

Зоологический журнал, 2022, T. 101, № 10, стр. 1120-1126
Некоторые данные о сроках нереста и эмбриональном развитии желтобрюхой камбалы (Pleuronectes quadrituberculatus, Pleuronectidae) северной части Охотского моря
Р. Р. Юсупов a, *, Рус. Р. Юсупов a
a Институт биологических проблем Севера ДВО РАН
685000 Магадан, ул. Портовая, 18, Россия
* E-mail: ryusupov_mag@mail.ru
Поступила в редакцию 14.10.2021
После доработки 04.11.2021
Принята к публикации 09.11.2021
- EDN: CPKDKL
- DOI: 10.31857/S0044513422080128
Аннотация
Представлены результаты исследования морфометрических показателей икринок и предличинок желтобрюхой камбалы (Pleuronectes quadrituberculatus) северной части Охотского моря. Проведена инкубация икринок, собранных в природе 4 июня 2021 г. от нерестящихся особей. Диаметр икринок 1.60–1.93 мм (в среднем 1.75 мм). Результаты наших исследований в северной части Охотского моря кратко дополняют имеющиеся в литературе сведения по параметрам икры и морфологии предличинок желтобрюхой камбалы. Полная длина (TL) только что вылупившихся предличинок составляет 6.48 мм при колебаниях 5.70–7.25 мм. Общая морфология эмбрионов и предличинок совпадают с описаниями, имеющимися в литературе. Вместе с тем, у описанных нами эмбрионов и предличинок, наряду с пигментацией тела черными меланофорами, отмечена пигментация и желтыми клетками. Предличинки желтобрюхой камбалы исследуемого района отличаются от особей того же возраста других районов крупными размерами и дифференцированным пищеварительным трактом. В работе впервые представлены фото эмбрионов и предличинок в прижизненном состоянии. Анализ этапов и стадий развития икринок дает основание полагать, что в северной части Охотского моря массовый нерест желтобрюхой камбалы происходит в апреле–начале мая.
Желтобрюхая (четырехбугорчатая) камбала (Pleuronectes quadrituberculatus Pallas [1814]) – один из самых распространенных видов тихоокеанских камбал. По азиатскому побережью ареал вида охватывает, с севера на юг, акватории от бухты Провидения Чукотского моря до залива Петра Великого Японского моря. Вдоль американского побережья встречается от юго-восточной части Чукотского моря на юг до залива Аляска (Моисеев, 1953; Фадеев, 1971, 1986, 1987, 2005; Федоров и др., 2003; Mecklenburg et al., 2002; NPFMC, 2015).
Имеющиеся в литературе данные достаточно подробно освещают особенности популяционной биологии и промыслово-биологических показателей желтобрюхой камбалы по ареалу (Моисеев, 1953; Фадеев, 1965, 1971, 1987, 2005; Токранов, Заварина, 1991; Kramer et al., 1995; Борец, 1997; Датский, Андронов, 2007; Matta, 2012; Дьяков, 2014, 2015, 2017; Орлов, Токранов, 2014).
Сведения по эмбриональному развитию и морфологии только что вылупившихся предличинок этой камбалы в литературе крайне скудны. По этой теме имеются данные Перцевой-Остроумовой (1955, 1961), которые в той или иной мере заимствованы другими исследователями (Matarese et al., 1989; Григорьев, 2007). При сравнении наших материалов по желтобрюхой камбале с соответствующими литературными данными обнаружились различия в морфологии эмбрионов и предличинок.
Цель данной работы – определение сроков размножения и описание морфологии эмбрионов и вылупившихся предличинок желтобрюхой камбалы северной части Охотского моря.
МАТЕРИАЛ И МЕТОДЫ
Сбор материала провели 4 июня 2021 г. на периферии Тауйской губы у северо-западного побережья о-ва Завьялова с помощью планктонной сети Джеди на разных горизонтах от поверхности до глубины 60 м (рис. 1).
Рис. 1.
Карта-схема района проведения работ. Заштрихованной областью отмечен участок ихтиопланктонной съемки.

Отловленную икру содержали в пластиковых емкостях 500 мл с хорошо аэрируемой водой. Один раз в сутки проводили полную замену воды. Для предотвращения обрастания икринок использовали фильтрованную воду. По мере общего прогрева атмосферы среднесуточная температура воды в инкубационных емкостях постепенно повышалась с 5.6 до 7.4°C и за весь период исследований составила в среднем 6.5°C.
Развитие икры наблюдали на живом материале под микроскопом МБС-10 в вертикальной камере Черняева (1962). Параметры икринок измеряли в делениях мерной сетки окуляр-микрометра с последующим пересчетом в миллиметры. Для этой цели использовали выборку из 209 икринок, остававшихся в полости тела отнерестившейся самки с последующим их оплодотворением семенной жидкостью самца. Измерения проводили на этапе формирования плазменного бугорка. У вылупившихся предличинок измеряли полную длину – TL. Иллюстративный материал представлен по результатам микросъемки с помощью цифровой фотокамеры “Rekam Presto 40M” через микроскоп МБС-10.
РЕЗУЛЬТАТЫ
Несмотря на широкий диапазон облова, все икринки желтобрюхой камбалы были обнаружены при использовании ихтиопланктонной сети на изобатах глубже 50 м.
По результатам измерений диаметр икринок 1.60–1.93 мм, в среднем 1.75 ± 0.01 мм; диаметр желтка 1.43–1.72 мм, в среднем 1.64 ± 0.01 мм соответственно.
На момент наблюдений все эмбрионы желтобрюхой камбалы находились на этапе формирования хвостового отдела, т.е. начала его выхода за пределы сферы желтка (Махотин, 2016). Одновременно с этим процессом, вдоль дорcальной и вентральной сторон тела эмбрионов развилась тонкая полоска плавниковой каймы (рис. 2A, 2B). С развитием крупного энцефаломера произошло разделение общего нервального тяжа на головной и спинной мозг. В сформированных глазных бокалах начинают формироваться хрусталики. Слуховые капсулы хорошо развиты, но еще без отолитов. На нижней стороне головы просматривается сформировавшаяся, но еще не функционирующая сердечная трубка.
Рис. 2.
Эмбриональное развитие желтобрюхой камбалы северной части Охотского моря: A, B – образование хвостового отдела; C, D – начало пульсации сердца и двигательной активности эмбриона; E, F – только что вылупившаяся предличинка. B, D, F – вид объектов в падающем свете на темном фоне.
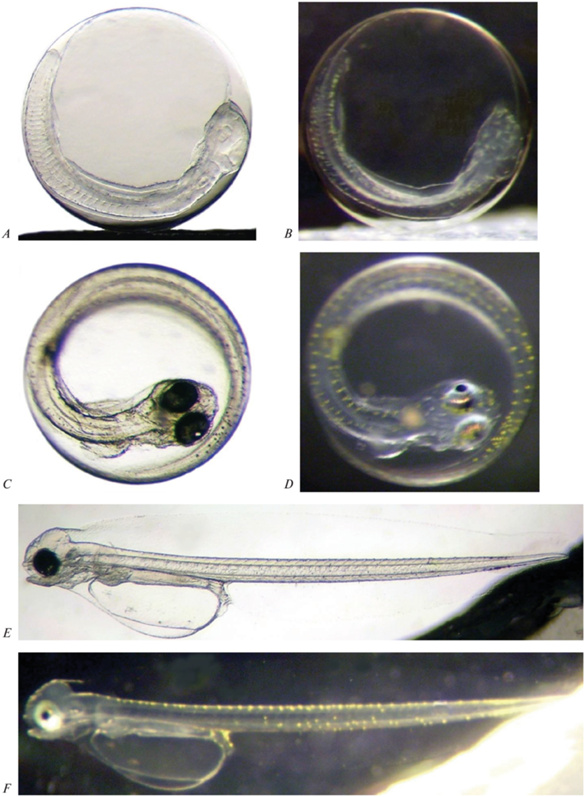
На поверхности тела эмбрионов развиваются первые пигментные клетки. В проходящем свете хорошо видны черные меланофоры, покрывающие дорcальную и, более разреженно, брюшную стороны тела. При изучении икринок в падающем свете на темном фоне обнаружено, что наряду с черными меланофорами, на поверхности тела эмбрионов развиваются многочисленные желтые и светло-желтые пигментные клетки. Наиболее густо они покрывают заднюю часть тела (включая еще несегментированный хвостовой отдел). Головной отдел пигментирован такими клетками до области зачатков грудных плавников. На поверхности каждого глазного бокала, еще не начавшего окрашиваться в темный цвет, так же развивается несколько крупных желтых пигментных клеток.
Через 6 дней инкубации (10 июня 2021 г.) у эмбрионов начинает функционировать сердце, сокращающееся с частотой 34 раз в минуту (рис. 2C, 2D). Тело эмбрионов полностью охватывает желточный мешок, конец хвостового отдела достигает заднего края грудных плавников, которые сместились вперед до уровня слуховых капсул с развитыми отолитами. Основание грудных плавников близко к вертикальному положению. Характер и интенсивность пигментации черными меланофорами остались прежними, в то же время интенсивность пигментации желтыми клетками усилилась. Глазные бокалы плотно пигментированы черными меланофорами. На поверхности радужной оболочки глаз развиваются иридоциты, придающие глазным бокалам золотистый металлический блеск.
Через 8 дней инкубации началось вылупление первых предличинок, которое завершилось через 2 сут. Размеры вылупившихся особей варьировали от 5.70 до 7.25 мм, составив в среднем 6.48 мм (рис. 2E, 2F). На фото в проходящем свете хорошо видно, что у предличинок завершается развитие не только нижней челюсти, но и верхней. Пищеварительный тракт дифференцирован на пищевод, желудок, кишечник, которые различаются структурой мышечной ткани. За исключением заднего отдела кишечника, во всех отделах пищеварительного тракта произошло расслоение внутренних стенок с образованием полости. В полости развивающегося желудка хорошо выражена складчатость слизистой оболочки. Вдоль дорсальной стороны тела по обе стороны плавниковой каймы желтые пигментные клетки образуют парный ряд. Менее упорядоченно они окрашивают вентральную сторону хвостового отдела. На участке тела между 28–32 миомерами сверху и снизу имеются небольшие продольные уплотнения пигментных рядов. Сгущение пигментации наблюдается в области ануса. По нескольку желтых пигментных клеток располагаются на поверхности нижней челюсти, области сердца, желудка и желточного мешка.
ОБСУЖДЕНИЕ
По немногочисленным литературным сведениям, нерест желтобрюхой камбалы происходит весной–в начале лета. В разных районах восточной части Берингова моря нерест наблюдался в марте–апреле (NPFMC, 2015) или в апреле–июне (Zhang et al., 1998). Мусиенко (1970) сообщает, что в Беринговом море нерест этого вида в целом длится около 3 мес., не превышая в разных районах 2–2.5 мес. В зависимости от географического положения нерестилищ и гидрологических условий сроки нереста этого вида варьируют и сдвигаются с севера на юг на более ранние сроки. Нерест начинается в ранневесенний период, в начале биологической весны, сразу же после распаления льда (с начала мая), и продолжается до середины июня (Перцева-Остроумова, 1960 цит. по: Мусиенко, 1970).
В районе Северных Курильских о-вов единичные икринки желтобрюхой камбалы были отмечены Перцевой-Остроумовой (1961) к юго-востоку от о-ва Парамушир в апреле. По мнению автора, время этих находок совпало с началом нереста. Икринки находились на I стадии развития и были недавно выметаны. В мае икра этого вида широко распространена в этом районе, как с тихоокеанской, так и с охотоморской стороны. По сообщению автора, в северной части Японского моря (южная часть Татарского пролива) начало нереста этой камбалы приходится на конец марта, а разгар нереста происходит в апреле. В районе западнокамчатского побережья Охотского моря нерестовые самки массово встречались в начале апреля (Четвергов, 2002).
Сведения о нересте четырехбугорчатой камбалы в северной части Охотского моря приводятся в работе Микулич (1959). Автор сообщает, что собранная в заливе Шелихова во второй половине мая икра находилась на завершающей стадии развития. Черешнев с соавторами (2001) указывают на более продолжительный нерест, который происходит с апреля по июнь.
Предпринятые нами в течение нескольких лет попытки обнаружить готовых к нересту производителей желтобрюхой камбалы в начале июня или позже результатов не дали. В это время все без исключения взрослые самки имели половые продукты в постнерестовом состоянии зрелости VI–II. Ихтиопланктонная съемка в начале июня 2021 г. позволила обнаружить икру. Все отловленные икринки находились на этапе отчленения хвостового отдела от желточного мешка. Поэтому можно полагать, что массовый нерест желтобрюхой камбалы в северной части Охотского моря, вероятно, происходит в апреле, а завершается воспроизводство в начале мая.
По литературным данным, размеры икринок желтобрюхой камбалы в исследованной части ареала варьируют незначительно (табл. 1). Разница между минимальными и максимальными показателями не превышают 0.54 мм. Наиболее крупные икринки этой камбалы (1.91–2.00 мм) зарегистрированы Перцевой-Остроумовой (1961) в Олюторском заливе юго-западной части Берингова моря.
Таблица 1.
Размеры икринок желтобрюхой камбалы в разных районах
| Район | Размеры, мм | Автор | |
|---|---|---|---|
| колебания | средние | ||
| Восток Берингова моря | 1.90–2.05 | – | Zhang et al., 2017 |
| Северо-восток Тихого океана | 1.67–2.21 | 1.70–1.90 | Matarese et al., 1989 |
| Район о-ва Святого Лаврентия | 1.84–1.92 | – | Перцева-Остроумова, 1961 |
| Анадырский залив | 1.73–2.02 | 1.80–1.94 | » |
| Олюторский залив | 1.76–2.21 | 1.91–2.00 | » |
| Восточное побережье Камчатки | 1.68–2.12 | 1.83–2.01 | » |
| Западное побережье Камчатки | 1.75–2.00 | 1.81–1.93 | » |
| Побережье Камчатки | 1.35–2.10 | 1.70–1.88 | Полутов, Трипольская, 1954 |
| Северная часть Охотского моря | 1.60–1.93 | 1.75 | Наши данные |
| Татарский пролив | 1.73–2.00 | 1.80–1.93 | Перцева-Остроумова, 1961 |
| Залив Петра Великого | 1.67–2.60 | – | Перцева-Остроумова, 1955 |
Несмотря на разницу в размерах икринок этой камбалы в северной части Охотского моря и на Западной Камчатке, запас питательных веществ в икринках обеих популяций примерно сходный. У особей из Охотского моря объем желтка в икринках варьирует от 1.53 до 2.66 мм3 (в среднем 2.31 ± 0.03 мм3), у особей с Западной Камчатки (рассчитан нами по данным Перцевой-Остроумовой, 1961) – 1.20–2.71 мм3. Примерно одинаковой для особей из обеих популяций оказалась и сумма тепла, необходимая для прохождения этапов и стадий от развития хвостового отдела до завершения эмбриогенеза. В нашем эксперименте эта величина составила 1248 град/ч, в опытах Перцевой-Остроумовой (1961) – 1224 град/ч.
Литературных сведений о морфологии только что вылупившихся предличинок желтобрюхой камбалы немного. Имеется описание, выполненное Перцевой-Остроумовой (1961). Автором в 1951 г. у берегов Западной Камчатки было проведено искусственное оплодотворение и инкубирование икры этого вида. Кроме того, было проанализировано большое количество икринок, предличинок и личинок, собранных на местах нереста в Татарском проливе, Беринговом и Охотском морях. Длина только что вылупившихся предличинок в опытах равнялась 5.6 (5.85) мм. Пойманные в июне 1952–1953 гг. у мыса Наварин, в Олюторском и Кроноцком заливах предличинки желтобрюхой камбалы с большими запасами желтка имели длину от 6.04 до 6.95 (5.77–6.60) мм. По сведениям Матарезе с соавторами (Matarese et al., 1989), в северо-восточной части Тихого океана вылупившиеся предличинки имели длину около 5.85 мм.
Предличинки желтобрюхой камбалы северной части Охотского моря выходят из оболочки сравнительно крупными и более развитыми, по сравнению с предличинками, использованными в опытах Перцевой-Остроумовой (1961). У только что вылупившихся в нашем эксперименте предличинок желтобрюхой камбалы исследуемого района завершается развитие не только нижней челюсти, но и верхней. Пищеварительный тракт особей этого возраста уже дифференцирован на отделы, различающиеся структурой мышечной ткани. Почти на всем протяжении кишечника произошло расслоение внутренних стенок с образованием полости. В полости желудка хорошо проявляется складчатость эпителия.
В проходящем свете характер пигментации черными меланофорами тела предличинок желтобрюхой камбалы северной части Охотского моря сходен с характером пигментации, описанным у представителей вида других районов (Перцева-Остроумова, 1961; Matarese et al., 1989; Григорьев, 2017). Отличие морфологии только что вылупившихся предличинок желтобрюхой камбалы североохотоморской популяции проявляется в развитии на теле не только черных меланофоров, но и желтых пигментных клеток.
Выявленные особенности морфологии икры и предличинок желтобрюхой камбалы исследуемого района могут рассматриваться в качестве важных диагностических признаков, позволяющих в комплексе с другими признаками надежно дифференцировать эту камбалу из общей массы уловов при проведении икорной и ихтиопланктонной съемок в северной части Охотского моря.
Список литературы
Борец Л.А., 1997. Донные ихтиоцены Российского шельфа Дальневосточных морей: состав, структура, элементы функционирования и промысловое значение // Монография. Владивосток: ТИНРО-центр. 217 с.
Григорьев С.С., 2017. Ранние стадии рыб северо-востока России (прибрежные морские воды и внутренние водоемы): атлас-определитель. Владивосток: Дальневост. отд. РАН, Тихоокеанский ин-т географии, Камч. филиал. 331 с.
Датский А.В., Андронов П.Ю., 2007. Ихтиоцен верхнего шельфа северо-западной части Берингова моря. Магадан: Северо-Восточный Научный Центр ДВО РАН. 261 с.
Дьяков Ю.П., 2014. Размерно-половая и половозрастная структура популяций дальневосточных камбал (Pleuronectidae) // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 177. С. 77–113.
Дьяков Ю.П., 2015. Половое созревание дальневосточных камбалообразных рыб (Pleuronectidae) // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого Океана. Вып. 39. С. 5–69.
Дьяков Ю.П., 2017. Общая характеристика плодовитости камбалообразных рыб северной части Тихого Океана // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 188. С. 54–88.
Махотин В.В., 2016. Эмбриональное и раннее личиночное развитие беломорской трески Gadus morhua marisalbi (Gadidae) // Вопросы ихтиологии. Т. 56. № 2. С. 177–199.
Микулич Л.В., 1959., Икринки и личинки рыб из северной части Охотского моря // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 47. С. 193–195.
Моисеев П.А., 1953. Треска и камбала дальневосточных морей // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 40. 288 с.
Мусиенко Л.Н., 1970. Размножение и развитие рыб Берингова моря // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 72. С. 166–224.
Орлов А.М., Токранов А.М., 2014. Распределение, некоторые черты биологии и динамика уловов желтоперой, четырехбугорчатой, сахалинской и колючей камбал в тихоокеанских водах северных Курильских островов и юго-восточной Камчатки // Вестник Астраханского государственного технического Университета. Серия: рыбное хозяйство. № 3. С. 29–51.
Токранов А.М., Заварина С.В., 1991. Динамика размерно-возрастной структуры желтобрюхой морской камбалы Pleuronectes quadrituberculatus Pallas на западнокамчатском шельфе // Рациональное использование биоресурсов Тихого океана. Тезисы докладов Всесоюз. конф. Владивосток: ТИНРО. С. 142–144.
Перцева-Остроумова Т.А., 1955. Определительные таблицы пелагической икры рыб для зал. Петра Великого // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 43. 135 с.
Перцева-Остроумова Т.А., 1961. Размножение и развитие дальневосточных камбал // Монография. М.: Изд-во АН СССР. 486 с.
Фадеев Н.С., 1965. Сравнительный очерк биологии камбал юго-восточной части Берингова моря и состояние их запасов // Труды ВНИРО – Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 58, 53. С. 121–138.
Фадеев Н.С., 1971. Биология и промысел тихоокеанских камбал. Владивосток: Дальиздат. 98 с.
Фадеев Н.С., 1986. Палтусы и камбалы // Биологические ресурсы Тихого океана. М.: Наука. С. 187–201.
Фадеев Н.С., 1987. Северотихоокеанские камбалы. М.: Агропромиздат. 175 с.
Фадеев Н.С., 2005. Справочник по биологии и промыслу рыб северной части Тихого океана. Владивосток: ТИНРО-Центр. 366 с.
Федоров В.В., Черешнев И.А., Назаркин А.В., Шестаков А.В., Волобуев В.В., 2003. Каталог морских и пресноводных рыб северной части Охотского моря. Владивосток: Дальнаука. 204 с.
Черешнев И.А., Волобуев В.В., Хованский И.Е., Шестаков А.В., 2001. Прибрежные рыбы северной части Охотского моря. Владивосток: Дальнаука. 197 с.
Черняев Ж.А., 1962. Вертикальная камера для наблюдения за развитием икры лососевидных рыб // Вопросы ихтиологии. Т. 2. Вып. 3. С. 457–462.
Четвергов А.В., 2002. Половое созревание западнокамчатских камбал. // Известия Тихоокеанского НИИ рыб. хоз-ва и океанографии. Т. 130. С. 940–953.
Четвергов А.В., Архандеев М.В., Ильинский Е.Н., 2000. Состав, распределение и состояние запасов донных рыб у Западной Камчатки в 2000 г. // Труды Камчатского филиала Тихоокеанского института географии ДВО РАН. Вып. IV. С. 227–257.
Matta M.E., 2012. Alaska plaice (Pleuronectes quadrituberculatus) // Resource Ecology and Fisheries Management Division, Alaska Fisheries Science Center, Seattle. 5 p.
Matarese A.C., Kendall A.W., Jr., Blood D.M., Vinter B.M., 1989. Laboratory guide to early life history stages of Northeast Pacific fishes. NOAA Tech. Rep. NMFS 80. 652 p.
Mecklenburg C.W., Mecklenburg T.A., Thorsteinson L.K., 2002. Fishes of Alaska // American Fisheries Society. Bethesda, Maryland. 1037 p.
NPFMC, 2015. Groundfish Species Profiles // NPFMC under NOAA Award. Anchorage. 60 p.
Kramer D.E. Barss W.H., Paust B.C., Bracken B.E., 1995. Guide to northeast Pacific flatfishes // Marine Advisory Bulletin. № 47. 104 p.
Zhang C., Wilderbuer T.K., Walter G.E., 1998. Biological Characteristics and Fishery Assessment of Alaska Plaice, Pleuronectes quadrituberculatus, in the Eastern Bering Sea // Marine Fisheries Review. V. 60 (4). P. 16–27.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал