

Зоологический журнал, 2022, T. 101, № 10, стр. 1096-1100
Особенности роста многощетинковых червей Spiochaetopterus typicus (Chaetopteridae)
С. Ю. Гагаев *
Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: sgagaev@zin.ru
Поступила в редакцию 10.02.2022
После доработки 14.02.2022
Принята к публикации 18.04.2022
- EDN: SWYFDD
- DOI: 10.31857/S0044513422100051
Аннотация
Изучены особенности роста популяции многощетинковых червей Spiochaetopterus typicus из Печорского моря на основе размерно-возрастной структуры популяции. С помощью линейной модификации уравнения Берталанфи установлено, что некоторые особи способны доживать до 8 лет. Возраст червей подтверждается на практике количеством годовых слоев на их трубках. Получены соотношения между диаметром тела червей, диаметром трубок и весом полихет. Предложен способ расчета биомассы полихет S. typicus с меньшими погрешностями, по сравнению с прежними методами, которые для расчета биомассы предусматривают использование исходя из диаметра трубки; в случае, если материал многочисленный, предложено вычислять биомассу с использованием среднего значения массы особи 1.5 ± 0.2 г.
Многощетинковые черви Spiochaetopterus typicus M. Sars 1856 принадлежат к одному из наиболее массовых видов полихет Северного Ледовитого океана и играют заметную роль в донных биоценозах. Длинные, тонкие черви, строящие прочные полупрозрачные трубки (которые способны долгое время после гибели червя сохраняться в грунте), в первой половине прошлого столетия являлись руководящим и характерным видом сообществ макрозообентоса почти всего Баренцева моря и некоторых районов Карского (Броцкая, Зенкевич, 1939; Филатова, Зенкевич, 1959; Денисенко, 2013). Однако повторная съемка конца 60-х Баренцева и середины 70-х Карского морей показала, что этот вид “почти повсеместно вышел из состава руководящих, а местами и характерных форм” (Семенов, 1989). Основными причинами такого изменения считают влияние тралений и возникновение методических ошибок при учете биомассы S. typicus (Жирков, 2001). Первую причину (траление) в настоящей статье оставляем без внимания, а со второй (определение биомассы) попытаемся разобраться. При работе с видом у каждого гидробиолога возникают затруднения с определением биомассы; очень часто трубки, содержащие тело червя, имеют сходный диаметр, но разную длину, так как повреждаются при отборе и промывке пробы. К примеру, взвешивая фрагменты одинакового диаметра, но разной длины 3–4 см и более 10 см, получаем разную биомассу. В настоящей работе предпринята попытка преодолеть подобную неточность и максимально приблизить значение биомассы к реальной.
Цель работы – получение способа расчета биомассы полихет S. typicus с меньшими погрешностями посредством изучения особенностей их роста.
МАТЕРИАЛ И МЕТОДИКА
Для изучения особенностей роста S. typicus были взяты 52 экз. из Печорского моря, собранные Трансарктической экспедицией 2019 г. с борта э/с “Профессор Мультановский” сотрудниками лаборатории морских исследований ЗИН РАН.
Наряду с общепринятой методикой тотальной фиксации полихет 4% раствором формалина, с дальнейшим переводом их спустя не менее 2 недель в 75% этиловый спирт, была использована методика извлечения червей из трубок после фиксации, которая была предложена ранее и которая упрощает этот процесс (Гагаев, 2010). Для этого фиксированных полихет помещали в чашку Петри с дистиллированной или пресной водой и все последующие действия проводили только в воде. Выдержав 5–10 мин, чтобы вода полностью проникла в трубки, под бинокуляром остро заточенным пинцетом вскрывали покровы трубки ближе к передней части червя по окружности. Далее пинцетом многократно и слегка сдавливали трубку, проходя по всей ее длине в любом направлении. Затем с предельной осторожностью освобождали передний отдел червя от более короткой части трубки и только после этого, слегка зажав одним пинцетом оставшуюся часть трубки, а другим – захватив червя, легкими рывками освобождали его. Нежелательно вытаскивать червя в противоположном направлении, за абдоминальный отдел; в этом случае щетинки будут препятствовать движению, и тело полихеты очень легко повредить. Таким способом (при приобретении некоторого опыта) полихеты извлекаются с большим успехом. Следует подчеркнуть, что материал предварительно должен быть хорошо зафиксирован в формалине; с мацерированным материалом желаемого результата достичь практически невозможно.
Из-за того, что трубки червей обрываются в любом месте, длина трубок не использовалась для анализа, измеряли тотально только их диаметр в самом толстом участке (обыкновенно в этом месте находится передняя часть тела червя). Отдельно было отобрано 12 целиком сохранившихся экземпляров полихет; их тела были аккуратно извлечены, измерен их диаметр в грудном отделе и определен вес с точностью до 0.001 г, а также вес наиболее полно сохранившейся трубки (размером от 5 до 10 см). На основании промеров целых и относительно неповрежденных экземпляров была определена зависимость между диаметром трубки и весом червя (с трубкой и без нее) по известной методике (Алимов, Голиков, 1974):
где Wч – вес тела червя или вес тела червя вместе с трубкой (Wтр), S – диаметр трубки в наиболее широком месте, а и b – коэффициенты.Возраст полихет аппроксимировали с помощью линейной модификации уравнения Берталанфи (Walford, 1946), опираясь на анализ размерно-возрастной структуры популяции:
где St – диаметр трубки в возрасте t, S∞ – теоретически максимальный диаметр трубки в данных условиях, k – константа роста, е – основание натуральных логарифмов. Полученные результаты были сопоставлены с количеством слоев трубки (окрашенных метиленовой синью) исходя из предположения, что они соответствуют возрасту червя.Вычисления выполнены в офисном приложении Microsoft Excel 2010 и программе PAST (Hammer et al., 2001).
РЕЗУЛЬТАТЫ
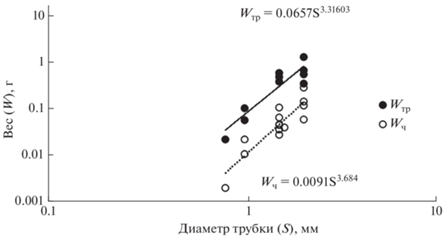
На основании проведенных измерений получена зависимость между диаметром трубки и весом тела без трубки (Wч) и с трубкой (Wтр), которые выражаются соответствующими уравнениями: Wтр = 0.0657S 3.316 и Wч = 0.0091S 3.684 (рис. 1).
Рис. 1.
Зависимость между диаметром трубки Spiochaetopterus typicus и весом тела без трубки (Wч) и с трубкой (Wтр). Масштаб логарифмический.
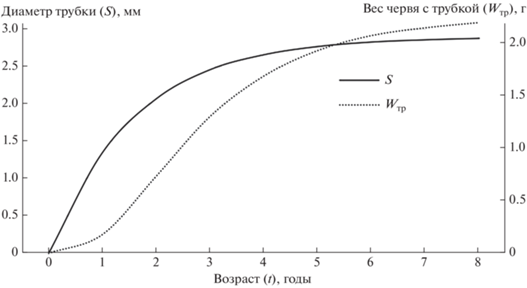
Анализ размерно-возрастной структуры популяции S. typicus позволил определить возраст размерных групп и аппроксимировать характер линейного и весового роста вида (рис. 2, 3). Проведенные расчеты показывают, что некоторые особи рассмотренной популяции могут доживать до 8 лет. Предположение подтверждается анализом поперечного среза “домиков” червей различного диаметра: число годовых слоев в трубках совпадает с расчетным возрастом S. typicus.
Рис. 2.
Размерно-возрастная структура популяции Spiochaetopterus typicus. По оси абсцисс – диаметр трубки (S), мм; по оси ординат – численность особей (N), экз.
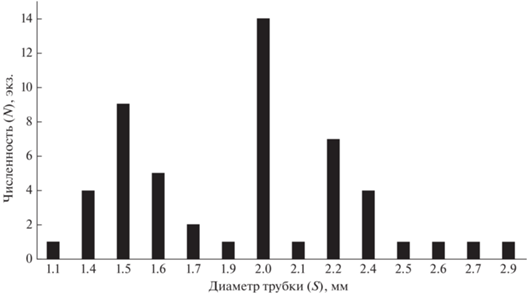
Рис. 3.
Рост Spiochaetopterus typicus. По оси абсцисс – возраст, годы; по осям ординат – диаметр трубки (S), мм и вес (W), г.
Полученные нами данные о диаметре трубки полихет и их весе приведены в табл. 1. Вычислено среднее для популяции значение веса особи с трубкой (1.5 ± 0.2 г), позволяющее с высокой точностью определять биомассу S. typicus в случае большого количества особей в выборке.
ОБСУЖДЕНИЕ
Полученная для рассматриваемого вида зависимость между весом и линейными размерами (шириной трубки и тела) свидетельствует об отрицательной аллометрии роста, т.е. о способности расти в большей степени в длину, чем в ширину или толщину. Рост тела червя и рост его трубки, измеренные в сравнительно раннем возрасте – на первом–третьем годах жизни, происходят синхронно, что в значительной степени предполагает сохранение этой зависимости вплоть до конечной стадии онтогенеза, а в случае с популяцией из Печорского моря – до 8 лет (рис. 1, 2, 3). Правильность полученной схемы роста S. typicus подтверждается совпадением числа годовых слоев в трубках животных соответствующего диаметра с их возрастом; так, при диаметре трубки 2.1 мм обнаруживаются 2 годовых слоя, соответствующих возрасту 2+, и при диаметре 2.7 мм – четыре слоя, что определяется возрастом 4+ (см. табл. 1).
Особи S. typicus становятся половозрелыми к концу первого–началу второго года жизни, в августе-сентябре, что отражено в перегибе кривой (рис. 3). Это заключение подтверждается опытом. Наличие подробных данных о полихетах других видов, обитающих в относительно сходных условиях, позволяет достаточно точно определить время их нереста (Гагаев, 2004).
Для более достоверного расчета биомассы исследуемого вида предлагается использовать данные, приведенные в табл. 1 или применить соответствующую формулу зависимости веса от ширины трубки червя. В том случае, когда численность в пробе приближается к нескольким десяткам экземпляров и измерение диаметра всех трубок потребует много времени, количество червей нужно умножить на средний вес особи, равный 1.5 г, и ни в коем случае не взвешивать трубки. Если червей немного или они явно разновозрастные, то следует измерить диаметр каждой трубки и использовать данные табл. 1 для вычисления.
Такая условность позволит провести расчет биомассы на станции с большей точностью, чем в случае, когда взвешиваются обрывки трубок самой различной длины.
S. typicus имеет планктонную личинку и расселен в Арктике на глубинах от 30 до 3440 м, при солености, близкой к океанической, в диапазоне температур от –1.8 до +11.32°С; на глубинах до 80–100 м вид сравнительно немногочислен, а глубже, в пределах шельфа, образует поселения с высокой плотностью с биомассой около 150–200 г/м2. По способу питания вид принадлежит к собирателям детрита с поверхности осадков (Фролова, 2009).
Список литературы
Алимов А.Ф., Голиков А.Н., 1974. Некоторые закономерности соотношения между размерами и весом у моллюсков // Зоологический журнал. Т. 53. № 4. С. 517–530.
Броцкая В.А., Зенкевич Л.А., 1939. Количественный учет донной фауны Баренцева моря // Труды ВНИРО. Т. 4. С. 5–126.
Гагаев С.Ю., 2004. Рост и биоэнергетические особенности популяций полихет из Чаунской губы Восточно-Сибирского моря // Океанология. Т. 44. № 2. С. 1–5.
Гагаев С.Ю., 2010. Фауна многощетинковых червей (Polychaeta) Восточно-Сибирского моря и их биогеографическая характеристика // Исследования фауны морей. Т. 63 (75). СПб.: ЗИН РАН. С. 8–40.
Денисенко С.Г., 2013. Биоразнообразие и биоресурсы макрозообентоса Баренцева моря: структура и многолетние изменения. СПб.: Наука. 284 с.
Жирков И.А., 2001. Полихеты Северного Ледовитого океана. М.: Янус-К. 632 с.
Семенов В.Н., 1989. Многолетние изменения биоценозов донной фауны Карского моря и соседних акваторий // Экология и биоресурсы Карского моря. Апатиты: Кольский научный центр АН СССР. С. 145–150.
Фролова Е.А., 2009. Фауна и экология многощетинковых червей (Polychaeta) Карского моря. Кольский научный центр РАН. Апатиты. 143 с.
Филатова З.А., Зенкевич Л.А., 1959. Количественное распределение донной фауны Карского моря // Труды ВГБО. Т. 8. С. 3–67.
Hammer Ø., Harper D.A.T., Ryan P.D., 2001. Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. № 4 (1). P. 1–9.
Walford L.F., 1946. A new graphic method of discribing the growth of animals // Biol. Bull. V. 90. № 2. P. 141–147.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал