

Зоологический журнал, 2022, T. 101, № 10, стр. 1190-1197
Структура сообщества бурозубок (Sorex, Eulipotyphla) Сеймчано-Буюндинской впадины (верховья реки Колыма)
Н. Е. Докучаев a, *, С. В. Киселев a, **
a Институт биологических проблем Севера ДВО РАН
685000 Магадан, Россия
* E-mail: dokuchaev@ibpn.ru
** E-mail: kiselevmagadan@mail.ru
Поступила в редакцию 30.10.2021
После доработки 05.11.2021
Принята к публикации 22.01.2022
- EDN: BKMWXS
- DOI: 10.31857/S0044513422080049
Аннотация
В границах верхнеколымского бассейна Сеймчано-Буюндинская впадина выделяется рядом природно-климатических особенностей (наличием обширного подруслового талика, богатством растительности долинно-лесного комплекса, сравнительно высокими летними температурами). По структуре сообщества бурозубок Сеймчано-Буюндинская впадина также существенно отличается от сопредельных территорий. При явном доминировании средней бурозубки положение субдоминанта здесь стабильно занимала равнозубая бурозубка. В пределах Охотско-Колымского края столь высокая доля Sorex isodon в сообществе бурозубок более характерна для Северного Приохотья. Сеймчано-Буюндинская впадина, таким образом, выступает своего рода “оазисом” для видов, проникших в верховья Колымы с Охотского побережья.
Территорию Северо-Восточной Азии населяют 10 видов землероек-бурозубок (Андреев и др., 2006). Видовой состав и долевое участие разных видов бурозубок в сообществе при этом могут сильно различаться в зависимости от географического месторасположения, разнообразия биотопов, фазы динамики численности животных и других факторов (Докучаев, 1990, 1994). Сообщество бурозубок Сеймчано-Буюндинской впадины, расположенной в верховьях р. Колыма, до недавнего времени оставалось слабоизученным. Сведения по насекомоядным этой части колымского бассейна были ограничены данными по видовому составу бурозубок, полученными при непродолжительных (в течение одного сезона) отловах мелких млекопитающих вблизи поселков Сеймчан (Юдин и др., 1976) и Усть-Среднекан (Охотина, 1971) в 60-х годах прошлого столетия. Согласно этим данным, Сеймчано-Буюндинскую впадину населяют шесть видов бурозубок: средняя (Sorex caecutiens Laxmann 1788), равнозубая (S. isodon Turov 1924), крупнозубая (S. daphaenodon Thomas 1907), бурая (S. roboratus Hollister 1913), крошечная (S. minutissimus Zimmermann 1780) и тундровая (S. tundrensis Merriam 1900). В период с 2001 по 2010 гг. в окрестностях заброшенного поселка Верхняя Буюнда (типичном участке Сеймчано-Буюндинской впадины) проводились стационарные работы по изучению популяционной экологии мелких млекопитающих – землероек-бурозубок и мышевидных грызунов (Лазуткин и др., 2007; Чернявский и др., 2007; Ямборко, 2008; Ямборко, Киселев, 2009; Лазуткин и др., 2012; Киселев, Ямборко, 2014). Достаточно длительный период и представительная выборка позволили провести анализ изменений в структуре данного сообщества бурозубок в связи с динамикой их численности. Подобного рода исследования важны для понимания принципов организации и функционирования сообществ мелких млекопитающих, их трансформации под воздействием факторов различной природы. Землеройки-бурозубки, часто формирующие многовидовые комплексы, в этом плане представляют особый интерес. Изучением динамики их сообществ занимались многие исследователи (Шефтель, 1983; Sheftel, 1989; Докучаев, 1990; Сергеев и др., 2001; Литвинов, Пожидаева, 2008; Виноградов, 2012; Литвинов и др., 2015; Локтионова и др., 2016; Нестеренко и др., 2016; Нестеренко, Локтионова, 2017; Киселев, 2019 и др.), тем не менее, особенности организации таких систем во многом остаются не раскрыты.
МАТЕРИАЛ И МЕТОДЫ
Сеймчано-Буюндинская впадина занимает нижнее течение р. Буюнда (правого притока Колымы), а также сопредельные низменные территории колымского левобережья (рис. 1). В пределах Колымского нагорья это наиболее крупное межгорное образование, которое имеет ряд природно-климатических особенностей. Согласно схеме климатического районирования (Клюкин, 1970), ее территория входит в зону резко континентального климата хвойных лесов, характеризующегося в общем достаточным увлажнением с отдельными засушливыми периодами, умеренно теплым летом и снежной зимой. Средняя температура самого холодного зимнего месяца – января (по данным м/с Сеймчан) – составляет –39.1°С, а абсолютный минимум –62°С (Справочник …, 1966). Летние температуры в Сеймчано-Буюндинской впадине самые высокие в Магаданской обл.: среднемесячная температура выше +15°С обеспечена здесь в течение 4–5 недель (Андреев, 2013: 283). Здесь возник уникальный по масштабам, крупнейший на Северо-Востоке России сквозной талик (талые грунты среди вечномерзлых пород) шириной до 19 км (Михайлов, Банцекина, 2001). Все это обусловливает разнообразие биотопов и, как следствие, сравнительно богатую растительность долинно-лесного комплекса (Хохряков, 1989: 101–103). На средних и возвышенных уровнях островной части поймы произрастают многоярусные тополево-чозениево-ивовые леса с подлеском из черной смородины (Ribes dikuscha), шиповника (Rosa acicularis), свидины (Thelycrania alba) и густым травостоем. Приречные террасы заняты лиственничными лесами ленточного типа, в настоящее время в основном вырубленными и находящимися в стадии медленной сукцессии. Лиственница часто растет в смеси с березой. Под пологом лиственниц и берез и на обширных полянах с мощным вейниковым травостоем произрастают жимолость (Lonicera edulis), шиповник, красная смородина (Ribes triste), княженика (Rubus arcticus). Надпойменные террасы покрыты обширными монотонными редкостойными лиственничными лесами на различных стадиях послепожарной сукцессии. Напочвенный покров здесь слагается мхами, лишайниками и брусникой (Vaccinium vitis-idaea), а подлесок состоит преимущественно из кустарниковых берез (Betula sp.), кедрового стланика (Pinus pumila) и голубики (Vaccinium uliginosum) (Докучаев и др., 2006).
Рис. 1.
Карта Сеймчано-Буюндинской впадины и места отлова бурозубок: поселки Сеймчан, Усть-Среднекан, Верхняя Буюнда.
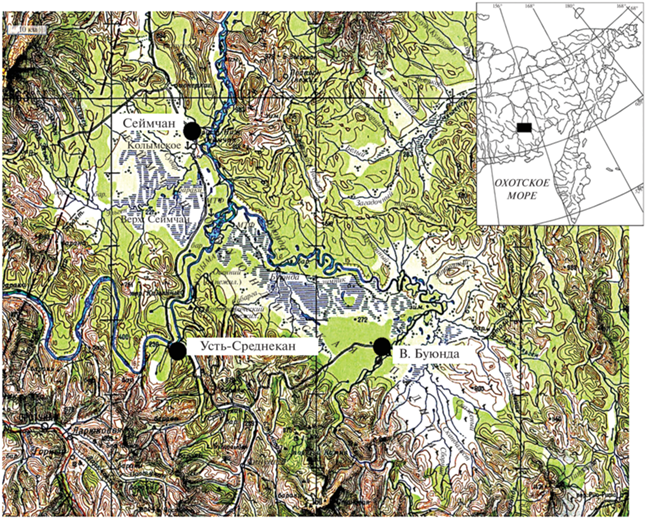
В анализе сообщества бурозубок Сеймчано-Буюндинской впадины использованы материалы, собранные в 2001–2010 гг. в окрестностях пос. Верхняя Буюнда (62°25.922′ с.ш., 153°20.476′ в.д.) (рис. 1). Животных отлавливали с использованием конусов, давилок и живоловок в трех основных биотопах: тополево-чозениево-ивовом лесу, пойменном лиственничнике и редкостойном лиственничном лесу на надпойменной террасе (Докучаев и др., 2006; Киселев, Ямборко, 2014). В конуса для быстрой гибели зверьков добавляли воду. В давилках и живоловках в качестве приманки использовали хлеб, смоченный подсолнечным маслом. Относительная численность бурозубок рассчитывалась на основе данных их отлова конусами и выражалась в количестве пойманных животных на 100 конусо-суток. Зависимость показателей относительной численности разных видов бурозубок оценивалась с использованием рангового коэффициента корреляции Спирмена. Всего на Буюндинском стационаре было отловлено 2604 экз. бурозубок пяти видов (табл. 1 ).
Таблица 1.
Виды бурозубок и их долевое участие в выборках из двух пунктов Сеймчано-Буюндинской впадины и пос. Усть-Среднекан
| Виды бурозубок |
Пункты и годы сбора материала | |||||
|---|---|---|---|---|---|---|
| пос. Усть-Среднекан, 1969 г. (Охотина, 1971) |
пос. Сеймчан, 1964 г. (Юдин и др., 1976) |
пос. Верхняя Буюнда, 2001–2010 гг. |
||||
| экз. | % | экз. | % | экз. | % | |
| S. caecutiens | 39 | 42.8 | 10 | 12.3 | 1931 | 74.1 |
| S. isodon | 28 | 30.8 | 1 | 1.2 | 406 | 15.6 |
| S. daphaenodon | 17 | 18.7 | 63 | 77.8 | 205 | 7.9 |
| S. roboratus | 3 | 3.3 | 3 | 3.7 | 46 | 1.8 |
| S. tundrensis | 2 | 2.2 | 2 | 2.5 | – | – |
| S. minutissimus | 2 | 2.2 | 2 | 2.5 | 16 | 0.6 |
| Всего | 91 | 100.0 | 81 | 100.0 | 2604 | 100.0 |
Для характеристики сообщества использовались информационные индексы Симпсона, Шеннона и Пиелу (Одум, 1986). Индекс Симпсона (Σ(ni/N)2, где n – число особей вида i; N – общее число особей) позволяет оценить степень доминирования, индекс Шеннона (–Σ ni/N ln(ni/N)) – степень разнообразия, индекс Пиелу (H/ln S, где H – индекс Шеннона, S – число видов) – равномерность (выровненность) распределения разных видов в сообществе. Статистическая обработка проведена с использованием пакета MS Excel 2010.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Из представленных в табл. 1 данных видно, что выборки бурозубок из разных участков Сеймчано-Буюндинской впадины (пос. Сеймчан и пос. Верхняя Буюнда), а также с сопредельной территории (пос. Усть-Среднекан) сильно различались. Особенно выделялась сеймчанская выборка, в которой 77.8% приходилось на крупнозубую бурозубку, а равнозубой бурозубки была отловлена всего одна особь. Существенно отличалась данная выборка и по информационным индексам (табл. 2 ). Однако собранный в течение одного сезона материал не позволяет судить, насколько типичным было в 1964 г. соотношение видов бурозубок на данном участке Сеймчано-Буюндинской впадины. В сборах из-под Усть-Среднекана три вида (средняя, равнозубая и крупнозубая бурозубки) образовали последовательно убывающей ряд. Общее долевое участие этих видов составило 92.3%. В окрестностях Буюндинского стационара во все годы доминирующее положение в сообществе занимала средняя бурозубка (рис. 2), доля которой в совокупной выборке составила 74.1% (табл. 1 ). Равнозубая бурозубка здесь занимала положение субдоминанта, и лишь однажды в этой категории ее заменила крупнозубая бурозубка. Бурая и крошечная бурозубки были отнесены в категорию второстепенных видов, поскольку их суммарная доля составила лишь 2.4%. Обращает на себя внимание и тот факт, что, несмотря на десятилетние отловы и достаточную выборку (2604 экз.), в сообществе бурозубок с Верхней Буюнды отсутствовала тундровая бурозубка, тогда как под Сеймчаном и Усть-Среднеканом этот вид был представлен в уловах, хотя и в незначительном количестве (табл. 1 ). Низкая численность S. tundrensis характерна и для других верхнеколымских районов, а в Северном Приохотье этот вид появляется только на побережье залива Шелихова (Докучаев, Поспехов, 2008).
Таблица 2.
Информационные индексы для выборок бурозубок из двух пунктов Сеймчано-Буюндинской впадины и пос. Усть-Среднекан
| Информационный индекс | Пункты и годы сбора материала | ||
|---|---|---|---|
| пос. Усть-Среднекан, 1969 г. (Охотина, 1971) |
пос. Сеймчан, 1964 г. (Юдин и др., 1976) |
пос. Верхняя Буюнда, 2001–2010 гг. |
|
| Доминирования Симпсона | 0.623 | 0.315 | 0.581 |
| Разнообразия Шеннона | 0.813 | 1.320 | 0.816 |
| Выровненности Пиелу | 0.454 | 0.737 | 0.507 |
| Число видов бурозубок | 6 | 6 | 5 |
Рис. 2.
Доля разных видов бурозубок в уловах в окрестностях пос. Верхняя Буюнда в период 2001–2010 гг.
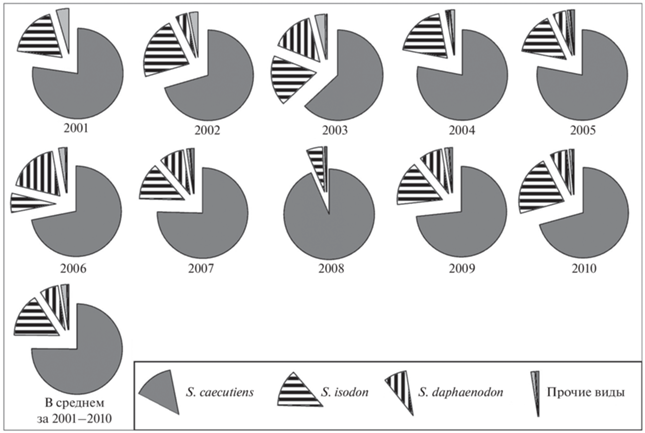
Анализ показал, что, несмотря на значительные колебания численности (рис. 3), сообщество бурозубок Буюндинского стационара характеризовалось довольно стабильной структурой. Средняя бурозубка здесь существенно преобладала в сборах во все годы исследований, составляя от 63 до 93.5%. Равнозубая бурозубка в большинстве лет занимала положение субдоминанта, лишь в 2006 г. уступив эту позицию крупнозубой бурозубке (рис. 2). В 2003 г. S. isodon и S. daphaenodon были представлены примерно в равных долях (18.0 и 15.1% соответственно). Увеличение долевого участия S. daphaenodon в сообществе в 2003 и 2006 гг. могло быть связано с фазой динамики численности S. caecutiens. Как отмечалось в ряде исследований (Ивантер, 1978; Sheftel, 1989; Докучаев, 1990; Киселев, 2019), в регуляции численности второстепенных видов бурозубок важнейшая роль отводится взаимоотношению с видом-доминантом. Динамика численности S. caecutiens в районе стационара носила циклический характер (Киселев, Ямборко, 2014). В период исследований у этого вида были сформированы три следующих друг за другом популяционных цикла (2002–2004, 2005–2007 и 2008–2010 гг.) (рис. 3). В 2003 и 2006 гг., когда представленность S. daphaenodon в сообществе была максимальной, численность S. caecutiens находилась на стадии роста после депрессий в 2002 и 2005 гг. В третьем популяционном цикле средней бурозубки (2008–2010 гг.) фаза роста численности отсутствовала. После депрессии 2008 г. численность S. caecutiens в 2009 г. сразу достигла пиковых значений (рис. 3), и существенного увеличения доли S. daphaenodon в сообществе отмечено не было (рис. 2). Бурая и крошечная бурозубки всегда были малочисленными, их суммарная доля в уловах во все годы не превышала 4.5%.
Рис. 3.
Динамика численности средней, равнозубой и крупнозубой бурозубок в бассейне р. Буюнда в 2002–2010 гг. (усреднено за июль–август по всем биотопам).
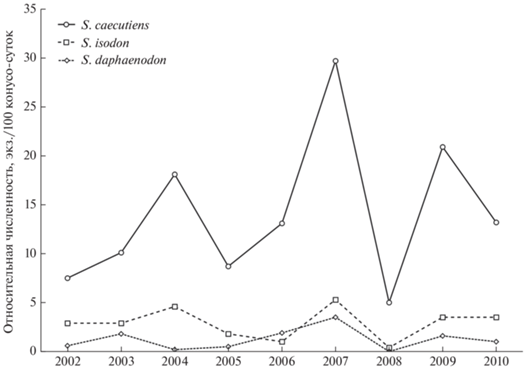
Численность разных видов бурозубок, в целом, изменялась согласованно (рис. 3). Показатели относительной численности S. isodon достоверно коррелировали с таковыми доминирующего вида – S. caecutiens (RS = 0.8, p < 0.01). Пики и минимумы численности S. daphaenodon и S. caecutiens также в большинстве лет совпадали, но связь показателей их относительной численности была недостоверна (RS = 0.6, p = 0.1). Синхронизированная динамика численности сосуществующих видов бурозубок (в особенности доминирующих) отмечалась и в других исследованиях (Sheftel, 1989; Сергеев и др., 2001; Литвинов, Пожидаева, 2008; Литвинов и др., 2015; Бобрецов, 2016; Киселев, 2019). Такая особенность может указывать на то, что данное сообщество бурозубок представляет собой хорошо связанную, стабильную во времени структуру (Литвинов, Пожидаева, 2008).
Информационные индексы в сообществе бурозубок Сеймчано-Буюндинской впадины в большинстве лет различались незначительно (табл. 3 ). Наибольшими отклонениями рассматриваемых показателей выделялись 2003 и 2008 гг. В 2003 г. индекс Симпсона имел наименьшие за весь период исследований значения, а индексы Шеннона и Пиелу – наибольшие. Было это обусловлено тем, что в этот год представленность субдоминантов и второстепенных видов была максимальной (37% в совокупности). В 2008 г. (при общей глубокой депрессии численности бурозубок) ситуация была прямо противоположной. В этот год сообщество характеризовалось низкими индексами разнообразия и выровненности, тогда как индекс Симпсона был наибольшим за все время.
Таблица 3.
Информационные индексы сообщества бурозубок Буюндинского стационара в 2001–2010 гг.
| Информационный индекс | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 |
|---|---|---|---|---|---|---|---|---|---|---|
| Доминирования Cимпсона | 0.632 | 0.545 | 0.454 | 0.645 | 0.633 | 0.557 | 0.595 | 0.878 | 0.570 | 0.551 |
| Разнообразия Шеннона | 0.65 | 0.821 | 1.021 | 0.634 | 0.723 | 0.842 | 0.802 | 0.261 | 0.835 | 0.82 |
| Выровненности Пиелу | 0.592 | 0.592 | 0.634 | 0.457 | 0.449 | 0.523 | 0.498 | 0.238 | 0.519 | 0.509 |
| Всего видов | 3 | 4 | 5 | 4 | 5 | 5 | 5 | 3 | 5 | 5 |
| Всего экз. | 22 | 193 | 338 | 405 | 218 | 260 | 460 | 154 | 356 | 198 |
Минимальным числом (по три вида бурозубок) были отмечены только 2001 и 2008 гг. (см. табл. 3 ). Если ситуацию в 2001 г. можно объяснить небольшой выборкой (всего 22 экз.), то отсутствие в уловах в 2008 г. части видов бурозубок скорее было связано с депрессией численности зверьков. Многочисленными исследованиями (Кошкина, 1966; Шефтель, 1983; Sheftel, 1989; Жигальский, Кшнясев, 2000; Жигальский, 2011) показано, что в популяциях многих видов мелких млекопитающих после лет с максимально высокой (пиковой) численностью, как правило, отмечается глубокая депрессия (популяционный “крах”). В 2007 г. численность бурозубок на Буюнде достигла максимальных за все годы значений (см. рис. 3). В 2008 г. в результате резкого сокращения их поголовья S. daphaenodon и S. minutissimus отсутствовали в уловах, а S. roboratus был представлен в единственном экземпляре.
Уже на ограниченном материале (Докучаев и др., 2006) было отмечено, что сообщество бурозубок Сеймчано-Буюндинской впадины имеет черты переходного характера между собственно колымским бассейном и Северным Приохотьем. Преобладание в сборах из-под Сеймчана в 1964 г. S. daphaenodon (табл. 1 ), сближает эту выборку с сообществом бурозубок из бассейна среднего течения р. Омолон (правого притока Колымы), где крупнозубая бурозубка в отдельные годы оказывалась в положении доминанта (Докучаев, 1990). “Колымский элемент” здесь также представляют тундровая и бурая бурозубки, отсутствующие на большей части Охотского побережья (Позвоночные …, 1996; Андреев и др., 2006; Докучаев, Поспехов, 2008). С другой стороны, высокое долевое участие в сообществе бурозубок Сеймчано-Буюндинской впадины равнозубой бурозубки придает ему значительное сходство с сообществами в приохотских районах, где этот вид весьма обычен. При этом в окрестностях стационаров “Контакт” и “Абориген”, также расположенных в верховьях Колымы (Биологические …, 1986), равнозубая бурозубка отлавливалась единичными экземплярами (Докучаев и др., 2006). В этой связи стоить отметить, что в бассейне Буюнды численность красно-серой полевки (Craseomys rufocanus (Sundevall 1846)) также существенно превышала таковую в других районах верховьев Колымы и была сопоставима с численность данного вида в Северном Приохотье (Чернявский, Лазуткин, 2004; Лазуткин и др., 2012). Переходный характер исследуемого сообщества бурозубок объясняется географическим местоположением Сеймчано-Буюндинской впадины, наличием обширного подруслового талика и сравнительно благоприятными климатическими условиями (территория защищена горами от проникновения холодных воздушных масс, как с севера, так и со стороны Охотского моря).
ЗАКЛЮЧЕНИЕ
В пределах Сеймчано-Буюндинской впадины сообщество бурозубок представлено шестью видами (средней, равнозубой, крупнозубой, бурой, крошечной и тундровой). При этом соотношение видов бурозубок на разных ее участках может сильно различаться. Так, среди 2604 экз. бурозубок, отловленных за 10-летний период в окрестностях пос. Верхняя Буюнда, S. tundrensis не была представлена, хотя ранее добывалась у пос. Сеймчан. В структуре сообщества бурозубок Сеймчано-Буюндинской впадины также выявлены существенные отличия от сообществ других территорий верхнеколымского бассейна. Здесь, при явном доминировании во все годы средней бурозубки, положение субдоминанта занимала равнозубая бурозубка. Столь высокая доля S. isodon в сообществе бурозубок Сеймчано-Буюндинской впадины сближает эту территорию с Северным Приохотьем, характеризующимся более мягкими климатическими условиями. В пределах верхнеколымского бассейна Сеймчано-Буюндинская впадина, таким образом, выступает своего рода “оазисом” для видов, проникших сюда с Охотского побережья. В целом изученное сообщество бурозубок характеризовалось относительной стабильностью, обусловленной согласованностью динамики численности составляющих его видов.
Список литературы
Андреев А.В., Докучаев Н.Е., Кречмар А.В., Чернявский Ф.Б., 2006. Наземные позвоночные Северо-Востока России: аннотированный каталог. Изд. 2-е, исправленное и дополненное. Магадан: СВНЦ ДВО РАН. 315 с.
Андреев А.В., 2013. Эталоны природы Охотско-Колымского края. Магадан: СВНЦ ДВО РАН. 322 с.
Биологические стационары советского Севера, 1986. Магадан: Магаданское книжное издательство. 78 с. (Биологические проблемы Севера).
Бобрецов А.В., 2016. Популяционная экология мелких млекопитающих равнинных и горных ландшафтов Северо-Востока европейской части России. М.: Товарищество научных изданий КМК. 381 с.
Виноградов В.В., 2012. Многолетняя динамика и структура сообщества землероек (Soricidae) горной тайги Восточного Саяна // Сибирский экологический журнал. № 1. С. 131–139.
Докучаев Н.Е., 1990. Экология бурозубок Северо-Восточной Азии. М.: Наука. 160 с.
Докучаев Н.Е., 1994. Структура и продуктивность сообществ землероек-бурозубок (Insectivora, Soricidae) Чукотки // Зоологический журнал. Т. 73. № 9. С. 114–123.
Докучаев Н.Е., Лазуткин А.Н., Ямборко А.В., Чернявский А.Ф., 2006. Бурозубки Сеймчано-Буюндинской впадины // Геология, география и биологическое разнообразие Северо-Востока России: Материалы Дальневосточной региональной конференции, посвященной памяти А.П. Васьковского и в честь его 95-летия (Магадан 28–30 ноября 2006 г.). Магадан: СВНЦ ДВО РАН. С. 338–340.
Докучаев Н.Е., Поспехов В.В., 2008. Землеройки материкового побережья залива Шелихова (Охотское море) // Сохранение биоразнообразия Камчатки и прилегающих морей: Материалы IX международной научной конференции, посвященной 100-летию с начала Камчатской экспедиции ИРГО, снаряженной на средства Ф.П. Рябушинского. (Петропавловск-Камчатский, 25–26 ноября 2008 г.). Петропавловск-Камчатский: Камчатпресс. С. 308–310.
Жигальский О.А., 2011. Структура популяционных циклов рыжей полевки (Myodes glareolus) в центре и на периферии ареала // Известия РАН. Серия биологическая. № 6. С. 733–746.
Жигальский О.А., Кшнясев И.А., 2000. Популяционные циклы европейской рыжей полевки в оптимуме ареала // Экология. № 5. С. 376–383.
Ивантер Э.В., 1978. Основные закономерности и факторы динамики численности мелких млекопитающих таежного Северо-Запада СССР // Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР. Петрозаводск: Карельский филиал АН СССР. С. 95–112.
Киселев С.В., 2019. Динамика численности и структура сообщества бурозубок в окрестностях г. Магадан (северное побережье Охотского моря) // Сибирский экологический журнал. № 5. С. 564–574.
Киселев С.В., Ямборко А.В., 2014. Динамика численности средней (Sorex caecutiens) и равнозубой (Sorex isodon) бурозубок в бассейне верхней Колымы // Зоологический журнал. Т. 93. № 9. С. 1106–1116.
Клюкин Н.К., 1970. Климат // Север Дальнего Востока. М.: Наука. С. 101–132.
Кошкина Т.В., 1966. О периодических изменениях численности полевок (на примере Кольского полуострова) // Бюллетень МОИП. Отдел биологический. Т. 71. Вып. 3. С. 14–26.
Лазуткин А.Н., Ямборко А.В., Киселев С.В., 2007. Динамика численности лесных полевок и землероек Верхней Колымы // Териофауна России и сопредельных территорий (VIII съезд Териологического общества). Материалы международного совещания (31 янв.–2 февр. 2007 г., г. Москва). М.: Товарищество научных изданий КМК. С. 249.
Лазуткин А.Н., Ямборко А.В., Киселев С.В., 2012. Популяционная динамика лесных полевок (p. Clethrionomys) верховьев Колымы (р. Буюнда) // Вестник СВНЦ ДВО РАН. № 4. С. 66–74.
Литвинов Ю.Н., Пожидаева Л.В., 2008. Анализ параметров биоразнообразия сообществ землероек гор Алтая // Сибирский экологический журнал. № 5. С. 793–798.
Литвинов Ю.Н., Дупал Т.А., Ержанов Н.Т., Абылхасанов Т.Ж., Сенотрусова М.М. и др., 2015. Особенности организации сообществ землероек открытых ландшафтов Сибири и Северного Казахстана // Сибирский экологический журнал. № 2. С. 259–267.
Локтионова Е.Ю., Нестеренко В.А., Бурковский О.А., 2016. Структура доминирования в таксоценах землероек Северного Сахалина // Вестник СВНЦ ДВО РАН. № 2. С. 93–99.
Михайлов В.М., Банцекина Т.В., 2001. Грунтово-фильтрационный талик в континентальной дельте р. Буюнда (Сеймчано-Буюндинская впадина) // Криосфера Земли. Т. 5. № 2. С. 20–28.
Нестеренко В.А., Локтионова Е.Ю., Бурковский О.А., 2016. Динамика структуры таксоцена землероек на юге о-ва Сахалин // Сибирский экологический журнал. № 3. С. 333–342.
Нестеренко В.А., Локтионова Е.Ю., 2017. Закономерности структурной динамики таксоценов землероек Сахалина // Известия РАН. Серия биологическая. № 4. С. 465–475.
Одум Ю., 1986. Экология. Т. 2. М.: Мир. 376 с.
Охотина М.В., 1971. Бурозубки (Sorex, Insectivora) Колымского нагорья // Биологические ресурсы суши севера Дальнего Востока. Т. 2. Владивосток. С. 312–314.
Позвоночные животные Северо-Востока России, 1996. Владивосток: Дальнаука. 308 с.
Сергеев В.Е., Ильяшенко В.Б., Онищенко С.С., Колегова И.А., 2001. Многолетняя динамика таксоцена бурозубок черневой тайги юга Западной Сибири // Сибирский экологический журнал. № 6. С. 785–790.
Справочник по климату СССР, 1966. Вып. 33. Магаданская область. Ч. 2. Температура воздуха и почвы. Л.: Гидрометеоиздат. 288 с.
Хохряков А.П., 1989. Анализ флоры Колымского нагорья. М.: Наука. 152 с.
Чернявский Ф.Б., Лазуткин А.Н., 2004. Циклы леммингов и полевок на Севере. Магадан: ИБПС ДВО РАН. 150 с.
Чернявский Ф.Б., Лазуткин А.Н., Ямборко А.В., 2007. Динамика и демография популяций лесных полевок (р. Clethrionomys) в таежной зоне правобережья Колымы // Вестник СВНЦ ДВО РАН. № 2. С. 107–111.
Шефтель Б.И., 1983. Зональные особенности населения насекомоядных млекопитающих енисейской тайги и лесотундры // Животный мир Енисейской тайги и лесотундры и природная зональность. М.: Наука. С. 184–203.
Юдин Б.С., Кривошеев В.Г., Беляев В.Г., 1976. Мелкие млекопитающие Севера Дальнего Востока. Новосибирск: Наука. 270 с.
Ямборко А.В., 2008. Значение трофического фактора в регуляции численности рыжих лесных полевок (р. Clethrionomys) в таежной зоне правобережья р. Колымы // Вестник СВНЦ ДВО РАН. № 1. С. 72–76.
Ямборко А.В., Киселев С.В., 2009. О синхронности флуктуаций лесных полевок (Clethrionomys, Rodentia) и землероек-бурозубок (Sorex, Insectivora) // Вестник Северо-Восточного государственного университета. Вып. 11. С. 94–95.
Sheftel B.I., 1989. Long-term and seasonal dynamics of shrews in Central Siberia // Annales Zoologici Fennici. V. 26. № 4. P. 357–369.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал