| Число волос на 1 см2 кожи* | 2140 | 2100 | 1751 | 1255 | 1100 | 84 |
| Категория волос, толщина M ± m | ||||||
| Направляющие | 91.8 ± 5.1 σ = 11.5 |
92.9 ± 4.1 σ = 9.2 |
87.0 ± 4.1 σ = 9.1 |
84.5 ± 0.5 σ = 4.5 |
90.3 ± 3.3 σ = 7.8 |
81.7 ± 0.2 σ = 0.3 |
| Ость I | 60.8 ± 2.9 σ = 6.6 |
60.8 ± 2.9 σ = 6.6 |
63.2 ± 2.9 σ = 6.6 |
45.0 ± 0.5 σ = 1.5 |
59.1 ± 0.4 σ = 0.7 |
57.9 ± 2.5 σ = 3.6 |
| Ость II | 40.6 ± 2.7 | 42.8 ± 2.2 σ = 9.2 |
46.6 ± 3.3 σ = 7.4 |
– | 45.2 ± 2.4 σ = 0.7 |
– |
| Пух I | 25.5 ± 2.2 σ = 4.9 |
21.8 ± 1.2 σ = 2.7 |
20.6 ± 1.6 σ = 3.3 |
23.0 ± 0.6 σ = 0.1 |
24.5 ± 0.05 σ = 0.1 |
– |
| Пух II | 13.8 ± 1.5 σ = 3.0 |
15.8 ± 1.2 σ = 2.7 |
11.8 ± 0.8 σ = 1.8 |
11.3 ± 0.6 σ = 0.1 |
16.9 ± 0.05 σ = 0.1 |
– |
| Пух Ш | 6.4 ± 0.4 σ = 0.9 |
6.2 ± 0.2 σ = 0.4 |
6.0 ± 0 σ = 0 |
6.0 ± 0 σ = 0 |
6.0 ± 0 σ = 0 |
– |
Рис. 2.
Строение волосяных пучков, микроструктура волос взрослого самца обыкновенного хомяка (Cricetus cricetus). A − на спине. В − на брюхе. C − поперечные срезы оснований волос разных категорий из одного пучка. Направляющий волос: D − поперечные срезы в середине стержня. E − орнамент кутикулы от корня до середины стержня (слева направо). F − продольные срезы основания и середины стержня (слева направо). Пуховой волос: G − орнамент кутикулы. Сужение стержня указано стрелкой. 1 − направляющий волос, 2 − ость I, 3 − ость II, 4 − пучок пуховых волос, 5 − волосяная воронка. Схема (A, B) , СЭМ (C−G). Масштаб: A, B −1 мм; C−G − 10 мкм.
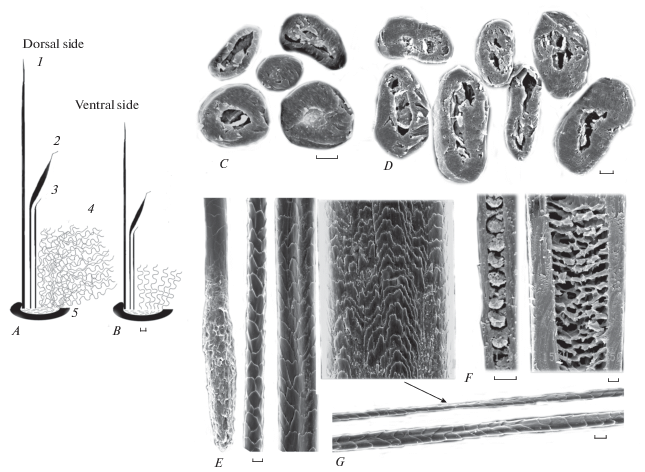
Шерсть трехъярусная: верхний (высотой 26 мм на спине и 21 мм на брюхе) ярус состоит из прямостоящего ворса (темных направляющих волос), средний ярус (соответственно 15 и 8 мм) – из остевых волос двух порядков (ость I и ость II), причем у более длинной и толстой ости I расширенная часть стержня (гранна) незначительно изогнута. Нижний ярус (подпушь) представлен многочисленными длинными (на спинной и боковой поверхностях тела) пуховыми волосами трех порядков (в зависимости от толщины стержня) или еще более короткими (с четырьмя волнами) пуховыми волосами (на брюхе) (рис. 2A, 2B). На спине подпушь примерно наполовину ниже ворса за счет того, что относительно длинные (до 20 мм в расправленном виде) пуховые волосы свойлачиваются (волнообразно извиваясь, переплетаясь, уплотняясь, образуют прилегающий к коже слой, подобный войлоку). На брюхе волосы всех категорий прямостоящие, а длина пуховых волос не превышает 6 мм.
Направляющие волосы темные, прямые, жесткие. Длинный корень постепенно переходит в тонкое и длинное основание, которое постепенно расширяется к верхней части цилиндрического стержня, хотя гранны ‒ резкого локального расширения не имеют, снабжены узкой, заостренной, немного изогнутой вершиной. В средней, немного уплощенной (рис. 2D) части стержня, по его вентральной стороне проходит неглубокая бороздка (рис. 2E). Сильно пигментированная сердцевина развита очень хорошо (составляет до 80% толщины стержня), но отсутствует в вершине стержня. В тонком основании стержня она однорядная лестничная, а в расширенных верхних отделах – сетчатая (рис. 2F). Пигментные гранулы в дисках немногочисленные. Орнамент кутикулы изменяется по стержню (рис. 2E). В тонком основании стержня орнамент кольцевидный (одна чешуйка оборачивает стержень), чешуйки высокие с гладким, имеющим одну впадину или треугольный вырост апикальным краем. Место смыкания обеих половинок чешуйки хорошо заметно. Выше по стержню кутикула грубая, многослойная, с хаотичным орнаментом из полиморфных изломанных чешуек, которые в самом расширенном участке стержня незначительно уплощаются, имеют треугольные выросты и сильно изрезанные свободные края (рис. 2E). Поперек стержня укладываются три-четыре чешуйки.
Ости I – светлые или темные, слабо пигментированные, имеют тонкое основание и расширенную верхнюю треть стержня (гранну), заканчивающуюся острой изогнутой вершиной (рис. 2A, 2B). Стержень может сгибаться под острым углом в области перед гранной, прикрывая нижележащий ярус. Сердцевина хорошо развита (до 60% толщины стержня). Кутикула сходна с таковой у направляющих волос, но более гладкая и менее травмированная.
Ости II немного короче, тоньше, слабее пигментированы, с менее развитой сердцевиной (45%), чем ости I.
Пуховые волосы трех категорий, тонкие слегка волнистые (рис. 2A, 2B), с сужением стержня в местах изгибов волн (рис. 2G, стрелка), слабо пигментированные (желтовато-серые) или бесцветные. Наиболее крупные пуховые волосы имеют развитую сердцевину (до 30% толщины стержня), а самые тонкие волоски (толщиной 6 мкм) вообще лишены сердцевины.
Таким образом, строение сердцевины изменяется в зависимости от толщины волос, но всегда одно-, двухрядное лестничное или сетчатое. Степень развития сердцевины закономерно возрастает в ряду от пуховых волос до остевых, а затем направляющих волос, у которых число рядов сердцевинных полостей, расположенных цепочками поперек стержня, может достигать семи. Хаотичный орнамент кутикулы обнаружен у направляющих и остевых волос разных порядков, а сами чешуйки полиморфны.
Абсолютная высота чешуек колеблется на разных участках стержня волос: у направляющих волос от основания до середины – от 18 до 21 мкм, у остевых – от 21 до 8 мкм, у пуха – от 16 до 14 мкм. Сравнение относительной высоты чешуек по Индексу кутикулы (отношение толщины волоса к высоте чешуйки) показывает, что у направляюших и остевых волос чешуйки гораздо выше в основании стержня (0.6–0.9), а выше по стрежню уплощаются (от 1.6–2.0 до 6.5–2.0).
Кожа и специфические кожные железы. Кожа на разных участках тела различается незначительно. На дорсальной и боковой поверхностях она непигментированная, складчатая, средней толщины (1.2–1.6 мм), с тонким (56–68 мкм, промер без рогового слоя) складчатым эпидермисом, покрытым относительно толстым (до 68 мкм), интенсивно десквамирующим роговым слоем. Дерма не подразделяется на четкие слои и состоит из сосочкового слоя (составляет до 90% дермы). Нижняя граница этого слоя проходит на уровне залегания луковиц волос, растущих пучками. Пучки волос располагаются неровными рядами. Мышцы, поднимающие волосы, не обнаружены. Сальные железы размером 40.6 ± 4.5 × 90.2 ± ± 6.26 мкм (n = 5) в числе двух-трех окружают пучок волос. Утолщенные пучки коллагеновых волокон образуют хаотичное переплетение. Подкожная жировая клетчатка хорошо развита (толщиной до 0.5 мм), состоит из многочисленных жировых клеток (42.8 ± 2.3 × 58.6 ± 4.3 мкм, n = 5).
Боковые железы (glandulae lateralis) расположены симметрично по бокам тела в задней трети туловища, каждая из них представляет собой железистое поле удлиненно-овальной формы, вытянутое в краниально-каудальном направлении и окруженное кожным валиком так, что само железистое поле залегает глубже (напоминает корытце). БЖ имеются у обоих полов, их абсолютные и относительные размеры крупнее у самцов, чем у самок, а также у взрослых особей по сравнению с молодыми (табл. 1, 3).
Таблица 3.
Продольная длина боковых и среднебрюшной желез у разновозрастных особей обыкновенного хомяка (Cricetus cricetus) (M ± m)
| Пол | Возраст | Длина тела, мм |
БЖ | СБЖ | ||
|---|---|---|---|---|---|---|
| Число особей | Длина железы, мм | Число особей | Длина железы, мм | |||
| Самцы | subad. | 227.6 ± 4.8 | 50 | 20.2 ± 0.4 | 104 | 5.8 ± 0.2 |
| ad., возраст ≥ 1 год | 258.4 ± 6.1 | 33 | 26.7 ± 0.7* | 85 | 7.8 ± 0.2 | |
| Самки | subad. | 219.0 ± 3.8 | 28 | 17.4 ± 0.5 | 57 | 5.2 ± 0.5 |
| ad., возраст ≥ 1 год | 254.3 ± 7.2 | 18 | 21.5 ± 0.8** | 21 | 6.4 ± 0.3 | |
Поверхность железистого поля непигментированная, с шероховатым десквамирующим эпидермисом, покрытым обильным сальным секретом, обломками корнеоцитов, с редкими и мягкими волосами (рис. 1С, 1D). На нем хорошо различимы волосяные воронки с пучками волос и свободные от волос самостоятельные довольно крупные (диаметром до 196 мкм) протоки БЖ с изливающимся секретом (рис. 1С, вставка, стрелка). Плотность волосяного покрова на железе почти вдвое меньше, чем на спине (табл. 2). Волосы на БЖ растут пучками по 3–15 в каждом или поодиночке, они густо покрыты секретом. Одиночные волоски очень короткие (до 5 мм), с тонкой цилиндрической (рис. 3В) прикорневой зоной и деформированным изогнутым стержнем, заканчивающимся тонкой и длинной вершиной (рис. 3A). Сердцевина в этих волосах развита хорошо, но ее тяж деформирован (рис. 3С). Орнамент кутикулы сходен с таковым у обычных остевых волос (рис. 1D, 3D). В верхних отделах стержня кутикула грубая, толстая и изломанная. Индекс кутикулы изменятся от 1.4 в основании стержня до 2.0 выше основания и 3.6 в середине стержня, а абсолютная высота чешуек соответственно равна 19.0 ± 2.9, 19.0 ± 1.0 и 21.0 ± 1.9 мкм.
Рис. 3.
Конфигурация и микроструктура волос специфических кожных желез взрослой самки обыкновенного хомяка (Cricetus cricetus). Боковая железа: A − конфигурация стержня. B − поперечные срезы основания стержня. C − то же середины стержня. D − орнамент кутикулы от основания до середины стержня (слева направо). Среднебрюшная железа: E − конфигурация стержня. F − орнамент кутикулы от основания до вершины стержня (слева направо). G − фрагмент вершины стержня с пазухами между чешуйками. СЭМ. Масштаб, мкм: A, E − 100; B−D, F, G − 10.
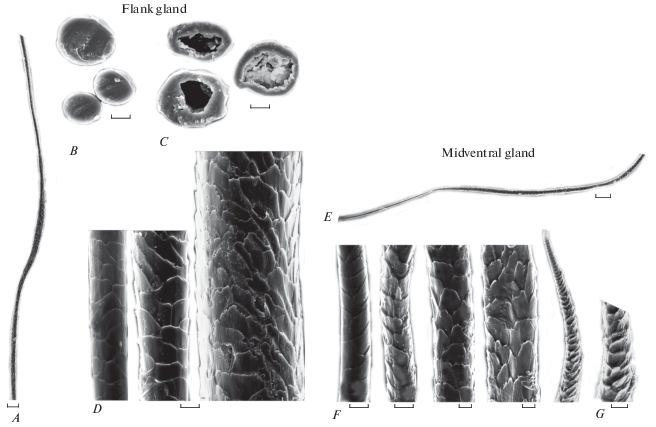
Орнамент кутикулы полукольцевидный (одна чешуйка не полностью оборачивает стержень), отличается у ости и пуха в основании стержня (рис. 3D). У остей поперек стержня укладывается по 2.5 копьевидной чешуйки, а у пуховых волос – по 1.0–1.5 чешуйки с округло-удлиненным апикальным концом. Высота чешуек в основании стержня изменяется от 19 до 21 мкм у волос разных категорий, и, судя по индексу кутикулы, они относительно крупнее у пуха (0.7–1.4), чем у остей (2.9–3.1). Конфигурация свободного апикального края чешуек гладкая, слабоволнистая. Раструбная (с сильно отодвинутыми от стержня краями) и ребристая кутикула не обнаружена.
Общий план строения кожи БЖ вполне типичен для железистых полей (рис. 4А). Эпидермис вместе с сосочковым слоем формирует сильно складчатую поверхность кожи, заполненную слоистым секретом (рис. 4С) и пронизанную выводными протоками БЖ, волосяными луковицами, растущими поодиночке или пучками. Пигментация имеется лишь в крупных волосяных сумках, но не в дермальном слое. Основную толщину кожи (до 70%) занимает железистая паренхима, подстилаемая снизу подкожной жировой клетчаткой. Клетчатка проникает и в кожные валики, окружающие железистое поле. Глубже клетчатки располагается нетолстый (до 0.7 мм) слой поперечнополосатой мускулатуры.
Рис. 4.
Микроструктура боковой железы взрослых самцов № 1 (А, B) и № 2 (C, D) обыкновенного хомяка (Cricetus cricetus) на трансверсальных (A, B) и сагиттальных (С, D) срезах. 1 − секреторная альвеола, 2 − проток-резервуар, 3 − слоистый секрет, 4 − соединительнотканная строма, 5 − капилляр, 6 − волосяной фолликул, 7 − гепатоидная железа, 8 − проток гепатоидной железы, 9 − эпидермис, 10 − дерма, 11 − подкожная клетчатка, 12 − подкожная мышца. Микрофото. Окраска гематоксилин-эозином. Масштаб 100 мкм.
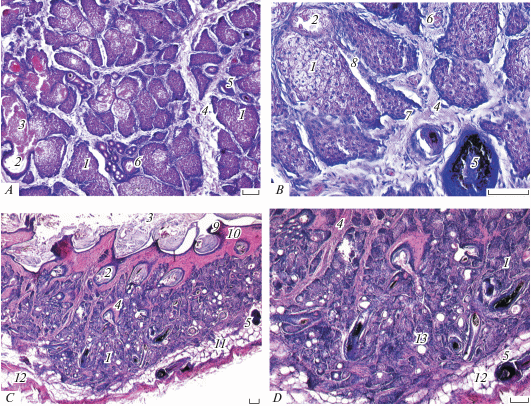
Железистая паренхима, помимо гипертрофированных сальных желез, включает систему крупных ветвящихся протоков и резервуаров секрета, а также мощную, обильно васкуляризованную, соединительнотканную строму (толщина прослоек достигает 107 мкм), которая, помимо кровеносных капилляров (рис. 4A, 46), содержит редкие гладкие мышцы волос. Площадь ацинусов на трансверсальных срезах БЖ варьируется от 2805 до 25 253 мкм2. Крупные протоки (до 226 мкм) собирают секрет от грозди крупных ацинусов железы в расширенные толстостенные резервуары с ороговевшими стенками, открывающимися на поверхности кожи широкими порами (рис. 4B, 2). Кроме того, железистая паренхима пронизана редкими некрупными фолликулами волос (рис. 4B, 5), в которые также входят более узкие протоки долей БЖ.
Секреторный эпителий в основном типичный голокриновый, ацидофильный, демонстрирует все стадии интенсивного процесса дегенерации ядер и цитоплазмы себоцитов и превращения их обломков в слоистый эозинофильный секрет. Этот секрет заполняет расширенные проксимальные отделы ацинусов и протоки. Он содержит фрагменты волос, подвергшихся лизису, при котором высвобождается незначительное количество пигмента.
Кроме голокриновых желез нами впервые у этого вида обнаружены некрупные мерокриновые гепатоидные железы (рис. 4A, 4B). Они более мелкие, часто ‒ однодольчатые, с непрозрачной темноокрашенной цитоплазмой, без признаков голокриновой секреции, но с различимыми межклеточными канальцами (рис. 4B). Альвеолы этих желез имеют форму пирамиды, расширяясь в проксимальной части, а узкие (11‒17 мкм) протоки открываются в протоки сальных желез (рис. 4B, 8).
Функциональное состояние сильно варьируется у обоих изученных самцов. У самца № 1 сальные альвеолы крупные, интенсивно функционирующие (рис. 4A, 4B), а у самца № 2 БЖ находятся в дистрофическом состоянии, полностью опорожнены, железистая паренхима почти не содержит крупных активно секретирующих альвеол, а состоит из массы мелких долек и многочисленных очень мелких цист (рис. 4C, 4D).
Среднебрюшная железа (glandula abdominalis) располагается по осевой линии в срединной части брюха впереди гениталий. Она крупнее у самцов, чем у самок и у взрослых хомяков по сравнению с молодыми (табл. 1, 3). Снаружи она заметна, благодаря полоске голой кожи и небольшому щелевидному подковообразному углублению длиной до 10 мм, имеющему хорошо выраженные края-валики толщиной 3–5 мм. На этих валиках растут пучки жестких, черных специализированных волосков, которые располагаются по спирали в боковых и донной части центрального углубления и на внутренних стенках мешка (рис. 1E, 1F). Кожные складки, лишенные волос, охватывают переднюю часть железы и сужаются в краниальном направлении. СБЖ представляет собой продолговатый мешок, залегающий под кожей. В центре этого мешка лежит центральная, выводящая секрет, тонкостенная полость, стенки которой, вероятно, способны сокращаться (рис. 1Е, 1F; рис. 5). Полость мешка может быть заполнена густым слоистым жировым секретом с фрагментами волос, подвергшихся лизису. Секрет густо покрывает темные волоски, растущие на внутренних стенках полости (рис. 1Е, 1F) и в глубоких отделах мешка. Последние особенно многочисленны у самца, у которого они заполняют всю полость железы: волосы в числе 70–97 расположены по спирали и направлены к центру железы, их стержни спутаны. Они жесткие, обильно покрытые частичками секрета, имеют короткое тонкое бесцветное основание, которое выше по стержню утолщается и уплощается, становится темным вплоть до согнутого под углом острого нерасщепленного кончика (рис. 1F; 3Е). Длина волос варьируется от 1.1 до 3.0 мм, толщина – от 23 до 34 мкм. Сердцевина волос занимает 2/3 толщины стержня. Ее строение не отличается от такового у обычных остевых волос с других топографических участков.
Орнамент кутикулы по стержню сходен с таковым обычных волос, за исключением шиловидной вершины стержня, на которой чешуйки оттопыриваются, образуя неглубокие пазухи, по-видимому, заполненные секретом (рис. 3F, 3G). Причем в области выше основания стержень искривлен, а кутикула волоса частично деформированная “оплывшая” (рис. 3F), что предполагает ее жировой лизис секретом железы, c последующим выпадением волос и их включением в состав секрета. Индекс кутикулы составляет 1.6 (11.3 ± 0.9 мкм, σ = 1.0) в основании стержня, 1.4 (12.7 ± 0.3 мкм, σ = 0.6) в деформированной области выше основания, 1.7 (37 ± 3 мкм, σ = 5.3) еще выше по стержню и 2.1 (20.3 ± 3.2 мкм, σ = 5.5) в середине стержня. Абсолютная высота чешуек изменяется значительно: по стержню – 11 мкм в основании, 37 мкм – выше основания и 20 мкм в середине стержня.
Анатомически CБЖ представляет собой настоящий мешотчатый многослойный железистый орган, толщина стенок которого достигает 2.3 мм (рис. 5A, 5B). У самца № 1 стенки-валики мешка покрыты тонким (17–23 мкм) непигментированным эпидермисом (лишенным зернистого и блестящего слоев) с небольшим (11–23 мкм), интенсивно десквамирующим роговым слоем. Характерно присутствие локальных мелких утолщений и редких коротких сосочков. Первичные и вторичные волосяные луковицы залегают поодиночке в нижних отделах сосочкового слоя, но выходят пучком из общей волосяной воронки (тип так называемых “ложных пучков”). Среди волосяных фолликулов наиболее заметны крупные изогнутые, деформированные фолликулы, содержащие сильно пигментированные направляющие волосы (рис. 4B, 5). Поверхностные отделы сосочкового слоя (толщиной 175 мкм) образованы вытянутыми параллельно поверхности кожи тонкими слабо извитыми пучками коллагеновых волокон. Гладкомышечные волокна, закрепляющиеся на волосяных фолликулах, тянутся в дорсальном направлении перпендикулярно толстому слою подкожной мускулатуры. Всю срединную часть кожи занимают сильно развитые многодольчатые голокриновые железы, толщина образуемого ими железистого слоя достигает 1.4 мм, или 60.8% толщины всей кожи. Железистый слой подстилается тонким (79 мкм) слоем подкожной клетчатки, образованной “нежной” вязью из коллагеновых волокон и содержащей лишь отдельные небольшие группы мелких (11 × 23–34 мкм) жировых клеток. Под этим слоем располагается толстый (до 2.1 мм) пласт подкожной поперечнополосатой мускулатуры, пучки которой в виде параллельных тяжей оплетают донную часть мешка (рис. 5A, 12; 5B, 12), но отдельные волокна проникают в кожные валики, окружающие полость железы. Мышечный слой подстилается рыхлой соединительнотканной прослойкой (315 мкм), не содержащей жировых клеток (рис. 5A, 14), ниже которой залегают разрозненные рыхлые конгломераты пигментных гранул (рис. 5, 15).
Рис. 5.
Микроструктура среднебрюшной железы взрослых самцов № 1 (А, B) и № 2 (C, D, E) обыкновенного хомяка (Cricetus cricetus) на сагиттальных срезах. 1–12 – как на рис. 4, 13 − центральная полость мешка, 14 − подкожная безжировая клетчатка, 15 − пигмент. D − протоки гепатоидных желез указаны стрелками. E − межклеточные канальцы гепатоидной железы указаны стрелкой. Микрофото. Окраска гематоксилин-эозином. Масштаб, мкм: A, C – 200; B – 100; D – 10.
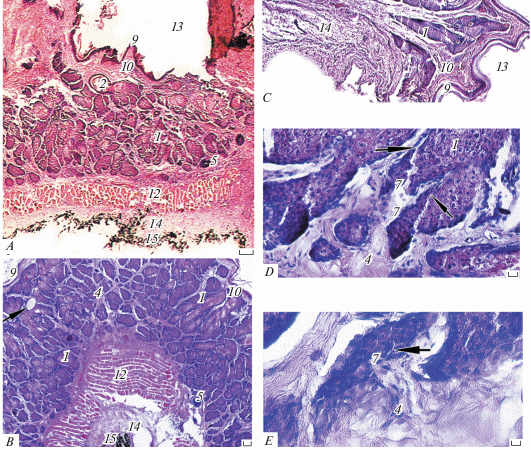
Гистология эозинофильной железистой паренхимы СБЖ в деталях отличается от таковой БЖ, и, хотя СБЖ, так же как и БЖ, многодольчатая голокриновая, в ней системы ветвящихся протоков и депонирования секрета не развиты (рис. 5A, 5B). В поверхностных отделах железистой паренхимы имеются расширенные резервуары (диаметром до 113 мкм), которые открываются вместе с волосяным пучком на глубине до 339 мкм. Размеры альвеол крупнее, чем в БЖ. Их площадь варьируется от 2541 до 67 281 мкм2. У самца № 1 секреторные альвеолы полностью заполнены себоцитами на разных стадиях дегенерации (интенсивная вакуолизация цитоплазмы и пикноз ядер). Причем вакуолизация настольно интенсивна, что опережает дегенерацию ядер, а липидные гранулы очень малы и многочисленны, поэтому цитоплазма выглядит “кружевной”. Большинство альвеол полностью заполнены обломками клеток и не имеют широких апикальных полостей, которые открывались бы в протоки, выходящие в волосяные луковицы, т.е. лизис компонентов желез не интенсивен, а выведение секрета затруднено, что компенсируется развитием мощного мышечного сфинктера. У самца № 2 СБЖ находится в состоянии инволюции (рис. 5С). Присутствие узких междольковых протоков, выводящих секрет из глубоких пирамидальных темноокрашенных альвеол и впадающих в сальные доли (рис. 5D, 5E, стрелки), а также наличие межклеточных канальцев (рис. 5E, стрелка) свидетельствуют о мерокриновой секреции этих гепатоидных желез и, в целом, голо-мерокриновой секреции всего комплекса. Это подтверждает присутствие единичных цист (рис. 5B, стрелка). Пигмент содержится лишь в луковицах крупных волос, а при их разрушении попадает в незначительном количестве в секрет.
ОБСУЖДЕНИЕ
Как отмечалось ранее, для палеарктических хомяков C. cricetus и представителей рода Mesocricetus характерен предупреждающий яркий окрас, тогда как маскирующая окраска типична для хомячков родов Allocricetulus и мелких хомячков Phodopus и Cricetulus (Воронцов, 1982; Чернова и др., 2021). Нами также подтверждены ранее опубликованные данные о том, что окрас шерсти обоих этих типов образуется за счет разнообразия и сочетания окраса и длины волос разных категорий и ярусов шерсти (Чернова и др., 2021).
Волосяной покров C. сricetus гораздо более разнороден, чем у представителей родов Phodopus (Феоктистова, 2008), Criсetulus и Allocricetulus (Чернова и др., 2021). Длинная шерсть обыкновенного хомяка состоит из нескольких отчетливо выраженных ярусов и волос нескольких категорий и порядков и отличается присутствием крупных и прямых направляющих и изогнутых остевых волос, надежно защищающих нижележащие ярусы от механических повреждений. Это подтверждается более гладкой неповрежденной кутикулой остевых и пуховых волос по сравнению с грубой и изломанной кутикулой направляющих волос. Более того, свойлачивание пуховых волос на дорсальной и боковой сторонах тела обыкновенного хомяка обеспечивает эффективную теплозащиту.
Степень развития и строение сердцевины (однорядная, двурядная лестничная, сильно пигментированная) в целом сходны у всех представителей Cricetinae, однако в крупных направляющих волосах C. сricetus сердцевина может быть сетчатой за счет увеличения поперечных рядов (до семи) сердцевинных “перегородок-дисков”.
Орнамент кутикулы сходен в основании остевых и пуховых волос с таковым у представителей Phodopus (Феоктистова 2008), но выше по стержню он у C. cricetus более сложный и грубый (рис. 2E). Орнамент кутикулы волос C. cricetus видоспецифичен, и его трудно отнести к какому-либо известному типу (Соколов и др., 1988; Чернова, Целикова, 2004).
В целом можно отметить, что структура шерстного покрова обыкновенного хомяка, наряду с другими особенностями биологии, позволяет ему существовать в широком спектре климатических условий и, соответственно, иметь огромный ареал, в отличие от большинства палеарктических хомяков.
Наше исследование в целом подтвердило ранее опубликованные данные о морфологии, гистологии, возрастной и половой изменчивости БЖ (Vrtiš, 1930, 1930a). Нами отмечено несоответствие между длиной железы у изученных нами особей (26.7 ± 0.7 мм, n = 33) и размерами БЖ, указанными в литературе – 40 × 8–10 мм, что соотносится с длиной тела, как 1 : 6 (Vrtiš, 1930а). У изученных нами особей разного возраста это соотношение изменяется у молодых особей от 1 : 11–13 до 1 : 10–12 – у взрослых. Возможно, эти различия связаны с тем, что исследованные нами и Вртишем особи относятся к разным филогеографическим линиям (Neumann et al., 2005; Feoktistova et al., 2017). Для сравнения, у взрослых самцов M. auratus это соотношение составляет 1 : 20, у взрослых самок – 1 : 11 (Vrtiš, 1930а), а, например, у водяной полевки (A. terrestris Lannaeus 1758) это соотношение у взрослых самцов – 1 : 10 (Vrtiš, 1930а) или 1 : 15–20 (Водяная полевка, 2007). У юго-западной водяной полевки (Arvicola sapidus Miller 1908) длина БЖ изменяется от 9.9 мм в декабре (n = 11) до 13.1 (n = 15) в мае (Rigaux et al., 2008). Таким образом, наблюдается высокая степень вариабельности в размерах железы у разных видов, полов, в разные сезоны года и в зависимости от условий обитания.
Наружная поверхность железистого поля БЖ не имеет покровной темной корки из затвердевшего слоя секрета, как это показано для аналогичных (или гомологичных) желез Arvicola terrestris, хотя и покрыта секретом, особенно обильным в жаркий период (Vrtiš, 1930а).
Интересно, что у обыкновенного хомяка и водяной полевки сходно устроена и мышечная система вокруг БЖ: мощный слой поперечнополосатой подкожной мускулатуры подстилает железистый пласт снизу, но у хомяка, в отличие от полевки, отдельные волокна проникают вверх в соединительнотканную строму вкупе с гладкомышечными волокнами волос (musculus arectores pilorum), что способствует выдавливанию секрета БЖ наружу (Vrtiš, 1930а).
По нашим данным, БЖ Cricetus, Criсetulus sokolovi и Allocricetulus не пигментирована. У С. griseus пигментированы только луковицы волос, расположенные в дермальном слое (Чернова и др., 2021). Это принципиально отличает БЖ вышеперечисленных видов от интенсивно пигментированных БЖ M. auratus (Lipkow, 1954). Таким образом, выделяемый секрет, который может служить пахучей меткой у M. auratus и у C. griseus в отличие от такового у других представителей Cricetinae, имеет темный цвет. Утверждение о том, что у C. cricetus БЖ сильно пигментирована и пигмент располагается в секреторных альвеолах и соединительнотканной строме (Reznik et al., 1978, 1979) нами не подтверждено.
Железистая паренхима C. cricetus в основной массе типичная голокриновая с разной степенью интенсивности секреторного процесса, которая зависит от пола, возраста особи и сезона года. Интересно отметить, что ранее авторы, как и мы, отмечали, что у М. auratus, помимо типичных голокриновых желез, действительно имеются “свободные” округлые альвеолярные дольчатые железы. Эти железы не связаны с волосяными фолликулами, лежащими в донной части и боковых частях железистого поля, и не имеют выводных протоков, а также выраженной типичной голокриновой секреции (Lipkow, 1954). Они интенсивнее окрашиваются и богаты непрозрачной цитоплазмой. По нашим данным, такие железы все же имеют редкие узкие выводные протоки, по-видимому, входящие в выводные протоки сальных желез, а также межклеточные канальцы, что служит признаком гепатоидных желез (Schaffer, 1940; Шабадаш, Чернова, 2006). У C. cricetus, возможно, эти железы относятся к типу гепатоидных желез, белковый секрет которых выделяется через клеточную мембрану в межклеточные канальцы и цисты, что, однако, требует цитологического и гистохимического подтверждения. Утверждается, что у М. auratus, по мере жирового перерождения голокриновых желез из поверхностного слоя в секреторную массу, эти эмбриональные железы продвигаются к поверхности кожи и перерождаются в сальные, замещая утраченные (Lipkow, 1954). С этим утверждением нельзя согласиться, поскольку в настоящее время известно, что в выделившую секрет сальную железу поступают новые стволовые клетки из определенной зоны “bulge” волосяного фолликула, которые под воздействием молекулярных сигналов начинают дифференцироваться в адипоциты и восстанавливают голокриновую секрецию железы (Терских и др., 2007). Гепатоидные железы являются самостоятельным компонентом голо-мерокриновых специфических желез (Шабадаш, Чернова, 2006). У многих видов млекопитающих гепатоидные железы часто содержат обильные пигментные включения, что характерно и для БЖ M. auratus (Lipkow, 1954), поэтому, вероятно, в железистой сальной паренхиме этого вида также присутствуют гепатоидные железы.
БЖ, безусловно, выделяют пахучий секрет, причем наиболее интенсивно во время спаривания, а также при мечении территории. Их расположение по бокам тела может быть связано, как и у других норных зверьков (например, у представителей землеройковых Soricidae) (Соколов, Чернова, 2001), с передвижением в узких ходах-туннелях (Lipkow, 1954). Косвенно это подтверждается продемонстрированной нами деформацией волосков, растущих на железистом поле, и его шершавой поверхностью. Наши наблюдения за поведением обыкновенного хомяка в природе показали, что как самцы, так и самки активно метят территорию около своих нор с помощью секрета этих желез. Для этого им приходится сильно изгибаться и осуществлять специфические движения (похожие на подпрыгивания), чтобы оставить секрет БЖ на камнях или растениях вблизи норы. Кроме того, в период рецептивности самки как у самцов, так и у самок БЖ становятся гораздо более заметными за счет выделения секрета, запах которого улавливает даже человек. Нами отмечалось, что при груминге хомяки интенсивно расчесывают БЖ передними и задними лапками, разнося секрет БЖ по шерсти.
У C. cricetus линейный размер СБЖ достоверно больше у самцов, чем у самок и у взрослых особей, чем у молодых, что вполне типично для большинства специфических кожных желез и, в частности, СБЖ у представителей трибы Cricetini (Воронцов, Гуртовой, 1959; Reznik et al., 1978, 1979; Феоктистова, 2008; Чернова и др., 2001). Эта особенность может быть связана c сигнальной функцией запаха секрета этой железы (Schaffer, 1940; Соколов, Чернова, 2001). Тем не менее мы отмечали активно выделяющийся секрет СБЖ у самок и даже у зверьков, только что вышедших из норы. Это никак не подтверждает роль его запаха в половом отборе. Вероятно, секрет используется при мечении ходов, входов в норы и разных объектов.
У разных представителей Cricetinae морфология СБЖ имеет свои особенности. СБЖ может представлять собой сумчатое железистое образование без волос (представители рода Allocricetulus) или со специализированными волосами (представители родов Cricetus, Phodopus) или железистое поле со специализированными волосами (представители рода Cricetulus) (Феоктистова, 2008; Чернова и др., 2001). Наибольшего развития СБЖ достигает у самого крупного представителя подсемейства – обыкновенного хомяка, у которого строение этой железы комплексное и она хорошо приспособлена для реализации функций химической коммуникации. Например, кожа перед краниальной частью железистого мешка снабжена несколькими складками, которые, судя по утолщенному сосочковидному эпидермису и лишенной желез и волос плотной дерме, трутся о субстрат, распространяя изливающийся из мешка секрет. На постоянное механическое воздействие указывают и сосочковидный и столбчатый эпидермис. СБЖ намного меньше БЖ, ее длина составляет всего лишь 2.3–2.5% длины тела обыкновенного хомяка. Специализированные волоски “осметрихии” (термин по: Müller-Schwarze et al., 1977), располагающиеся на внутренних стенках мешка, деформированы и имеют пазухи между чешуйками кутикулы, в которых накапливается секрет. К тому же они частично подвергаются лизису и эти продукты распада волос также входят в состав секрета СБЖ.
ЗАКЛЮЧЕНИЕ
Комплексное изучение кожно-волосяного покрова обыкновенного хомяка показало, что (1) его пестрый окрас определяется разнообразием цвета волос разных категорий – от черных и белых направляющих до зонарно окрашенных остевых и однотонных или бесцветных пуховых, а также ярусным строением шерсти, в которой направляющие волосы образуют верхний защитный ярус, а изогнутые остевые прикрывают сверху тонкие остевые и пуховые волосы. Причем длина пуховых волос дорсальной и боковых сторон тела значительна, и пух свойлачивается, образуя плотную подпушь, вероятно, играющую важную роль в терморегуляции. Следствием этого может быть огромный видовой ареал обыкновенного хомяка, позволяющий существовать в широком температурном диапазоне. (2) Волосы разных категорий сгруппированы в “ложные” пучки (объединяются только в общей волосяной воронке), которые располагаются неровными рядами на теле зверька. (3) Микроструктура направляющих волос видоспецифична, а наружная кутикула многослойная и грубая, служит хорошей защитой стержню. (4) Из многочисленного набора специфических желез особое положение занимают пахучие парные БЖ и непарная СБЖ, которые присутствуют у особей обоих полов разных возрастов. Анатомически они устроены неодинаково ‒ БЖ представляют собой железистое поле, а СБЖ ‒ мешотчатый орган, но гистологически они имеют много сходных черт. Волосы на обеих железах деформированы, что предполагает постоянное их травмирование, лизис, выпадение и включение в состав секрета, а также участие в нанесении или накоплении пахучего секрета (осметрихии СБЖ). (5) Многослойность СБЖ как мешотчатой структуры вместе с соединительнотканно-мышечной стромой обеспечивают эффективное выдавливание сального секрета. (6) Железистая паренхима не пигментирована, что противоречит данным литературы (Reznik et al., 1978, 1979), однако в глубоких слоях органа имеется пигментная пленка. (7) Обе железы многодольчатые, преимущественно голокриновые, с системой ветвящихся протоков и резервуаров секрета, васкуляризованной соединительнотканной стромой и мышечными элементами. (8) В обеих железах впервые у этого вида нами обнаружены мерокриновые гепатоидные железы, которые добавляют в сальный секрет белковую составляющую, возможно, придающую запах их секрету. (9) Голо-мерокриновая секреция, соединительнотканная и мышечная строма различается в деталях у БЖ и СБЖ, что предполагает различия химического состава железистой паренхимы и производимых ею секретов. (10) Сравнительно-морфологический анализ кожи и ее производных в пределах трибы Cricetini (Phodopus, Cricetulus, Allocricetulus, Mesocricetus) и хорошо изученного представителя подсемейства Microtinae (Arvicola terrestris) показал, что C. cricetus имеет целый набор структурных особенностей волос и желез, эффективно обеспечивающих его маркировочную активность и участие секретов БЖ и СБЖ в химической коммуникации вида.
Список литературы
Бодяк Н.Д., 1994. Морфофункциональные особенности гардеровой железы некоторых видов млекопитающих. Автореф. дис. … канд. биол. наук. М.: ИЭМЭЖ РАН. 24 с.
Водяная полевка. Образ вида, 2007. Пантелеев П.А., ред. М.: Наука. 512 с.
Воронцов Н.Н., 1982. Низшие хомякообразные мировой фауны. Фауна СССР. Млекопитающие / О.А. Скарлато, ред. Т. III. Ч. I. Ленинград: Наука. Ленинградское отделение. 451 с.
Воронцов Н.Н., Гуртовой Н.Н., 1959. Строение среднебрюшной железы настоящих хомяков // Доклады Академии Наук СССР. Т. 125. № 3. С. 673–676.
Кириков С.В., 1934. К вопросу о распространении черного хомяка и количественном отношении его к обыкновенной форме // Зоологический журнал. Т. 13. № 2. С. 361–368.
Крускоп С.В., 2002. Млекопитающие Подмосковья. М.: МСЮН. 2 изд. допол. исправ. 172 с.
Соколов В.Е., 1973. Кожный покров млекопитающих. М.: Наука. 489 с.
Соколов В.Е., Бодяк Н.Д., Степанова Л.В., 1994. Липидсекретирующие кожные железы обыкновенного хомяка (Cricetus cricetus Linnaeus, 1758) // Известия Российской академии наук. Серия биологическая. № 6. С. 889–897.
Соколов В.Е., Чернова О.Ф., 2001. Кожные железы млекопитающих. М.: ГЕОС. 648 с.
Соколов В.Е., Скурат Л.Н., Степанова Л.В., Шабадаш С.А., 1988. Руководство по изучению кожного покрова млекопитающих. М.: Наука. 279 с.
Терских В.В., Васильев А.В., Воротеляк Е.А., 2007. Ниши стволовых клеток // Известия Российской Академии наук. Серия биологическая. № 3. С. 261–272.
Феоктистова Н.Ю., 2008. Хомячки рода Phodopus. Систематика, филогеография, экология, физиология, поведение, химическая коммуникация. М.: Товарищество научных изданий КМК. 414 с.
Чернова О.Ф., Целикова Т.Н., 2004. Атлас волос млекопитающих (Тонкая структура остевых волос и игл в сканирующем электронном микроскопе). М.: Товарищество научных изданий КМК. 429 с.
Чернова О.Ф., Куприянов В.П., Феоктистова Н.Ю., Суров А.В., 2021. Особенности строения кожи и ее дериватов у некоторых представителей подсемейства Cricetinae (Cricetidae, Rodentia): почему это важно знать // Зоологический журнал. Т. 100. № 11. С. 1‒17.
Шабадаш С.А., Чернова О.Ф., 2006. Гепатоидные кожные железы млекопитающих. М.: Товарищество научных изданий КМК. 217 с.
Banaszek A., Bogomolov P., Feoktistova N., La Haye M., Monecke S., Reiners T.E., Rusin M., Surov A., Weinhold U., Ziomek J., 2020. Cricetus cricetus. The IUCN Red List of Threatened Species 2020: e.T5529A111875852.
Brown J.C., Williams J.D., 1972. The rodent preputial gland // Mammal Review. V. 2. P. 105–147.
Eibl-Eibesfeldt I., 1953. Zur Ethologie des Hamsters (Cricetus cricetus L.) // Zeitschrift für Tierpsychologie. Bd. 10. S. 204–266.
Feoktistova N.Yu., Meschersky I.G., Bogomolov P.L., Sayan A.S., Poplavskaya N.S., Surov A.V., 2017. Paleogeographic structure of the Common Hamster (Cricetus cricetus L.): Late Pleistocene connections between Caucasus and Western European populations // PloS ONE. V. 12 (11). e0187527. https://doi.org/10.137/journal pone 0187527
Galatík A., Galatík J., Kral Z., Galatik A., Jr., 2011. http://www.furskin.cz/overview.php?furskin=Cricetus%20cricetus
Kourist W., 1957. Das Haarleid des Hamsters (Cricetus cricetus Linne, 1758) // Weissenschaftliche Zeitschrift der Martin-Luther-University Halle-Wittenberg. Bd. 6. S. 413–438.
Kryštufek B., Hoffmann I., Nedyalkov N., Pozdnyakov A., Vohralík V., 2020. Cricetus cricetus (Rodentia: Cricetidae) // Mammalian Species. V. 52 (988). P. 10–26.
Lebedev V.S., Bannikova A.A., Neumann K., Ushakova M.V., Ivanova N.V., Surov A.V., 2018. Molecular phylogenetics and taxonomy of dwarf hamsters Cricetulus Milne-Edwards, 1867 (Cricetidae, Rodentia): description of a new genus and reinstatement of another // Zootaxa. V. 4387 (2). P. 331–349.
Lipkow J., 1954. Über das Seitenorgan des Goldhamsters Mesocricetus auratus auratus Waterh // Zeitschrift für Morphologie und Ökologie der Tiere. Bd. 42. № 4. S. 333–372.
Märkel K., 1952. Zur Kenntnis der Seitendrüsen des Goldhamsters // Zool. Anz. Bd. 149. S. 216–225.
Müller-Schwarze D., Volkman N.J., Zemanek K.F., 1977. Osmetrichia: Specialized scent hair in black-tailed deer // J. Ultrastruct. Res. V. 59. № 3. P. 223–230.
Nechay G., 2000. Status of Hamsters, Cricetus cricetus, Cricetulus migratorius, Mesocricetus newtoni and other hamster species in Europe. Strasbourg, France Council of Europe. 73 p.
Neumann K., Machaux J.R., Maak S., Jansman H.A.H., Kayser A., 2005. European common hamsters (Cricetus cricetus) — a result of repeated range expansion and 12S rRNA genes and the nuclear vWF gene // Molecular Phylogenetics and Evolution. V. 39. P. 135–145.
Neumann K., Michaux J., Lebedev V., Yigit N., Colak E., Ivanova N., Poltoraus A., Surov A., Markov G., 2006. Molecular phylogeny of the Cricetinae subfamily based on the mitochondrial cytochrome b and 12S rRNA genes and the nuclear vWF gene // Molecular phylogenetics and evolution. Academic Press. V. 39. № 1. P. 135–148.
Reznik G., Schuller H., Mohr U., 1978. Clinical Anatomy of the European Hamster: Cricetus cricetus. Philadelphia, PA: National Cancer Institute. Accessed May 11, 2019 at https://ia801305.us.archive.org/33/items/clinicalanatomyo00rezn/clinicalanatomyo00rezn.pdf
Reznik G., Reznik-Schiller H., Mohr U., 1979. Clinical anatomy of the European hamster: Cricetus cricetus. Ed. P. Dobson. DC: US Government Printing Office. 247 p.
Rigaux P., Vaslin M., Noblet J. F., Amori G., Muñoz L.J.P., 2008. “Arvicola sapidus”. The IUCN Red List of Threatened Species: e.T2150A9290712
Ružić A., 1977. Study of the population dynamics of common hamster (Cricetus cricetus L.) in Vojvodina // Zašt Bilja. V. 28. P. 289–300.
Schaffer J., 1940. Die Hautdrüsenorgane der Säugetiere. Berlin: Urban, Schwarzenberg. 464 s.
Surov A., Banaszek A., Bogomolov P., Feoktistova N., 2016. Dramatic global decrease in the range and the reproduction rate of the European hamster Cricetus cricetus // Endangered Species Research. V. 31 (1). P. 119–145.
Vrtiš V., 1930. Glandular organ on the flanks of the hamster, Cricetus cricetus (L.) // Biologické spisy. Brünn: Vysoká Skola veterinari. T. 9. F. 13–14. P. 1–31. (In Czech)
Vrtiš V., 1930a. Über die sogenannten Seitdrüsen der Wasserratte (Arvicola) und des Hamsters (Cricetus) // Archivio zoologico italiano. V. 16. S. 790–796.
Wilson D.E., Reeder D.M., 2005. Mammal Species of the World. A Taxonomic and Geographic Reference 3rd ed. Baltimore: Johns Hopkins University Press. 2142 p. www. designessentials.com
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал