

Зоологический журнал, 2022, T. 101, № 10, стр. 1162-1172
Распределение лесных полевок Myodes, Craseomys (Rodentia, Cricetidae, Arvicolinae) в Западной Cибири
А. А. Кислый a, b, *, Ю. С. Равкин b, c, В. П. Стариков d, С. М. Цыбулин b, В. В. Панов b, В. А. Юдкин b, e, И. Н. Богомолова b
a Омский государственный педагогический университет
644099 Омск, Россия
b Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
c Национальный исследовательский Томский государственный университет
634050 Томск, Россия
d Сургутский государственный университет
628412 Сургут, Россия
e Новосибирский государственный университет
630090 Новосибирск, Россия
* E-mail: alphaedeliways@yandex.ru
Поступила в редакцию 02.03.2022
После доработки 30.05.2022
Принята к публикации 02.06.2022
- EDN: WNOWQG
- DOI: 10.31857/S0044513422100087
Аннотация
По материалам, собранным во второй половине лета за период с 1954 по 2016 гг. в равнинной и горной частях Западной Сибири, проанализированы особенности распределения лесных полевок в зональном и провинциальном аспектах. На основании кластерного анализа матриц коэффициентов сходства показателей обилия составлены классификации местообитаний по степени благоприятности условий среды для каждого из исследованных видов. Выделено пять типов условий: оптимальные, субоптимальные, субпессимальные, пессимальные и экстремальные. При этом для красной полевки первый тип разделен на два подтипа: горный и равнинный. На основе классификаций установлена связь пространственной неоднородности обилия видов с влиянием факторов среды и их неразделимых сочетаний – природно-антропогенных режимов. Для лесных полевок характерна идентичная (кроме некоторых исключений исключениями) иерархия факторов, которые условно разделены на три группы по силе связи с распределением. Факторы первой группы – тепло- и влагообеспеченность вместе с ее широтными и высотными вариациями, второй группы – различия растительного покрова местообитаний, третьей группы – заболоченность, заливание в половодье, распашка и застроенность местообитаний, которые оказывают значительное локальное, но малое в целом по Западной Сибири влияние на распределение полевок.
Род лесных полевок Myodes Pallas 1779 в Западной Сибири и Алтае-Саянской горной стране представлен тремя видами: рыжей (Myodes glareolus Schreber 1780), красной (Myodes rutilus Pallas 1779) и красносерой (Craseomys11 rufocanus Sundevall 1846) (Павлинов, 2019).
Рыжая полевка, судя по литературным сведениям, предпочитает поляны и редколесья. Всюду избегает густых лесов с сомкнутым древостоем (Громов, Поляков, 1977; Hille, Mortelliti, 2011). На востоке ареала этой полевки больше в светлых вторичных лесах или на поросших кустарниками хвойных опушках. Обычна она в пойменных лесах (Громов, Ербаева, 1995).
Красная полевка в таежной зоне Западной Сибири и в Алтае-Саянской горной стране предпочитает леса, особенно с участием темнохвойных пород, в первую очередь кедра (Лаптев, 1958; Юдин и др., 1979; Виноградов, 2007). Кроме того, она встречена во всех местообитаниях, где имеются хотя бы отдельные участки древесной или кустарниковой растительности, обычна по окраинам моховых болот, заходит в поля, луга и поселки (Юдин и др., 1979; Лукьянова, 1980; Равкин и др., 1985; Виноградов, 2007).
Красносерая полевка обитает в лесах, часто в долинах рек, где, как правило, предпочитает густые заросли (Соколов, 1977; Christensen, Hörnfeldt, 2006). Обычна она в горно-таежных ельниках и кедрачах, по речным долинам проникает в равнинную и горную тундры (Павлинов, 2019).
Все три вида группы “лесных полевок” в Западной Сибири отнесены к лесному типу пространственного распределения как при экспертной оценке (Кислый и др., 2019), так и при интерпретации результатов кластерного анализа (Кислый и др., 2018). Тем не менее в обоих случаях можно проследить различия в предпочтении видов среди разных по степени облесенности и составу лесообразующих пород лесных и мозаичных местообитаний. Эти особенности отражены на подтиповом уровне классификаций. Цель данного исследования – количественная характеристика особенностей пространственного распределения каждого вида лесных полевок и выявление факторов среды, определяющих плотность их популяций.
МАТЕРИАЛЫ И МЕТОДИКА
Особенности распределения лесных полевок исследованы на территории Западной Сибири, в которую включена часть Западно-Сибирской равнины без правобережья Енисея, а также часть Алтай-Саянской горной страны: Кузнецко-Салаирская и Алтайская горные области (Западная Сибирь …, 1963).
Кроме личных материалов, в работе использованы сведения из литературных источников (Равкин, Лукьянова, 1976; Глотов и др., 1978; Юдин и др., 1979; Лукьянова, 1980; Вартапетов, 1982; Равкин и др., 1985, 1996, 2006; Вознийчук и др., 2006; Виноградов, 2007) и неопубликованные материалы вкладчиков банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН, собранные, как правило, с 16 июля по 31 августа за период с 1954 по 2016 гг. (Банк данных …, 2012). В общей сложности проанализированы результаты учета мелких млекопитающих в 3479 биотопах, считая места повторного сбора данных в аналогичных местообитаниях в разные годы. Помимо авторов настоящей статьи и ряда публикаций, материалы которых были использованы в данном исследовании, в сборе сведений участвовали Е.Н. Бочкарева, Е.А. Горбунова, Н.Г. Дмитриева, В.Г. Козин, А.Н. Конунова, А.В. Макаров, А.А. Одинцева, С.В. Пучковский и многие другие – всего 80 специалистов.
Учеты зверьков проведены с помощью давилок, ловчих канавок и направляющих заборчиков. Карты мест проведения учетов приведены ранее (Кислый и др., 2019b). Все показатели обилия (плотности) даны в пересчете на 100 цилиндро-суток (ц.-с.). Для этого число особей в пересчете на 100 давилко-суток сначала приведено к их количеству на 1 км2 (умножением на 400; Никифоров, 1963), а затем уменьшено в 145 раз (множитель для перевода со 100 ц.-с. на 1 км2; Равкин, Ливанов, 2008). Усредненные показатели рассчитаны без учета соотношения площадей местообитаний.
Для описания распределения лесных полевок использованы материалы, усредненные за все годы проведения учетов. Кроме того, использованы карты растительности Западной-Сибирской равнины (Ильина и др., 1985), составленные раздельно по зонам и подзонам, а также по группам выделов. Это позволяет в определенной степени минимизировать влияние межгодовых и частных (локальных) колебаний обилия исследуемого вида на результаты анализа его пространственного распределения. Из-за того, что изменения природных условий при смещении к полюсам в приполярных районах происходят гораздо интенсивнее, чем на территориях южнее северной тайги, подзона субарктических тундр при расчетах разделена на три подзональные полосы (северные моховые, низкокустарниковые и южные кустарниковые тундры), а подзона северной тайги – на две полосы (северотаежные редколесья и типичная северная тайга) в соответствии с геоботаническим районированием (Ильина и др., 1976). Для горной части исследованной территории данные усреднены по группам выделов рукописной карты В.П. Седельникова “Экосистемы республики Алтай” раздельно по провинциям с уточнением по “Ландшафтной карте Алтае-Саянского экорегиона” (2001). В указанные в тексте группы выделов входят коренные формации и их производные за исключением мелколиственных лесов, а также полностью или частично распаханных участков. Таким образом, коренные и производные мелколиственные леса рассмотрены как единое местообитание, так же как и поля (открытые, без включений древесной растительности, и в сочетании с перелесками). Поэтому, если в тексте названа лишь коренная формация, сказанное о ней относится и к большей части ее производных, кроме оговоренных.
Вид считали многочисленным в местообитаниях, где его обилие составляет 10 и более особей на 100 ц.-с., обычным – от 1 до 9, редким – от 0.1 до 0.9, очень редким – менее 0.1 (Кузякин, 1962). В случаях, когда рядом с конкретным значением не приведена единица пересчета, она соответствует 100 ц.-с.
Пространственно-типологическая организация населения и распределения отдельных видов и их групп выявлена с помощью одного из методов кластерного анализа с использованием программы “Факторная классификация” (Трофимов, 1976). В качестве меры сходства выбран коэффициент Жаккара для количественных признаков (Наумов, 1964). Алгоритм этой программы предусматривает агрегацию всех имеющихся проб в незаданное число групп таким образом, чтобы учитываемая объединением часть дисперсии матрицы сходства была максимальной. Для этого сначала из всех коэффициентов вычитают среднее по матрице значение. В результате показатели ниже этого порога становятся отрицательными. Далее строки с положительными в сумме значениями попарно объединяются, и выбирается та пара строк, объединение которых учитывает наибольшую часть дисперсии. Данные по этой паре объединяют, а значения сходства по ней усредняют. После этого процедуру поиска и агрегации повторяют. Подобное преобразование продолжается, пока учитываемая его результатами дисперсия возрастает (Трофимов, 1976; Трофимов, Равкин, 1980). При необходимости крупные классы могут быть дополнительно разделены на подклассы.
Оценка связи с факторами среды и природными режимами, как совокупностью неразделимых сочетаний факторов, проведена с помощью линейной качественной аппроксимации матриц связи (Равкин и др., 1978). Она не требует количественной оценки проявления факторов среды, достаточно их балльной оценки (например, степень облесенности) или неранжированных признаков (лесообразующие породы деревьев) (Равкин, Ливанов, 2008). Для снятия дисперсии из коэффициентов сходства проб, вошедших в одну градацию фактора или класса, вычитали среднее внутриклассовое значение сходства, а к межклассовым связям прибавляли среднее по ним значение. Это приводит к уменьшению различий в коэффициентах сходства и, соответственно, их дисперсии. Доля ее уменьшения принята в качестве учтенной части дисперсии. Объясненной ее можно считать, если анализируемый класс соответствует градациям того или иного фактора, например высокой, средней или низкой облесенности. Оценка множественной аппроксимации в этом случае рассчитана как общее уменьшение дисперсии всеми факторами и режимами, вместе взятыми. Коэффициент множественной корреляции примерно равен корню квадратному из величины доли учтенной дисперсии, представленной в долях единицы. Статистическая обработка материала проведена с использованием программного обеспечения банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН (Равкин, Ефимов, 2009).
Применение метода факторной классификации в изучении распределения одного вида вызвало необходимость некоторой корректировки методики расчета. Например, нулевое обилие полевок дает нулевые значения коэффициента сходства и образует значительное количество одиночных классов, что приводит к трудно интерпретируемым результатам. В то же время, два равных, но бесконечно малых числа, фактически мало отличающихся по предметным соображениям от нуля, сходны на 100%. Для устранения подобных искажений нулевые варианты включены в наиболее сходные по условиям среды группы выделов с отличным от нуля обилием, независимо от плотности полевок в этих выделах. Нулевые значения обилия для всех местообитаний зоны, подзоны, подзональной полосы или горной провинции, куда отдельный вид лесных полевок не проникает, заменены при расчетах на бесконечно малую величину (десятая часть наименьшего по выборке ненулевого значения). Это приводит к объединению таких проб в один класс, так же как в случае, если даже сильно различающиеся показатели обилия имеют в ряду плавный переход. Оценка информативности факторов среды и режимов проведена на основании матрицы сходства по обилию без такой замены.
После формализованного разбиения состав выделенных классов идеализирован, т.е. концептуально переработан. Для этого отклоняющиеся от разработанной концепции пробы перенесены в те группы, в которые они должны входить в соответствии с принятым объяснением. При этом, с одной стороны, оценка информативности классификации (доля учтенной ею дисперсии) снижается. С другой стороны, подобная перестановка упрощает понимание и однозначность классификации, приводя ее в соответствие с нашими представлениями о предмете исследования (Равкин, Ливанов, 2008).
РЕЗУЛЬТАТЫ
Рыжая полевка обычна как в целом по Западной Сибири, так отдельно по равнине и горным областям (по 1 особи на 100 ц.-с.22). На Западно-Сибирской равнине она распространена от подзональной полосы типичной северной тайги до лесостепи включительно, с наибольшим обилием в южной тайге (5), к северу и к югу от которой обилие уменьшается. Эта полевка встречена во всех горных провинциях Кузнецко-Салаирской и Алтайской горных областей, исключая Юго-Восточный Алтай. В горах ее обилие снижается при смещении вверх: чаще всего эту полевку встречали в предгорьях (2), реже – в низко- и среднегорьях (1 и 0.9), а высокогорные местообитания для нее наименее привлекательны (0.08). В целом в населенной рыжей полевкой части Западной Сибири этот вид предпочитает мелколиственные леса и поля-перелески. В горах благоприятна для нее черневая тайга. Эта полевка обычно избегает открытых, особенно заболоченных и заливаемых в половодье территорий, равно как и местообитаний с недостаточной теплообеспеченностью (высокогорий и, особенно, равнинных тундр) (Кислый, Одинцев, 2020).
По результатам кластерного анализа составлена классификация местообитаний по степени их благоприятности для рыжей полевки. Построены пространственно-типологический граф и картосхема ее распределения (рис. 1, табл. 1). Наиболее благоприятные условия среды, обеспечивающие максимальную плотность вида, чаще всего характерны для мелколиственных и темнохвойно-мелколиственных лесов лесной зоны равнины и ряда горных провинций. Обилие этой полевки снижается в направлении к лесам других типов и, особенно, к открытым местообитаниям.
Рис. 1.
Пространственно-типологическая неоднородность благоприятности условий среды для рыжей полевки в Западной Сибири. Граф построен на уровне типа местообитаний (порог значимости сходства 32%). Круглыми значками обозначены типы классификации. Цифры внутри – номера таксонов, соответствующие номерам в табл. 1. Рядом с индексом показано среднее сходство проб, вошедших в тип. Интенсивность заливки нижней половины значков соответствует интенсивности заливки на картосхеме. Цифры у связей между таксонами означают среднее сходство вошедших в них проб.
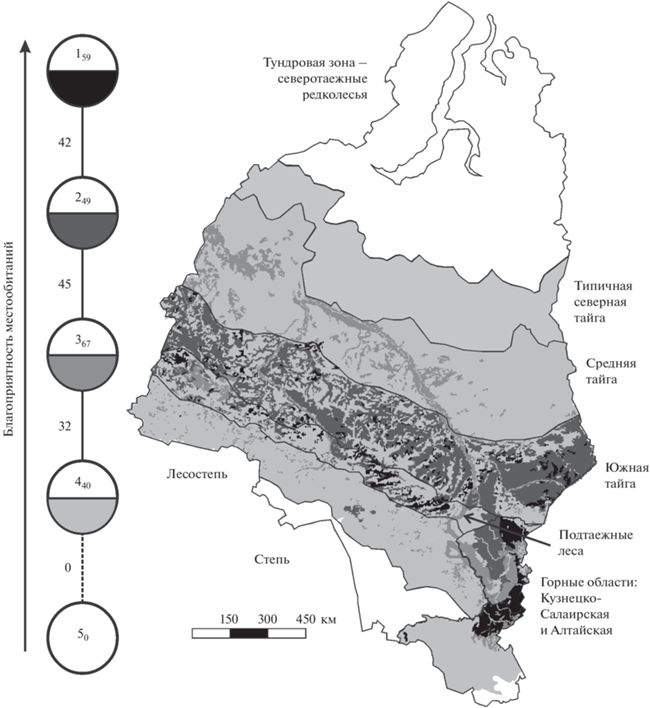
Таблица 1.
Классификация местообитаний Западной Сибири по степени их благоприятности для рыжей полевки
| Типы | Условия среды в местообитаниях | Обилие/в среднем, особей на 100 ц.-с. |
|---|---|---|
| 1 | Оптимальные в мелколиственных и темнохвойно-мелколиственных лесах южной тайги, подтаежных лесов, Северо-Предалтайской и Северо-Восточной провинций Алтая и Кузнецкого Алатау; в полях-перелесках Северо-Предалтайской провинции | 2–10/6 |
| 2 | Субоптимальные в хвойных лесах и на открытых полях южной тайги; в лиственничных лесах Северо-Восточного Алтая и Кузнецкого Алатау; в полях-перелесках равнины и Кузнецко-Салаирской горной области; в степях Кузнецкой котловины; в равнинных селитебных местообитаниях | 0.2–12/3 |
| 3 | Субпессимальные в мелколиственных лесах равнины и Кузнецко-Салаирской горной области, кроме оптимальных; в темнохвойных и сосновых лесах подтаежной подзоны, Северо-Восточного Алтая и Кузнецко-Салаирской горной области; в поймах крупных рек и долинах их притоков от средней тайги до подтаежных лесов включительно | 0.4–3/1 |
| 4 | Пессимальные в хвойных лесах и редколесьях равнины, кроме южнотаежных; в подгольцовых редколесьях; на Северо-Западном, Северном, Центральном и Восточном Алтае; в полях-перелесках Северо-Восточного Алтая; в открытых, кроме оптимальных и субоптимальных; на болотах; в поймах крупных рек и долинах их притоков в типичной северной тайге и лесостепной зоне; в сообществах пойм горных рек; в горных селитебных местообитаниях | 0.02–1/0.2 |
| 5 | Экстремальные севернее типичной северной тайги, в степной зоне и на Юго-Восточном Алтае | 0 |
Наибольшая индивидуальная связь с распределением рыжей полевки по местообитаниям Западной Сибири прослежена для тепло- и влагообеспеченности (46% учтенной дисперсии; табл. 2). Несколько меньше выражена связь с зональностью и подзональностью (33%), более чем вдвое меньше – с провинциальностью (16%). Тип растительности, высотная поясность, облесенность и состав лесообразующих пород объясняют 8–10% дисперсии. Еще слабее связано распределение вида с распашкой (1%). Менее 1% неоднородности матрицы сходства приходится на такие факторы среды, как заливание в половодье, застройка и заболоченность.
Таблица 2.
Оценка силы связи факторов среды и обилия лесных полевок в Западной Сибири
| Группа факторов, фактор, режим | Полевки | ||
|---|---|---|---|
| красносерая | рыжая | красная | |
| Группа I | |||
| Тепло- и влагообеспеченность | 28 | 46 | 10 |
| Зональность и подзональность | 25 | 33 | 10 |
| Провинциальность | 9 | 16 | 2 |
| Высотная поясность | 10 | 9 | 1 |
| Группа II | |||
| Тип растительности | 8 | 10 | 3 |
| Облесенность | 7 | 9 | 5 |
| Состав лесообразующих пород | –3 | 8 | 3 |
| Группа III | |||
| Заболоченность | 0.9 | 0.1 | 0.4 |
| Заливание в половодье | 0.6 | 0.9 | 0.7 |
| Распашка | 0.4 | 1 | 0.2 |
| Застроенность | 0.01 | 0.4 | 0.2 |
| Сочетания | |||
| Все факторы | 34 | 48 | 21 |
| Режимы классификации по благоприятности условий среды в местообитаниях | 45 | 59 | 21 |
| Все факторы и режимы | 50 | 63 | 27 |
Классификационные режимы (по благоприятности местообитаний) учитывают 59% дисперсии матрицы сходства. Оценка связи неоднородности обилия рыжей полевки со всеми выявленными факторами среды и их сочетаниями равна 63% учтенной дисперсии.
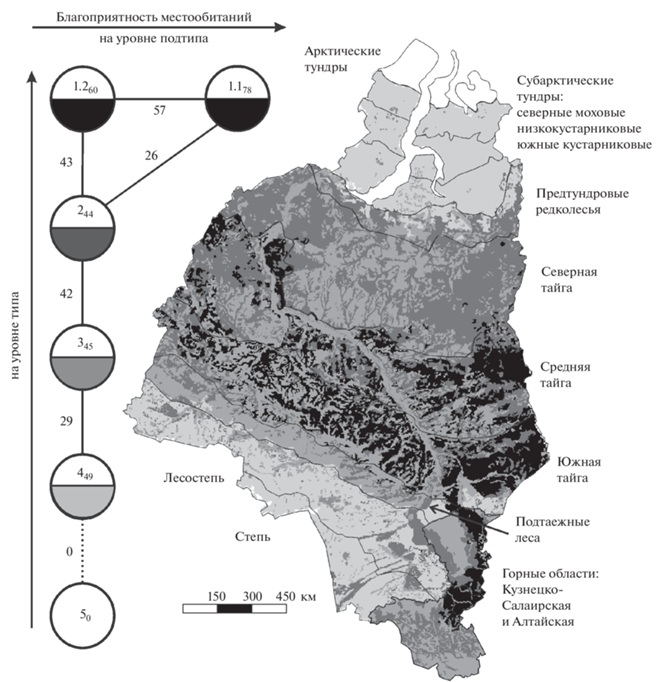
Красная полевка в части ее пространственного распределения, проанализированного по результатам кластерного анализа и аппроксимации, описана ранее (Кислый и др., 2019а). В целом по Западной Сибири наибольшее ее обилие характерно для темнохвойной (особенно горной) и черневой тайги (табл. 3). Смена лесообразующих пород и, в большей степени, уменьшение облесенности снижают благоприятность условий среды для этой полевки (табл. 2). В данной работе пространственно-типологический граф классификации местообитаний по степени благоприятности условий среды для вида дополнен картосхемой (рис. 2).
Таблица 3.
Классификация местообитаний Западной Сибири по степени их благоприятности для красной полевки
| Типы и подтипы | Условия среды | Обилие/в среднем, особей на 100 ц.-с. |
|---|---|---|
| 1 | Оптимальные в лесах с участием темнохвойных пород | 7–31/19 |
| 1.1 | Оптимальные на Северо-Восточном Алтае и Кузнецком Алатау | 19–31/24 |
| 1.2 | Оптимальные на равнинах | 7–19/14 |
| 2 | Субоптимальные (кроме оптимальных) в лесах и редколесьях | 0.4–33/6 |
| 3 | Субпессимальные (кроме пессимальных) в не лесных местообитаниях | 0.07–10/3 |
| 4 | Пессимальные в равнинных степях и тундрах | 0.04–5/0.7 |
| 5 | Экстремальные в арктических тундрах | 0 |
Рис. 2.
Пространственно-типологическая неоднородность благоприятности условий среды для красной полевки в Западной Сибири. Граф построен на уровне подтипа местообитаний (порог значимости сходства 26%). Круглыми значками обозначены типы и подтипы классификации. Цифры внутри – номера таксонов, соответствующие номерам в табл. 3. Остальные условные обозначения как на рис. 1.
Красносерая полевка в целом по Западной Сибири обычна (1), а на равнине – редка (0.9). Здесь она населяет лесную и лесостепную зоны. В среднем по зонам и подзонам больше всего этой полевки в южной тайге (3), к северу и к югу от которой обилие снижается. В целом по горной территории Западной Сибири красносерая полевка обычна (3) и встречена во всех исследованных провинциях. В горах она одинаково обычна от предгорий до среднегорий (3), а в высокогорьях ее втрое меньше. В целом по Западной Сибири красносерая полевка чаще всего населяет леса и редколесья. Различия в предпочтении насаждений с разным составом лесообразующих пород не постоянны от места к месту и, вероятно, случайны либо имеют лишь локальное значение. Несмотря на отсутствие этой полевки в равнинной тундровой зоне, ее немало в горных тундрах, особенно мохово-лишайниковых и каменистых (Кислый, 2020).
Согласно полученной классификации, наиболее благоприятны для красносерой полевки лесные участки территории южной тайги, Кузнецкого Алатау, Северо-Западного, Северного, Центрального и Северо-Восточного Алтая, включая подгольцовые редколесья. Среднее обилие по группам выделов геоботанических карт уменьшается к лесам других регионов Западной Сибири, а также к мозаичным и, особенно, открытым местообитаниям (рис. 3, табл. 4).
Рис. 3.
Пространственно-типологическая неоднородность благоприятности условий среды для красносерой полевки в Западной Сибири. Граф построен на уровне типа местообитаний (порог значимости сходства 32%). Круглыми значками обозначены типы классификации. Цифры внутри – номера таксонов, соответствующие номерам в табл. 4. Остальные условные обозначения как на рис. 1.
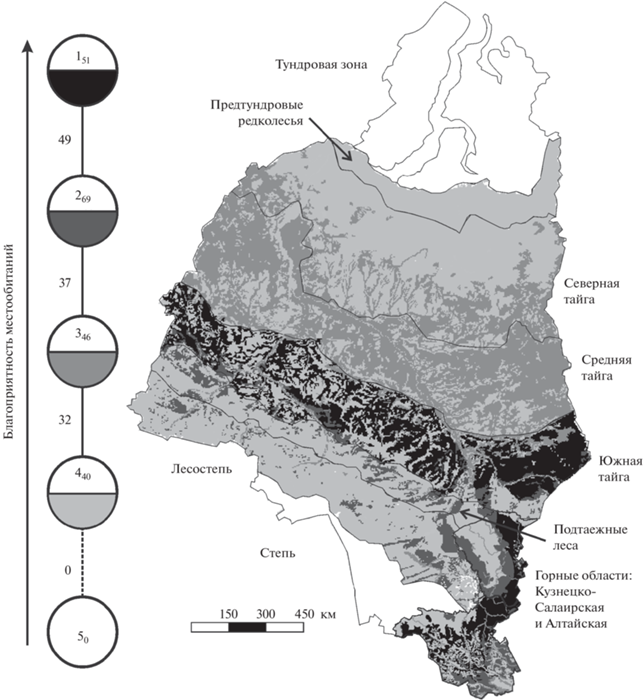
Таблица 4.
Классификация местообитаний Западной Сибири по степени их благоприятности для красносерой полевки
| Типы | Условия среды в местообитаниях | Обилие/в среднем, особей на 100 ц.-с. |
|---|---|---|
| 1 | Оптимальные в лесах и редколесьях южной тайги, Северо-Западного, Северного, Центрального и Северо-Восточного Алтая, а также Кузнецкого Алатау | 0.2–19/5 |
| 2 | Субоптимальные в лесах и редколесьях в горах, кроме оптимальных, и южнее южной тайги на равнине; в полях-перелесках южной тайги и Кузнецкого Алатау; на лугах Кузнецкого Алатау; в мохово-лишайниковых и каменистых тундрах в горах | 0.5–5/2 |
| 3 | Субпессимальные в лесах северной и средней тайги; в поймах горных и крупных равнинных рек южнее средней тайги до лесостепной зоны включительно вместе с долинами их притоков; на горных болотах; в луговых и ерниковых горных тундрах; в селитебных местообитаниях | 0.01–3/0.9 |
| 4 | Пессимальные (кроме субпессимальных) от предтундровых редколесий до средней тайги включительно; (кроме субоптимальных) на полях; на внепойменных лугах, кроме Кузнецкого Алатау; на равнинных болотах; в степных местообитаниях лесостепной зоны и тундростепях | 0.02–1/0.2 |
| 5 | Экстремальные в тундровой и степной зонах | 0 |
Наибольшая связь распределения красносерой полевки по местообитаниям Западной Сибири прослежена с тепло- и влагообеспеченностью (28% учтенной дисперсии; табл. 2). Связь с зональностью и подзональностью менее информативна (25%). Высотная поясность, провинциальность, тип растительности и облесенность объясняют от 7 до 10% дисперсии каждый. Оценка силы связи заболоченности, заливания в половодье, распашки и застройки с распределением красносерой полевки составляет менее 1%. Классификационные режимы (по благоприятности местообитаний) учитывают 45% дисперсии матрицы сходства и дают приращение в 16%. Множественная оценка связи со всеми выявленными факторами среды и их сочетаниями равна 50% учтенной дисперсии.
ОБСУЖДЕНИЕ
Выявленный по выполненным классификациям местообитаний Западной Сибири список факторов среды и их сочетаний, связанных со встречаемостью лесных полевок, можно условно разделить на три группы по силе этих связей. Список этих факторов, а также их внутригрупповая иерархия не одинаковы для каждого из исследованных видов полевок.
Первая группа включает в себя тепло- и влагообеспеченность – наиболее информативный для всех исследованных видов полевок фактор, представляющий собой сочетание зональности и подзональности на равнине, провинциальности и высотной поясности в горах.
Во вторую группу входят факторы, демонстрирующие связь распределения лесных полевок с обликом растительности: тип растительного покрова, облесенность и состав лесообразующих пород.
Третью группу факторов составляют заливание в половодье, заболоченность, распашка и застроенность. Связь их с неоднородностью обилия лесных полевок в целом по Западной Сибири невелика и чаще всего проявляется в местообитаниях вне основных биотопических преференций.
Несмотря на явное предпочтение лесными полевками облесенных местообитаний Западной Сибири, межвидовые различия в преференции существенны. Это подтверждают и результаты оценки силы связи обилия видов и природно-антропогенных факторов среды. Велика среди лесных полевок и разница в доле дисперсии матрицы сходства, учтенной всеми выявленными факторами и режимами вместе. Причиной этого служит различный характер распределения и среднего обилия полевок в целом по Западной Сибири: вслед за увеличением доли нулевых проб снижается дисперсия показателей обилия в выборке, в том числе и связанная с межгодовыми и локальными различиями в численности животных. Доли учтенной всеми выявленными факторами среды и режимами дисперсии варьируют от 27% для красной полевки, встреченной на всей исследованной территории, кроме арктических тундр, до 63% для рыжей полевки, отсутствующей севернее типичной северной тайги, в степной зоне и на Юго-Восточном Алтае (среднее по Западной Сибири обилие: 6 и 1 особь на 100 ц.-с. соответственно).
ЗАКЛЮЧЕНИЕ
Все три вида лесных полевок в целом предпочитают леса, а красносерая полевка – еще и подгольцовые редколесья. Рыжая и красная полевки избирательны в предпочтении лесных биотопов с разным породным составом деревьев: для первой большее обилие прослежено в мелколиственных и черневых лесах, а для второй – в местообитаниях с участием темнохвойных пород.
Иерархия выявленных факторов среды, сопряженных с неоднородностью обилия для каждого из видов лесных полевок, в целом сходна. Эти факторы разделены на три группы. Первую группу составляет тепло- и влагообеспеченность, включая зонально-подзональные, провинциальные и высотные варианты этого фактора. В целом по Западной Сибири сила связи этой группы с распределением наиболее высока. Вторая группа состоит из факторов, непосредственно определяющих предпочтения полевок: тип растительности, облесенность и состав лесообразующих пород. Их влияние выражено меньше, чем влияние тепло- и влагообеспеченности, однако может превосходить по силе связи отдельные (зонально-подзональные, провинциальные или высотные) вариации этого градиента среды. Третья группа – факторы со слабо выраженным в целом по Западной Сибири влиянием на распределение лесных полевок: заливание в половодье, заболоченность, распашка и застроенность.
Список литературы
Абрамсон Н.И., Лисовский А.А., 2012. Подсемейство Arvicolinae // Млекопитающие России: систематико-географический справочник. Павлинов И.Я., Лисовский А.А. (ред.). Москва: Товарищество научных изданий КМК. С. 127–141.
Банк данных: информация, правила для вкладчиков. Сайт лаборатории зоологического мониторинга ИСиЭЖ СО РАН [Электронный ресурс]. Режим доступа: http://eco.nsc.ru/zoomonit/zoomonit_r.htm (дата обновления: 05.01.2012).
Вартапетов Л.Г., 1982. Сообщества мелких млекопитающих таежных междуречий Западной Сибири // Размещение и численность позвоночных Сибири. Новосибирск: Наука. С. 237–253.
Виноградов В.В., 2007. Мелкие млекопитающие Кузнецкого Алатау. Красноярск: Изд-во Красноярского государственного педагогического университета. 212 с.
Вознийчук О.П., Богомолова И.Н., Ливанов С.Г., Вартапетов Л.Г., 2006. Классификация мелких млекопитающих Центрального Алтая по сходству распределения // Сибирский экологический журнал. Т. 13. № 4. С. 541–547. [Vozniychuk O.P., Bogomolova I.N., Livanov S.G., Vartapetov L.G., 2006. Classification of Small Mammals of the Central Altay According to the Similarity of Distribution // Contemporary Problems of Ecology. Vol. 13. № 4. P. 541–547. (in Russian)].
Глотов И.Н., Ердаков Л.Н., Кузякин В.А., Максимов А.А., Мерзлякова Е.П., Николаев А.С., Сергеев В.Е. и др., 1978. Сообщества мелких млекопитающих Барабы. Новосибирск: Наука. 232 с.
Громов И.М., Поляков И.Я., 1977. Полевки (Microtinae). Фауна СССР, млекопитающие. Л.: Наука. 504 с.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: ЗИН РАН. 522 с.
Западная Сибирь: природные условия и естественные ресурсы СССР, 1963. Отв. ред. Г.Д. Рихтер. М.: АН СССР. 487 с.
Ильина И.С., Лапшина Е.И., Лавренко Н.Н., Мельцер Л.И., Романова Е.А., Богоявленский Б.А., Махно В.Д., 1985. Растительный покров Западно-Сибирской равнины. Новосибирск: Наука. 251 с.
Ильина И.С., Лапшина Е.И., Лавренко Н.Н., Мельцер Л.И., Романова Е.А., Богоявленский Б.А., Махно В.Д., 1976. Растительность Западно-Сибирской равнины. Карта масштаба 1: 1 500 000. М.: ГУГК СССР.
Кислый А.А., 2020. Распределение красносерой полевки Craseomys rufocanus (Sundevall, 1846) на равнинных и горных территориях Западной Сибири // Симбиоз-Россия 2020: сборник статей XII Всероссийского конгресса молодых ученых-биологов с международным участием. Пермь: ПГНИУ. С. 129– 134.
Кислый А.А., Одинцев О.А., 2020. Распределение рыжей полевки Myodes glareolus (Schreber, 1780) на равнинных и горных территориях Западной Сибири // Познание и деятельность: от прошлого к настоящему: материалы II Всероссийской междисциплинарной научной конференции. Омск: ОмГПУ. С. 311–316.
Кислый А.А., Равкин Ю.С., Богомолова И.Н., Одинцева А.А., Цыбулин С.М., Одинцев О.А., 2019. Распределение и территориальная неоднородность населения мелких млекопитающих южной тайги Западной Сибири // Зоологический журнал. Т. 98. № 3. С. 343–352.
Кислый А.А., Равкин Ю.С., Богомолова И.Н., Цыбулин С.М., Стариков В.П., Панов В.В., Юдкин В.А., Вартапетов Л.Г., Соловьев С.А., 2019а. Распределение красной полевки Myodes rutilus (Pallas, 1779) в Западной Сибири // Сибирский экологический журнал. Т. 26. № 1. С. 14–28. [Kislyi A.A., Ravkin Yu.S., Bogomolova I.N., Tsybulin S.M., Starikov V.P., Panov V.V., Yudkin V.A., Vartapetov L.G., Solovev S.A., 2019. Distribution of Northern Red-Backed Vole Myodes rutilus (Pallas, 1779) in Western Siberia // Contemporary Problems of Ecology. Vol. 12. № 1. P. 10–22. DOI: 10.1134/S1995425519010086 (in Russian)].https://doi.org/10.15372/SEJ20190102
Кислый А.А., Равкин Ю.С., Стариков В.П., 2018. Распределение мелких млекопитающих в равнинных и горных ландшафтах Западной Сибири // Актуальные вопросы биогеографии. СПб.: СПбГУ. С. 186–188.
Кузякин А.П., 1962. Зоогеография СССР // Ученые записки Московского областного пединститута. Т. 109. № 1. С. 3–182.
Ландшафтная карта Алтае-Саянского экорегиона, 2001. М 1: 2 250 000. М.: ИГЕМ РАН – WWF Russia.
Лаптев И.П., 1958. Млекопитающие таежной зоны Западной Сибири. Томск: ТГУ. 285 с.
Лукьянова И.В., 1980. Количественная характеристика населения мелких млекопитающих Северо-Восточного Алтая // Проблемы зоогеографии и истории фауны. Новосибирск: Наука. С. 255–273.
Наумов Р.Л., 1964. Птицы природного очага клещевого энцефалита Красноярского края. Автореф. дис. … канд. биол. наук. М.: Московский областной пединститут. 19 с.
Никифоров Л.П., 1963. Опыт абсолютного учета численности мелких млекопитающих в лесу // Организация и методы учета птиц и вредных грызунов. М.: АН СССР. С. 237–243.
Павлинов И.Я., 2019. Звери России: справочник-определитель. Ч. 1. Насекомоядные, рукокрылые, зайцеобразные, грызуны. М.: Товарищество научных изданий КМК. 340 с.
Равкин Ю.С., Богомолова И.Н., Ердаков Л.Н., Панов В.В., Буйдалина Ф.Р., Добротворский А.К., Вартапетов Л.Г., Юдкин В.А., Торопов К.В., Лукьянова И.В., Покровская И.В., Жуков В.С., Цыбулин С.М., Фомин Б.Н., Стариков В.П., Шор Е.Л., Чернышова О.Н., Соловьёв С.А., Чубыкина Н.Л., Ануфриев В.М., Бобков Ю.В., Ивлева Н.Г., Тертицкий Г.М., 1996. Особенности распределения мелких млекопитающих Западно-Сибирской равнины // Сибирский экологический журнал. Т. 3. № 3–4. С. 307–317. [Ravkin Yu.S., Bogomolova I.N., Erdakov L.N., Panov V.V., Buidalina F.R., Dobrotvorsky A.K., Vartapetov L.G., Yudkin V.A., Toropov K.V., Lukyanova I.V., Pokrovskaya I.V., Zhukov V.S., Tsybulin S.M., Fomin B.N., Starikov V.P., Shor E.L., Chernyshova O.N., Solovev S.A., Chubykina N.L., Anufriev V.M., Bobkov Yu.V., Ivleva N.G., Tertitsky G.M., 1996. Peculiarities of distribution of small mammals of West Siberian Plain // Contemporary Problems of Ecology. Vol. 3. № 3–4. P. 307–317. (in Russian)].
Равкин Ю.С., Гуреев С.П., Покровская И.В., Фомин Б.Н., Вартапетов Л.Г. Бурский О.В., Вахрушев А.А., Преображенская Е.С., Малков Н.П., Равкин Е.С., Козлов Н.А., Торопов К.В., Блинов В.Н., Юдкин В.А., Жуков В.С., Стариков В.П., Богомолова И.Н., Родькина В.И., 1985. Пространственно-временнáя динамика животного населения. Новосибирск: Наука. 206 с.
Равкин Ю.С., Ефимов В.М., 2009. Банк данных по численности и распределению животных в пределах бывшего СССР // Формирование баз данных по биоразнообразию – опыт, проблемы, решения. Материалы Международной научно-практической конференции. Барнаул: “АRТIКА”. С. 205–214.
Равкин Ю.С., Куперштох В.Л., Трофимов В.А., 1978. Пространственная организация населения птиц // Птицы лесной зоны Приобья. Новосибирск: Наука. С. 253–269.
Равкин Ю.С., Ливанов С.Г., 2008. Факторная зоогеография. Новосибирск: Наука. 205 с.
Равкин Ю.С., Лукьянова И.В., 1976. География позвоночных южной тайги Западной Сибири. Новосибирск: Наука. 338 с.
Равкин Ю.С., Юдкин В.А., Панов В.В., Стариков В.П., Ердаков Л.Н., Вартапетов Л.Г., Богомолова И.Н., Ильяшенко В.Б., Онищенко С.С., Цыбулин С.М., Сорокина Н.В., Соловьёв С.А., Блинов В.Н., Жуков В.С., Покровская И.Н., Блинова Т.К., Торопов К.В., Сазонова И.А., Чернышова О.Н., Ануфриев В.М., Тертицкий Г.М., Москвитина Н.С., Бахина Е.В., 2006. Особенности картографирования и выявления пространственно-типологической структуры населения мелких млекопитающих (на примере Западной Сибири) // Биоразнообразие и динамика экосистем. Информационные технологии и моделирование. Интеграционные проекты. Новосибирск: Изд-во Сибирского отделения Российской академии наук. Вып. 7. С. 258–276.
Соколов В.Е., 1977. Систематика млекопитающих. Отряды: зайцеобразных, грызунов. М.: Высшая школа. 494 с.
Трофимов В.А., 1976. Модели и методы качественного факторного анализа матрицы связи // Проблемы анализа дискретной информации. Новосибирск: Наука. Ч. 2. С. 24–36.
Трофимов В.А., Равкин Ю.С., 1980. Экспресс-метод оценки связи пространственной неоднородности животного населения и факторов среды // Количественные методы в экологии животных. Ленинград: Наука. С. 113–115.
Юдин Б.С., Галкина Л.И., Потапкина А.Ф., 1979. Млекопитающие Алтае-Саянской горной страны. Новосибирск: Наука. 296 с.
Christensen P., Hörnfeldt B., 2006. Habitat Preferences of Clethrionomys rufocanus in Boreal Sweden // Landscape Ecology. V. 21. P. 185–194. https://doi.org/10.1007/s10980-005-1052-6
Hille S., Mortelliti A., 2011. Microhabitat partitioning of Apodemus flavicollis and Myodes glareolus in the sub-montane alps: a preliminary assessment // Hystrix, the Italian Journal of Mammalogy. V. 21. P. 157–163. https://doi.org/10.4404/Hystrix-21.2-4458
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал