

Зоологический журнал, 2022, T. 101, № 11, стр. 1286-1299
Ископаемая фауна мелких млекопитающих из пещеры Иманай (Южный Урал, Россия)
Т. В. Фадеева a, *, Д. О. Гимранов b, **, П. А. Косинцев b, ***, А. Г. Яковлев c, ****
a Горный институт, Пермский федеральный исследовательский центр Уральского отделения РАН
614007 Пермь, Россия
b Институт экологии растений и животных Уральского отделения РАН
620144 Екатеринбург, Россия
c Институт геологии Уфимского федерального исследовательского центра РАН
450077 Уфа, Россия
* E-mail: fadeeva.tatyana@mail.ru
** E-mail: djulfa250@rambler.ru
*** E-mail: kpa@ipae.uran.ru
**** E-mail: a_jakovlev@mail.ru
Поступила в редакцию 27.01.2022
После доработки 20.03.2022
Принята к публикации 22.03.2022
- EDN: ABOMPV
- DOI: 10.31857/S004451342210004X
Аннотация
Исследована ископаемая фауна мелких млекопитающих из отложений пещеры Иманай (53°02′ с.ш., 56°26′ в.д.) (Южный Урал, Россия). В фауне преобладают виды открытых биотопов, узкочерепная полевка (Microtus (Stenocranius) gregalis) является доминирующим видом. Выявлено два типа фауны, которые характеризуют ее состав и структуру в конце позднего плейстоцена и, предположительно, в раннем голоцене. Содоминирующим видом фауны первого типа (нижние и средние отложения пещеры) является степная пеструшка (Lagurus lagurus). Для этого типа характерны значительное преобладание доли (83.0–92.2%) степных видов и низкая доля (1.5–3.9%) лесных видов млекопитающих, что является доказательством распространения в позднеледниковье на территории южной части Уральских гор преимущественно открытых ландшафтов. В фауне второго типа (верхние отложения пещеры) содоминируют пищухи (Ochotona sp.) и относительно высока доля лесных видов (14.3–21.4%), что свидетельствует о появлении лесных формаций в конце позднеледниковья или в раннем голоцене на этой территории. Выборки первых нижнекоренных зубов узкочерепных полевок характеризуются высокой долей зубов (>50%) с простыми вариантами строения непарной петли антероконидного отдела (“грегалоидный” морфотип). В выборках зубов степных пеструшек зафиксирована высокая доля (до 51.6%) зубов “транзиенсногo” морфотипа. Среди зубов в нижней части отложений пещеры обнаружены бескорневые цементные первые нижнекоренные зубы (m1) и третьи верхнекоренные зубы (M3), строение жевательной поверхности которых (широкое слияние треугольников Т4-Т5 и Т2-Т3, соответственно) характерно для древних полевок Microtus (Stenocranius) gregaloides и M. (Terricola) arvalidens из фаун первой половины раннего плейстоцена–второй половины среднего плейстоцена.
Позднеплейстоценовый период характеризовался множественными резкими колебаниями климата (Dansgaard et al., 1993; Wohlfarth et al., 2008; Rasmussen et al., 2014 и др.), которые оказали большое влияние на популяции позвоночных животных. В этот относительно короткий геологический период произошли значительные изменения ареалов видов во времени (Sommer, Nadachowski, 2006; Sommer, Zachos, 2009; Маркова и др., 2008; Cooper et al., 2015; Baca et al., 2017) и вымирание многих видов крупных млекопитающих (Stuart, Lister, 2007; Stewart, 2008; Pacher, Stuart, 2009 и др.). Особый интерес вызывает история развития фауны млекопитающих на территории Уральского региона, уникального биогеографического перекрестка Северной Евразии, где в настоящее время обитают представители европейской, сибирской и транспалеарктической фаун (Большаков и др., 2000). Результаты многочисленных исследований позднеплейстоценовой фауны этой территории в сочетании с информацией о развитии окружающей среды дают сложную картину динамики видов в прошлом и вызывают новые проблемные вопросы по отдельным аспектам четвертичной истории Урала.
Среди пещер южной части Южного Урала по богатству фаунистических находок выделяется обнаруженная в 2009 г. на территории национального парка “Башкирия” (Мелеузовский р-н Республики Башкортостан, Россия) пещера Иманай. У входа пещеры и внутри нее обнаружен богатый комплекс останков плейстоцен-голоценовых млекопитающих (Гимранов и др., 2016; Яковлев и др., 2016; Gimranov, Kosintsev, 2020). В отложениях пещеры найдены многочисленные кости крупных пещерных львов и пещерных медведей (Сотникова, Гимранов, 2017; Гимранов и др., 2016, 2017, 2018, 2021). Подробно исследованы термические свойства, гранулометрический, фазовый, химический и нормативно-минеральный составы элювиальных грунтов пещеры, выявлены минералого-геохимические особенности пещерной фоссилизации ископаемых костей пещерных медведей и львов (Силаев и др., 2018, 2020). В отложениях пещеры найден комплекс каменных орудий, отнесенных к среднему палеолиту (мустьерской культуре) (Котов и др., 2020). По результатам изучения костных останков мелких млекопитающих с поверхности пола и из отложений разведочного шурфа пещеры (2013 г.) был сделан вывод, что отложение этого материала (до глубины 65 см) происходило в конце позднего неоплейстоцена и в раннем голоцене с последующим внесением позднеголоценовых костей (Яковлев и др., 2016). В результате последующих раскопок этой пещеры (2016 г.) в отложениях до глубины 120 см были обнаружены многочисленные костные останки мелких млекопитающих, количество которых оказалось существенно выше в нижней половине вскрытых отложений. Было выдвинуто предположение, что нижняя половина вскрытых отложений пещеры образована не позже середины позднего плейстоцена. Данное предположение было основано на результатах радиоуглеродного датирования костей крупных млекопитающих (с глубины 0.0–0.1 м 26 320 ± 1790 GIN 14 244; с глубины 0.1–0.2 м 34 250 ± 120 IGAN 8464, 38210 ± 200 IGAN 8466, 46 260 ± 350 IGAN 8465; с глубины 0.2–0.3 м 31 150 ± 110 IGAN8462), преимущественная локализация которых зафиксирована в верхней половине отложений пещеры.
В данной статье мы приводим результаты исследования ископаемой фауны мелких млекопитающих из раскопа 2016 г. пещеры Иманай. Цели данного исследования – определить временнóй период образования отложений пещеры и охарактеризовать соответствующие этому периоду состав и структуру сообществ мелких млекопитающих (Euliphotyphla, Chiroptera, Lagomorpha, Rodentia) на территории южной части Уральских гор.
Смысл термина “ископаемая фауна” в данной работе трактуется как “набор видов одного конкретного местонахождения” (Агаджанян, 2009). В настоящей работе “типы фауны” рассматриваются как определенные фазы (хронологические этапы) развития фауны, характеризующиеся количественным преобладанием тех или иных таксонов.
МАТЕРИАЛЫ И МЕТОДЫ
Пещера Иманай расположена в 7 км к северо-востоку от поселка Нугуш (53°02′ с.ш., 56°26′ в.д.) (рис. 1). Пещера коридорного типа длиной около 100 м. Первая часть пещеры представляет собой узкий лаз (0.7 × 0.3 м) длиной 18 м, затем следует коридор (2 × 2 м) длиной 75 м, заканчивающийся гротом размером 5 × 6 × 5.6 м (Гимранов и др., 2016). В центральном гроте пещеры заложен раскоп общей площадью 9.5 м2. Отложения состоят из двух слоев: слой 1 суглинок сероватый с известняковой крошкой, попадаются единичные угольки и скопления угольков, отдельные кусочки красной охры, изделия из кремня; мощность слоя – 0.6 м; слой 2 суглинок бурый с единичными камнями известняка и отдельными глыбами; вскрытая мощность – 0.6 м (Гимранов и др., 2021).
Рис. 1.
Географическое положение пещеры Иманай (Южный Урал, Россия) и схема центрального грота пещеры с обозначением исследованных участков раскопа (Г4 и разведочный шурф (черный квадрат)). Серым цветом выделена площадь всего раскопа. Квадраты раскопок обозначены цифрами и буквами.
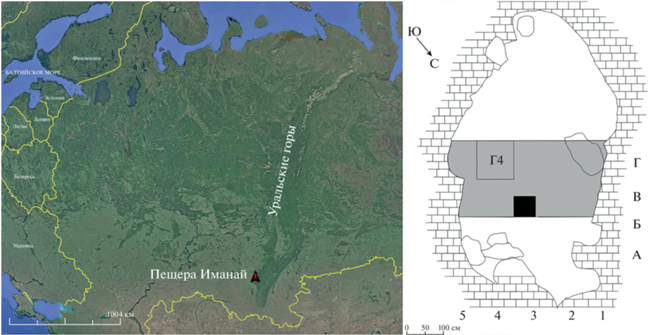
Фауна мелких млекопитающих изучена из квадрата Г4 (рис. 1). Из исследованных отложений 12 условных горизонтов (глубина 0–120 см) этой части центрального грота установлена таксономическая принадлежность 15 430 зубов и челюстей млекопитающих из четырех отрядов Euliphotyphla, Chiroptera, Lagomorpha, Rodentia (табл. 1). Ископаемый костный материал сильно раздроблен и представлен в основном изолированными зубами.
Таблица 1.
Таксономический состав и количество костных останков млекопитающих (Euliphotyphla, Chiroptera, Lagomorpha, Rodentia) из отложений квадрата Г4 пещеры Иманай
| Таксон/глубина отложений, см | 110–120 | 100–110 | 90–100 | 80–90 | 70–80 | 60–70 | 50–60 | 40–50 | 30–40 | 20–30 | 10–20 | 0–10 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Talpa sp. | 1 | 1 | – | 1 | 1 | 1 | – | 1 | – | – | – | 1 |
| Crocidura sp. | 1 | 1 | – | – | – | – | – | – | – | 2 | 1 | 2 |
| Sorex araneus Linnaeus 1758 | – | – | – | – | – | – | 2 | – | – | 2 | 4 | – |
| Sorex isodon Turov 1924 | – | – | – | – | – | – | – | – | – | – | – | 1 |
| Sorex tundrensis Merriam 1900 | – | 4 | 5 | – | 7 | 3 | 4 | 3 | 2 | – | 3 | 11 |
| Sorex minutus Linnaeus 1766 | – | – | – | – | – | – | – | – | – | – | – | 1 |
| Sorex sp. | 1 | 3 | 3 | 2 | 4 | 7 | 19 | 4 | 2 | 6 | 24 | 21 |
| Eptesicus nilssonii (Keyserling & Blasius 1839) | – | – | – | – | – | – | 1 | 1 | – | – | – | 1 |
| Plecotus auritus (Linnaeus 1758) | – | 3 | – | – | – | – | 1 | – | 1 | – | 2 | 2 |
| Chiroptera | 1 | – | 2 | 1 | 4 | 1 | 2 | – | 1 | 1 | 5 | |
| Ochotona sp. | 13 | 102 | 88 | 37 | 158 | 141 | 230 | 68 | 29 | 37 | 68 | 73 |
| Lepus sp. | – | – | – | – | 1 | – | 2 | 3 | – | – | 1 | – |
| Spermophilus sp. | – | – | 2 | – | 3 | 4 | 5 | 1 | – | – | 1 | 3 |
| Sicista subtilis Pallas 1773 | – | – | – | – | – | – | 1 | – | – | – | – | – |
| Sicista sp. | – | – | – | – | – | – | 1 | – | – | – | – | 2 |
| Allactaga major (Kerr 1792) | – | – | 2 | – | 1 | – | 1 | – | – | – | – | – |
| Alactagulus sp. | – | – | – | 2 | 1 | 4 | 10 | 2 | – | – | 1 | – |
| Cricetus cricetus Linnaeus 1758 | 2 | 8 | 10 | 4 | 22 | 5 | 11 | 6 | – | – | 3 | 11 |
| Allocricetulus eversmanni Brandt 1859 | 2 | 4 | 5 | 2 | 1 | 6 | 24 | 1 | 2 | 2 | 1 | 8 |
| Cricetulus migratorius Pallas 1773 | 7 | 2 | 5 | 3 | 15 | 14 | 39 | 2 | 1 | – | – | – |
| Ellobius talpinus Pallas 1770 | 3 | 9 | 9 | 3 | 21 | 12 | 22 | 6 | 6 | 1 | 9 | 12 |
| Lemmus sibiricus Kerr 1792 | – | – | – | – | – | – | – | – | – | – | – | 1 |
| Craseomys rufocanus Sundevall 1846 (M2) | – | 1 | 1 | 1 | – | – | – | – | – | – | 2 | 2 |
| Myodes glareolus Schreber 1780 (M2) | 2 | 6 | 1 | 2 | 4 | 4 | 7 | – | 2 | – | 2 | 4 |
| Myodes rutilus Pallas 1779 (M2) | – | – | 3 | – | – | – | – | – | – | – | – | – |
| Craseomys sp., Myodes sp. | 9 | 31 | 12 | 8 | 20 | 22 | 47 | 10 | 7 | 10 | 22 | 23 |
| Lagurus lagurus Pallas 1773 | 61 | 494 | 410 | 259 | 830 | 837 | 1667 | 262 | 68 | 3 | 31 | 32 |
| Eolagurus luteus Eversmann 1840 | 14 | 27 | 41 | 15 | 52 | 64 | 165 | 52 | 4 | 12 | 1 | – |
| Arvicola amphibius (Linnaeus 1758) | – | 1 | – | – | – | – | 3 | – | 2 | – | 5 | 8 |
| Alexandromys oeconomus (Pallas 1776) (m1) | 1 | 5 | – | 2 | 4 | 5 | 8 | 3 | 2 | – | 3 | – |
| Microtus (Stenocranius) gregalis Pallas 1779 (m1) | 28 | 122 | 98 | 46 | 159 | 167 | 345 | 93 | 20 | 13 | 12 | 20 |
| cf. Microtus (Stenocranius) gregaloides Hinton 1923 (m1) | 1 | 4 | 2 | 2 | 6 | 2 | 4 | 3 | – | – | – | – |
| cf. Microtus (Terricola) arvalidens Kretzoi 1958 (m1) | – | – | – | 1 | – | – | – | – | – | – | – | – |
| Microtus arvalis (Pallas 1779) (m1) | – | 2 | 2 | – | 5 | 1 | – | 3 | 1 | – | 2 | 1 |
| Microtus agrestis (Linnaeus 1761) (m1, M2) | – | 2 | 3 | – | 1 | 2 | 4 | 2 | – | 1 | 2 | 4 |
| Microtus ex gr. agrestis-arvalis (m1) | – | 1 | 6 | 3 | 7 | 3 | 6 | 2 | 1 | – | – | 2 |
| Microtus sp. | 119 | 612 | 466 | 435 | 1161 | 978 | 2159 | 332 | 167 | 85 | 148 | 194 |
| Sylvaemus sp. | – | 1 | – | – | 1 | – | – | – | 4 | 1 | ||
| ∑ (количество останков) | 266 | 1446 | 1176 | 828 | 2488 | 2284 | 4791 | 860 | 318 | 175 | 357 | 441 |
| N (количество таксонов) | 15 | 19 | 17 | 17 | 20 | 18 | 22 | 16 | 13 | 11 | 20 | 21 |
Радиоуглеродное датирование образцов проведено в ЦКП “Лаборатория радиоуглеродного датирования и электронной микроскопии” Института географии РАН, Москва, и Центре прикладных изотопных исследований Университета Джорджии, США. Для отложений квадрата Г4 по костям мелких млекопитающих получено две радиоуглеродные даты: 13 255 ± 60 IGAN 9116 (глубина 50–60 см) и 17 100 ± 50 IGAN 9117 (100–110 см). Приведены не калиброванные даты.
Первые нижние моляры узкочерепных полевок (Microtus (Stenocranius) gregalis) и степных пеструшек (Lagurus lagurus) дифференцированы по морфотипам непарной петли антероконида (Большаков и др., 1980; Яковлев, 2015). Первые нижнекоренные зубы серых полевок (Microtus agrestis, M. arvalis) идентифицированы с помощью классификационных функций (Маркова, Бородин, 2005). Видовая идентификация единственного зуба (M3) представителя трибы Lemmini была проведена по соответствующим методикам (Смирнов и др., 1997; Ponomarev et al., 2015). Оцифровка материала проведена с помощью сканирующего электронного микроскопа VEGA 3 LMH в лаборатории геологии месторождений полезных ископаемых Горного института УрО РАН.
Все выявленные виды грызунов из исследованных отложений пещеры отнесены к определенным экологическим группам, исходя из экологических предпочтений рецентных видов (Маркова, 2008). Выделено 6 групп: степные виды: Spermophilus sp., Sicista subtilis, Microtus gregalis, Lagurus lagurus, Allocricetulus eversmanni, Cricetulus migratorius; полупустынные виды: Allactaga major, Alactagulus sp., Ellobius talpinus, Eolagurus luteus; лесные виды: Sylvaemus sp., Craseomys rufocanus, Myodes sp., Microtus agrestis; луговые: Cricetus cricetus, Microtus arvalis; интразональные виды: Arvicola amphibius, Alexandromys oeconomus; тундровый вид: Lemmus sibiricus.
РЕЗУЛЬТАТЫ
Тафономические особенности. Костный материал из исследованных отложений имеет высокую степень раздробленности. Все кости посткраниального скелета мелких млекопитающих (плечевые, локтевые, лучевые, бедренные, берцовые, тазовые) фрагментированы. Целые черепа и нижние челюсти отсутствуют. Все зубы грызунов представлены в изолированном виде, часть зубов насекомоядных млекопитающих и рукокрылых сохранились во фрагментированных нижних челюстях. Костный материал по всей глубине отложений желтого цвета. Единичные кости имеют внутренние темно-серые пятна и редкие наружные точечные минеральные образования черного цвета. Депигментированные зубы бурозубок единичны. Исходя из высокой степени раздробленности костного материала, можно предположить, что кости мелких млекопитающих имеют экскрементное происхождение. Зоогенные отложения были образованы в результате жизнедеятельности хищных млекопитающих (лисица, песец), использующих внутренний грот пещеры в качестве вре́менного убежища.
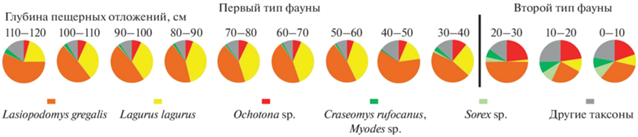
Состав и структура фауны. Наибольшее количество определенных таксонов мелких млекопитающих (18–22) выявлено в верхних условных горизонтах отложений (0–20 см) и в средней части отложений (50–80 см) (табл. 1). По всей глубине отложений преобладают зубы узкочерепной полевки, содоминирующими видами являются степная пеструшка или пищухи (Ochotona sp.). Эти три таксона составляют 57.0–74.7% от общего количества определенных зубов и челюстей мелких млекопитающих в отложениях верхних условных горизонтов (0–30 см) и 81.5–90.5% – в отложениях средних и нижних условных горизонтах (30–120 см). В верхних условных горизонтах (0–30 см) содоминантами узкочерепных полевок являются пищухи (19.8–23.4% от общего количества зубов и челюстей всех видов мелких млекопитающих), в средних и нижних условных горизонтах (30–120 см) содоминирующий вид другой – степная пеструшка (16.4–40.5%), а доля зубов пищух становится существенно меньше (4.4–10.9%). Кроме этих различий, можно отметить бо́льшую долю бурозубок и лесных полевок в отложениях верхних горизонтов по сравнению с таковой в средних и нижних горизонтах (3.8–9.0 и 6.3–8.7%, 0.0–1.3 и 0.9–3.7%, соответственно). Зубы желтой пеструшки (Eolagurus luteus) единичны в верхних горизонтах (0–20 см) и составляют от 1.5 до 7.6% в нижележащих отложениях. Долевое участие костных останков большинства остальных таксонов, определенных по зубам, составляет менее 1% по всей глубине отложений.
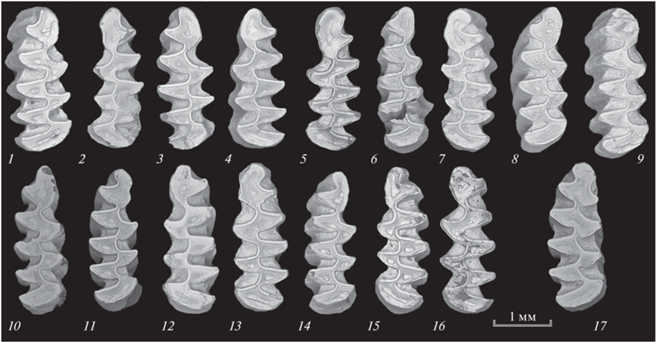
Среди зубов грызунов в нижней половине отложений обнаружены бескорневые цементные первые нижнекоренные зубы (m1) с широким слиянием антероконидных треугольников Т4 и Т5 (рис. 2). Такое “питимисное” строение жевательной поверхности с определенными формами непарной петли антероконидного отдела характерны для древних полевок Microtus (Stenocranius) gregaloides и M. (Terricola) arvalidens из фаун первой половины раннего плейстоцена–второй половины среднего плейстоцена (Maul, Мarkova, 2007). Среди бескорневых цементных третьих верхнекоренных зубов (M3) полевок обнаружены зубы со слиянием T2 и T3. По сохранности и цветовым характеристикам эти зубы аналогичны зубам других видов мелких млекопитающих, обнаруженных в исследованных отложениях пещеры.
Рис. 2.
Зубы полевок из отложений квадрата Г4 пещеры Иманай. cf. Microtus (Stenocranius) gregaloides (Hinton 1923): 1, 2 – глубина 40–50 см; 3–5 – 50–60 см; 6–9 – 70–80 см; 10, 11 – 80–90 см; 12 – 90–100 см; 13–15 – 100–110 см; 16 – 110–120 см. cf. Microtus (Terricola) arvalidens Kretzoi 1958: 17 – 80–90 см.
Фауну мелких млекопитающих из исследованных отложений по долевому участию доминирующих видов условно можно разделить на 2 типа (рис. 3). В первом типе фауны (отложения условных горизонтов с глубины 30–120 см) основную массу идентифицированных зубов составляют зубы узкочерепных полевок и степных пеструшек, лесные виды представлены единичными зубами лесных полевок и фрагментами нижних челюстей бурозубок. Второй тип характерен для отложений верхних условных горизонтов (глубина 0–30 см), где доминируют зубы узкочерепной полевки и пищухи, условно лесные виды (лесные полевки и бурозубки) обычны.
Рис. 3.
Два типа ископаемой фауны, дифференцированных по соотношениям количества костных останков доминирующих таксонов мелких млекопитающих. Пещера Иманай, квадрат Г4.
Таксономические замечания
Отряд Rodentia Bowdich 1821
Семейство Cricetidae Fischer von Waldheim 1817
Подсемейство Arvicolinae Gray 1821
Род Microtus Schrank 1798
Подрод Stenocranius Kastshenko 1901
Microtus (Stenocranius) gregalis Pallas 1779
Материал. Определены 1123 изолированных первых нижнекоренных зуба (m1), измерены 236 m1.
Описание и сравнение. Бескорневые зубы с отложениями наружного цемента во входящих углах. Микротусный тип эмали (толщина эмали заметно тоньше на выпуклых стенках петель). Размерный диапазон длины зубов от 2.15 до 3.30 мм.
В исследуемых выборках зубов зафиксировано однотипное распределение частот всех групп морфотипов (рис. 4; табл. 2). В выборках преобладают (более 50% от всех m1 вида) зубы простого строения непарной петли антероконидного комплекса, без входящего угла на буккальной стороне (“грегалоидный” морфотип). Зубы со сложным строением петли, на буккальной стороне которой входящий угол составляет более 20° (“микротидный” морфотип), немногочисленны. Зубы со сложной морфой составляют около 10% или менее, за исключением выборки из отложений самого верхнего горизонта. Преобладание “грегалоидного” морфотипа зафиксировано и в позднеледниковых-раннеголоценовых выборках зубов узкочерепной полевки из пещеры Байслан-Таш и Максютовского грота, расположенных приблизительно в 30 км юго-восточнее пещеры Иманай на правом берегу р. Белой (табл. 2). Ранее доминирование зубов с “грегалоидным” морфотипом зафиксировано в выборках зубов вида из отложений среднего плейстоцена и первой половины позднего плейстоцена местонахождений Южного Урала (Смирнов и др., 1990; Фадеева и др., 2019; Яковлев, 2020). Однако позднеледниковые выборки зубов вида из местонахождений севера горной части Южного Урала имеют несколько другой морфологический облик с преобладанием “грегалоидно-микротидного” морфотипа (Смирнов и др., 1990). В выборках зубов современных подвидов узкочерепных полевок с территории Урала (за исключением выборки “Ямал”) доминируют зубы сложного строения (“микротидный” морфотип) (Головачев и др., 2001).
Рис. 4.
Соотношение морфотипов первых нижнекоренных зубов (m1) Microtus (Stenocranius) gregalis. Пещера Иманай, квадрат Г4.
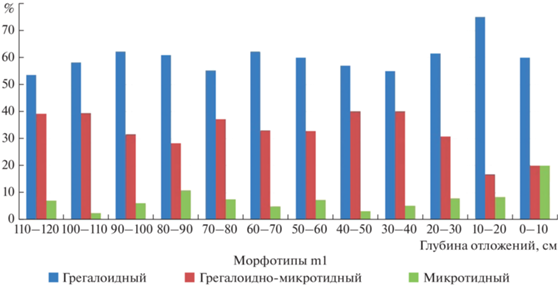
Таблица 2.
Соотношение морфотипов и размеры первого нижнего моляра (m1) в выборках зубов Microtus (Stenocranius) gregalis из ископаемых и современных местонахождений Урала
| Местонахождение | Слой, глубина, cм | Радиоуглеродная датировка/период | N | Морфотипы m1 | Длина m1 (мм) (минимум–среднее–максимум [n]) |
Источник | ||
|---|---|---|---|---|---|---|---|---|
| грегалоидный | грегалоидно-микротидный | микротидный | ||||||
| Ямал (M. g. major) | – | Современность | 59 | 6.8 | 55.9 | 37.3 | 2.68–2.96–3.30 [59] | Головачев и др., 2001 |
| Красный Камень (M. g. major) | – | 109 | – | 23.8 | 76.2 | 2.38–2.69–3.05 [107] | ||
| Паюта (M. g. major) | – | 81 | 1.2 | 42.0 | 56.8 | 2.55–2.94–3.23 [81] | ||
| Бреды (M. g. gregalis) | – | 42 | – | 33.3 | 66.7 | 2.25–2.51–2.90 [42] | ||
| Камышловский (M. g. gregalis) | – | 42 | 4.7 | 40.5 | 54.8 | 2.08–2.50–2.88 [42] | ||
| Устиново | Сл. 1 | 4380 ± 170 ИЭРЖ 47 | 29 | 6.9 | 65.5 | 27.6 | 2.30–2.61–2.80 [22 ] | Смирнов и др., 1990 |
| Сл. 3 | 12 400 ± 300 ИЭРЖ 49 | 24 | 20.8 | 55.3 | 23.9 | 2.35–2.75–3.10 [16 ] | ||
| Прижим II | Сл. 1–3 | 16 650 ± 400 ИЭРЖ 32, 17 070 ± 1017 IEMAE 700 | 68 | 33.8 | 53.0 | 13.2 | 2.35–2.74–3.15 [67] | |
| Сл. 6–8 | 21 085 ± 630 ИЭРЖ 37 | 86 | 30.2 | 55.9 | 13.9 | 2.40–2.74–3.25 [83] | ||
| Игнатиевская II | Cл. 2 | 14 038 ± 490 IEMAE 366 | 177 | 30.5 | 54.2 | 15.3 | 2.53–2.69–3.25 [175] | |
| Байслан-Таш | 225–450 | 9616 ± 62 IEMAE 1340, 13 560 ± 250 GIN 10 853 |
61 | 57.4 | 27.9 | 14.7 | 2.50–2.73–3.13 [44] | Yakovlev et al., 2006; Danukalova et al., 2011; данное исследование |
| Максютовский грот | Сл. 2 | 15 650 ± 150 SOAN 7755 | 27 | 44.4 | 40.7 | 14.8 | 2.50–2.71–3.02 [15 ] | |
| Иманай Г4 | 50–60 | 13 255 ± 60 IGAN 9116 | 345 | 60.0 | 32.8 | 7.2 | 2.15–2.65–3.30 [141] | |
| 100–110 | 17 100 ± 50 IGAN 9117 | 122 | 58.2 | 39.3 | 2.5 | 2.30–2.63–2.95 [95] | ||
| Игнатиевская V раскоп 1985 г. |
160–195 | >27 500 ИЭРЖ 21 | 34 | 47.1 | 41.2 | 11.7 | 2.40–2.71–3.15 [34] | Смирнов и др., 1990 |
| 195–255 | 64 | 39.1 | 53.0 | 7.8 | 2.30–2.66–3.10 [64] | |||
| 255–315 | 39 | 61.5 | 30.8 | 7.7 | 2.35–2.61–3.00 [40] | |||
| 315–400 | 201 | 42.8 | 50.7 | 6.5 | 2.25–2.60–3.30 [196] | |||
| раскоп 2014 г. | 325–400 | Конец среднего
плейстоцена (?)–начало позднего плейстоцена |
594 | 52.1 | 37.1 | 10.8 | 2.30–2.64–3.30 [254] | Фадеева и др., 2019 |
| 400–520 | 311 | 73.2 | 23.2 | 3.6 | 2.20–2.58–2.90 [105] | |||
| Горнова | – | Первая половина позднего плейстоцена | 34 | 38.2 | 32.4 | 29.4 | 2.35–2.61–2.87 [13 ] | Яковлев, 2020 |
| Климовка | – | Средний плейстоцен | 27 | 37.1 | 44.4 | 18.5 | 2.3–2.67–3.2 [15 ] | |
| Груздевка | – | Средний плейстоцен | 21 | 52.4 | 23.8 | 23.8 | 2.5–2.71–3.0 [7 ] | |
| Красный Яр | – | Первая половина среднего плейстоцена | 77 | 59.7 | 27.3 | 13.0 | 2.45–2.68–3.0 [40] | |
Пределы длины зубов из ископаемых выборок местонахождений южной части Южного Урала (Климовка, Максютовский грот, пещеры Байслан-Таш и Иманай) составляют 2.15–3.30 мм. Нижний предел размерного ряда выше (2.30 мм) в ископаемых выборках из местонахождений северной части Южного Урала (Груздевка, Горнова, Красный Яр, пещеры Игнатиевская и Прижим II, навес Устиново). Размерные пределы длины зубов ископаемых узкочерепных полевок Южного Урала ближе всего к таковым современной выборки северного подвида M. g. gregalis из окрестностей горы Красный Камень (Северный Урал) (табл. 2).
Род Lagurus Gloger 1841
Lagurus lagurus (Pallas 1773)
Материал. Определены 4 954 изолированных коренных зуба, измерены 233 m1.
Описание и сравнение. Бескорневые, бесцементные коренные зубы. Эмаль микротусного типа. Размерный диапазон длины зубов от 2.10 до 2.65 мм.
Выборки первых нижних моляров степных пеструшек из отложений пещеры Иманай отличаются от всех ранее исследованных ископаемых выборок вида с территории Южного Урала высокой долей (28.0–51.6%) “транзиенсногo” морфотипа, который характерен для зубов ранне-среднеплейстоценового вида Lagurus transiens Janossy 1962 (рис. 5; табл. 3). Для территории Урала ранее обнаружены ископаемые выборки зубов степных пеструшек, где максимальное количество зубов “транзиенсного” строения составляет не более трети от количества всех зубов в выборке (табл. 3).
Рис. 5.
Соотношение морфотипов первых нижнекоренных зубов (m1) Lagurus lagurus. Пещера Иманай, квадрат Г4.
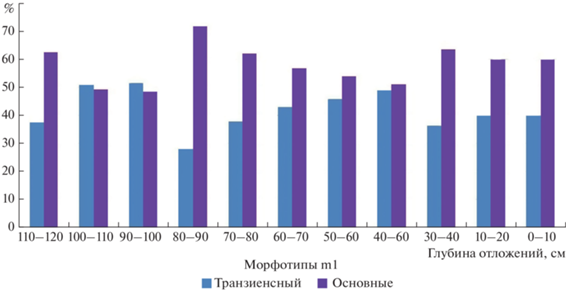
Таблица 3.
Соотношение морфотипов и размеры первого нижнего моляра (m1) в выборках зубов Lagurus lagurus из местонахождений Южного Урала
| Местонахождение | Слой, глубина, см |
Радиоуглеродная датировка/период | N | Морфотипы | Длина m1 (мм) (минимум–среднее–максимум [n]) |
Источник | ||
|---|---|---|---|---|---|---|---|---|
| транзиенсный | основные | |||||||
| переходный | лагурусный | |||||||
| Поволжье | – | Современность | 29 | 6.7 | 66.7 | 26.6 | 2.17–2.40–2.69 [29] | Малеева, Воробьева, 1973 |
| Северный Казахстан | – | 30 | 6.7 | 73.3 | 20.0 | 1.99–2.36–2.69 [30] | ||
| Байслан-Таш | 225–450 | 9616 ± 62 IEMAE 1340; 13 560 ± 250 GIN 10853 |
29 | 21.1 | 39.5 | 39.5 | 2.32–2.63–3.00 [29] | Yakovlev et al., 2006; Danukalova et al., 2011; данное исследование |
| Максютовский грот | Сл. 2 | 15 650 ± 150 SOAN 7755 | 8 | 12.5 | 25.0 | 62.5 | 2.55–2.69–2.90 [6 ] | |
| Иманай Г4 | 50–60 | 13 255 ± 60 IGAN 9116 | 174 | 43.0 | 42.3 | 14.7 | 2.10–2.45–2.65 [86] | |
| 100–110 | 17 100 ± 50 IGAN 9117 | 59 | 50.8 | 28.8 | 20.3 | 2.20–2.45–2.65 [35] | ||
| Игнатиевская V (раскоп 2014 г.) |
325–400 | Конец среднего плейстоцена (?)–начало позднего плейстоцена | 70 | 34.3 | 17.1 | 48.6 | 2.25–2.52–2.90 [36] | Фадеева и др., 2019 |
| 400–430 | 19 | 21.1 | 21.1 | 57.9 | 2.45–2.54–2.70 [6 ] | |||
| Горнова | – | Первая половина позднего плейстоцена | 39 | 2.6 | 30.8 | 66.6 | 2.35–2.56–2.75 [25 ] | Яковлев, 2015 |
| Климовка | – | Средний плейстоцен | 16 | 18.8 | 31.3 | 50.0 | 2.30–2.65–2.87 [10 ] | |
| Груздевка | – | Средний плейстоцен | 18 | 16.7 | 16.7 | 66.7 | 2.27–2.55–2.78 [9 ] | |
| Красный Яр | – | Первая половина среднего плейстоцена | 519 | 18.1 | 41.4 | 40.5 | 2.20–2.61–3.02 [100] | |
Первые нижнекоренные зубы степных пеструшек из отложений пещеры Иманай отличаются от других ископаемых выборок вида из местонахождений Южного Урала сравнительно мелкими размерами (табл. 3). Схожие размеры m1 характерны для современных степных пеструшек с территорий Поволжья и Казахстана (Малеева, Воробьева, 1973), однако в этих выборках доля зубов “транзиенсного” морфотипа существенно меньше (<7%).
ОБСУЖДЕНИЕ
Пещера Иманай входит в южную группу пещер горной части Южного Урала, расположенных на широтных участках долин рек Белая и Нугуш (Danukalova et al., 2011, 2020). Для этой территории до сих пор были известны фауны мелких млекопитающих второй половины позднего плейстоцена, среднего и позднего голоцена (Данукалова и др., 2002; Яковлев, 2003, 2014; Яковлев и др., 2004; Yakovlev et al., 2006; Бачура, Косинцев, 2010; Kosintsev, Bachura, 2013; Косинцев и др., 2018 и др.). В позднеплейстоценовых и среднеголоценовых фаунах этой территории доминируют узкочерепные полевки, степные пеструшки и пищухи, в позднеголоценовых – лесные полевки. В конце позднего голоцена с этой территории исчезают белозубки, пищухи, тушканчики, серые хомячки, хомячки Эверсмана, слепушонки, желтые пеструшки, степные пеструшки и узкочерепные полевки. В настоящее время здесь обитают только представители лесных и интразональных видов мелких млекопитающих (Danukalova et al., 2011).
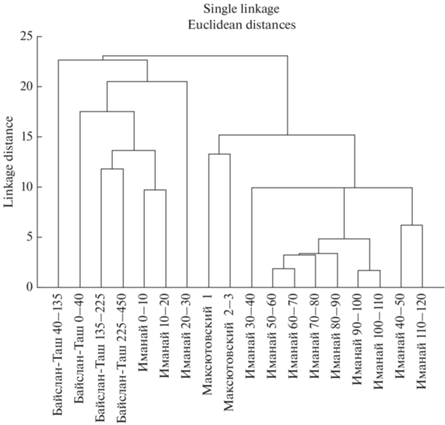
Выделенные по долевому участию доминирующих видов два типа фауны пещеры Иманай (рис. 6) сравнены с позднеледниковыми и голоценовыми фаунами других местонахождений, расположенных на берегу р. Белой, приблизительно в 30 км юго-восточнее пещеры.
Рис. 6.
Кластерный анализ ископаемых фаун южной группы пещер горной части Южного Урала (пещера Иманай, квадрат Г4 (глубины в см); пещера Байслан-Таш (глубины в см); Максютовский грот (номера слоев)).
Первый тип фауны (нижние и средние условные горизонты) пещеры Иманай и фауна нижних отложений Максютовского грота (сл. 2 – 15 650 ± ± 150 SOAN 7755) (Danukalova et al., 2011) образованы в позднеледниковье и характеризуются резким преобладанием степных видов грызунов, доминированием узкочерепной полевки и содоминированием степной пеструшки. Костные останки этих видов грызунов составляют 70.9–84.4% в средних и нижних отложениях Иманая и 62.8–68.6% в отложениях Максютовского грота.
Фауна второго типа (верхние условные горизонты) пещеры Иманай и фауна третьего (135–225 см) и четвертого (225–450 см) слоев исследованных отложений пещеры Байслан-Таш (52°54′ c.ш., 56°51′ в.д.) очень близки. В пещере Байслан-Таш третий слой датирован средним голоценом (7140 ± 170 GIN 10854), а самый нижний четвертый слой отнесен к позднеледниковью–раннему голоцену (9616 ± 62 IEMAE 1340; 13 560 ± ± 250 GIN 10853) (Yakovlev et al., 2006). Сравниваемые фауны характеризуются доминированием узкочерепных полевок, содоминированием пищух и степных пеструшек, значительными долями бурозубок и лесных полевок.
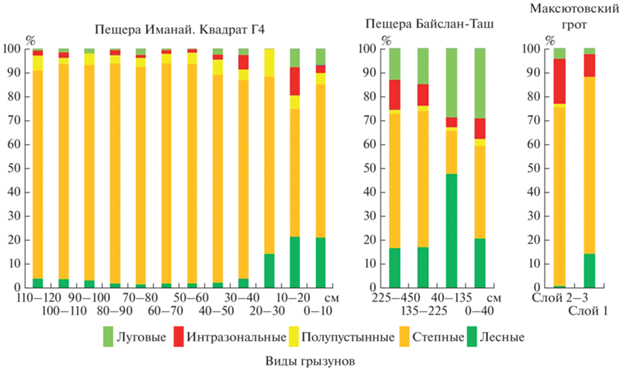
Для всех рассмотренных фаун местонахождений южной группы горной части Южного Урала характерно присутствие степных, лесных, интразональных, луговых и полупустынных видов грызунов. Единственный зуб тундрового вида Lemmus sibiricus обнаружен только в верхних отложениях пещеры Иманай. В этих верхних отложениях пещеры, так же как и в отложениях пещеры Байслан-Таш, доля костных останков луговых и лесных видов составляет около 30% и более (рис. 7). Среди костных останков мелких млекопитающих в средних и нижних слоях отложений квадрата Г4 пещеры Иманай значительную долю составляют степные виды (83.0–92.2%), в то время как количество останков лесных и луговых представителей сравнительно мало (2.2–6.5%) (рис. 7). Такие соотношения костных останков видов из различных экологических группировок из этих отложений пещеры Иманай более близки к таковым из отложений Максютовского грота (степные 74.6%; лесные и луговые 5.3–16.6%). Большая доля интразональных видов (18.7%) в позднеледниковых отложениях Максютовского грота сравнительно с таковой в средних и нижних отложениях пещеры Иманай (до 6.1%), вероятно, обусловлена расположением грота на берегу реки.
Рис. 7.
Соотношения количеств костных останков грызунов из различных экологических группировок (в соответствии с их современными местообитаниями) в отложениях южной группы пещер горной части Южного Урала.
Морфотипические характеристики выборок ископаемых первых нижнекоренных зубов узкочерепной полевки из пещер Иманай, Байслан-Таш и Максютовского грота очень близки (табл. 2). Морфология зубов этого вида из отложений второй половины позднего плейстоцена из местонахождений южной группы пещер горной части Южного Урала (Иманай, Байслан-Таш, Максютовский грот) сходна с таковой из отложений среднего плейстоцена и первой половины позднего плейстоцена местонахождений северной группы равнинной и горной части Южного Урала (Груздевка, Красный Яр, Горнова, пещера Игнатиевская). Все эти выборки ископаемых зубов вида характеризуются большой долей зубов простого строения антероконидного отдела, что отличает их от ископаемых выборок второй половины позднего плейстоцена–голоцена местонахождений северной группы (пещера Игнатиевская, Прижим II, Устиново) и современных выборок зубов узкочерепных полевок с территорий Северного и Южного Урала.
Выборки первых нижнекоренных зубов степных пеструшек из отложений пещеры Иманай характеризуются высокой долей зубов “транзиенсного” морфотипа (28.0–59.7%). Зубы этого морфотипа составляют около 30% в объединенной выборке зубов вида из отложений первой половины позднего плейстоцена пещеры Игнатиевская. Доли таких зубов в отложениях пещеры Байслан-Таш и Максютовского грота существенно ниже. Кроме того, зубы степных пеструшек из отложений пещеры Иманай имеют более мелкие размеры по сравнению с зубами из всех других ранее изученных выборок из местонахождений Южного Урала. Для понимания причин существенной разницы в морфологических характеристиках первых нижнекоренных зубов степных пеструшек из позднеплейстоценовых местонахождений Южного Урала требуются дополнительные исследования.
ЗАКЛЮЧЕНИЕ
В исследованных отложениях пещеры Иманай идентифицированы 32 таксона млекопитающих из четырех отрядов (Eulipotyphla, Chiroptera, Lagomorpha, Rodentia). Во всех условных горизонтах отложений пещеры Иманай установлен единственный доминирующий вид – Microtus gregalis. Нижняя и средняя части отложений характеризуются содоминированием Lagurus lagurus, в верхних горизонтах отложений иной содоминант – Ochotona sp. Нижняя и средняя части отложений пещеры Иманай, судя по радиоуглеродным датам, образованы в позднеледниковье. Состав и структура фауны из этих отложений (первый тип фауны) очень близки к таковым позднеледниковой ископаемой фауны Максютовского грота. В фауне (второй тип) верхних условных горизонтов пещеры сравнительно более высокая доля лесных видов, схожая с таковой в фауне нижних горизонтов отложений пещеры Байслан-Таш (позднеледниковье–ранний голоцен). Выборки первых нижнекоренных зубов узкочерепных полевок из позднеледниковых отложений южной группы местонахождений горной части Южного Урала (пещеры Иманай, Байслан-Таш, Максютовский грот) характеризуются высокой долей зубов простого строения (“грегалоидный” морфотип). Однако выборки зубов степных пеструшек из отложений пещеры Иманай отличаются от других позднеледниковых выборок местонахождений Южного Урала мелкими размерами и сравнительно высокой долей зубов “транзиенсного” морфотипа. К особенностям ископаемой фауны пещеры Иманай относятся и находки зубов с “питимисным” строением жевательной поверхности, характерным для древних (ранний–средний плейстоцен) видов полевок, но их идентичная сохранность с зубами других видов мелких млекопитающих из отложений исключает версию переотложенности материала.
Исходя из составов и структур выделенных типов фауны, можно заключить, что в южной части Уральских гор в позднеледниковье существовали открытые степные ландшафты, которые, предположительно, в конце этого периода или в раннем голоцене трансформировались в лесостепные.
Список литературы
Агаджанян А.К., 2009. Мелкие млекопитающие плиоцена-плейстоцена Русской равнины // Труды Палеонтологического института РАН. Т. 289. С. 1− 676.
Бачура О.П., Косинцев П.А., 2010. Фауны млекопитающих в позднем плейстоцене и голоцене на Южном Урале // Вестник Оренбургского государственного университета. № 12 (118). С. 42–48.
Большаков В.Н., Васильева И.А., Малеева А.Г., 1980. Морфотипическая изменчивость зубов полевок. М.: Наука. 140 с.
Большаков В.Н., Бердюгин К.И., Васильева И.А., Кузнецова И.А., 2000. Млекопитающие Свердловской области. Справочник-определитель. Екатеринбург: “Екатеринбург”, 2000. 240 с.
Гимранов Д.О., Котов В.Г., Румянцев М.М., Яковлев А.Г., Сотникова М.В., Нурмухаметов И.М., Сатаев Р.М., Косинцев П.А., 2016. Пещера Иманай – новое палеонтологическое и археологическое местонахождение на Южном Урале // 100-летие Палеонтологического общества России. Проблемы и перспективы палеонтологических исследований: материалы 62-й сессии Палеонтологического общества при РАН. СПб.: ВСЕГЕИ. С. 231–233.
Гимранов Д.О., Косинцев П.А., Нурмухаметов И.М., Некрасов А.Е., 2017. Первая находка дикообраза (Hystrix sp.) и кабана (Sus scrofa) в позднем плейстоцене Южного Урала // Фундаментальные проблемы квартера: Итоги изучения и основные направления исследований: материалы X Всерос. совещания по изучению четвертичного периода. М.: Геос. С. 89.
Гимранов Д.О., Котов В.Г., Румянцев М.М., Силаев В.И., Яковлев А.Г., Яковлева Т.И., Зеленков Н.В., Сотникова М.В., Девяшин М.М., Пластеева Н.А., Зарецкая Н.Е., Нурмухаметов И.М., Смирнов Н.Г., Косинцев П.А., 2018. Крупнейшее в Евразии захоронение ископаемых львов (Carnivora, Filidae, Pantera (Leo) ex gr. fossilis spelaea) // Доклады РАН. Т. 482 (2). С. 234–237.
Гимранов Д.О., Косинцев П.А., Бачура О.П., Жилин М.Г., Котов В.Г., Румянцев М.М., 2021. Малый пещерный медведь (Ursus ex gr. savini-rossicus) как объект охоты древнего человека // Вестник археологии, антропологии и этнографии. Т. 2 (53). С. 5–14.
Головачев И.Б., Смирнов Н.Г., Добышева Э.В., Пономарев Д.В., 2001. К истории современных подвидов узкочерепной полевки // Современные проблемы популяционной, исторической и прикладной экологии. Материалы конф. молодых ученых. Екатеринбург. С. 49–57.
Данукалова Г.А., Яковлев А.Г., Алимбекова Л.И., Косинцев П.А., Морозова (Осипова) Е.М., Еремеев А.А., 2002. Биостратиграфия четвертичных отложений пещер и речных террас широтного течения р. Белой // Экологические аспекты Юмагузинского водохранилища. Уфа: Гилем. С. 32–57.
Косинцев П.А., Котов В.Г., Пантелеев А.В., Яковлев А.Г., 2018. Использование известнякового сырья в верхнем палеолите Урала (по материалам стоянки в пещере Балатукай) // Вестник Пермского университета. История. Т. 1 (40). С. 5–19.
Котов В.Г., Румянцев М.М., Гимранов Д.О., 2020. Стоянка среднего палеолита в пещере Иманай-1 на Южном Урале: предварительные итоги археологических исследований // Oriental Studies. Т. 13 (5). С. 1271–1291.
Малеева А.Г., Воробьева Т.Д., 1973. Степная пеструшка (Lagurus lagurus Pall.) из состава “смешанной фауны” юга Тюменской области // Фауна Европейского Севера, Урала и Западной Сибири. Свердловск. С. 49–66.
Маркова А.К., 2008. Индикаторные виды и экологические группы млекопитающих // Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л. н.). Москва: Товарищество научных изданий КМК. С. 29–39.
Маркова А.К., ван Кольфсхотен Т., Бохнкке Ш., Косинцев П.А., Мол И., Пузаченко А.Ю., Симакова А.Н., Смирнов Н.Г., Верпоорте А., Головачев И.Б., 2008. Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л. н.). М.: Товарищество научных изданий КМК. 556 с.
Маркова Е.А., Бородин А.В., 2005. Определение видовой принадлежности полевок подрода Microtus Schrank, 1798 Урала и Западной Сибири по промерам переднего нижнего зуба // Фауны Урала и Сибири в плейстоцене и голоцене. Челябинск: Рифей. С. 2–10.
Силаев В.И., Симакова Ю.С., Паршукова М.Н., Гимранов Д.О., 2018. Костеносные элювиальные грунты в Иманайской пещере на Южном Урале // Проблемы минералогии, петрографии и металлогении. Научные чтения памяти П.Н. Чирвинского: сборник статей. Пермь: Пермский гос. ун-т. Вып. 21. С. 168–184.
Силаев В.И., Паршукова М.Н., Гимранов Д.О., Филиппов В.Н., Киселева Д.В., Смолева И.В., Тропников Е.М., Хазов А.Ф., 2020. Минералого-геохимические особенности пещерной фоссилизации ископаемых костей на примере пещеры Иманай (Южный Урал) // Вестник Пермского университета. Геология. Т. 19 (4). С. 323–358.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А., Панова Н.К., Коробейников Ю.И., Ольшванг В.Н., Ерохин Н.Г., Быкова Г.В., 1990. Историческая экология животных гор Южного Урала. Свердловск: УрО АН СССР. 244 с.
Смирнов Н.Г., Головачев И.Д., Бачура О.П., Кузнецова И.А., Чепраков М.Ю., 1997. Сложные случаи определения зубов грызунов из отложений позднего плейстоцена и голоцена тундровых районов Северной Евразии // Материалы по истории и современному состоянию фауны севера Западной Сибири. Челябинск: Рифей. С. 60–90.
Сотникова М.В., Гимранов Д.О., 2017. О находках гигантских львов в мустьерских слоях (поздний плейстоцен) пещеры Иманай (Южный Урал) // Интергративная палеонтология: перспективы развития для геологических целей: материалы LXIII сессии Палеонтологического общества при РАН. СПб.: ВСЕГЕИ. С. 207–208.
Фадеева Т.В., Косинцев П.А., Гимранов Д.О., 2019. Млекопитающие горной части Южного Урала в последнее межледниковье // Зоологический журнал. Т. 98. Вып. 11. С. 1304–1322.
Яковлев А.Г., 2003. Микропалеотериологические исследования неоплейстоцена и голоцена Южного Предуралья и западного макросклона Южного Урала // Четвертичная палеозоология на Урале: Сб. научных трудов. Екатеринбург: Изд-во Уральского ун-та. С. 116–122.
Яковлев А.Г., 2014. Поздненеоплейстоценовая фауна мелких млекопитающих из местонахождения в пещере Кульюрт-Тамак (Южный Урал) // Геологический сборник. № 11. Уфа. С. 84–85.
Яковлев А.Г., 2015. Морфологическая характеристика моляров степных пеструшек (Lagurus lagurus Pallas, 1773) из неоплейстоценовых местонахождений Южного Предуралья // Геологический сборник. № 12. С. 56−61.
Яковлев А.Г., Данукалова Г.А., Яковлева Т.И., Алимбекова Л.И., Морозова (Осипова) Е.М., 2004. Биостратиграфическая характеристика голоценовых отложений местонахождения “Грот Ташмурун” (Южный Урал) // Геологический сборник. № 4. Уфа. С. 101–105.
Яковлев А.Г., Яковлева Т.И., Гимранов Д.О., 2016. Мелкие позвоночные (земноводные, пресмыкающиеся и млекопитающие) из местонахождения Иманай 1 (Южный Урал) // Геология, полезные ископаемые и проблемы геоэкологии Башкортостана, Урала и сопредельных территорий: Материалы и доклады. 11-я Межрегиональная науч.-практ. конференция, посвященная 65-летию Института геологии УНЦ РАН, Уфа, 17–19 мая 2016 г. Уфа: ДизайнПресс. С. 81–83.
Яковлев А.Г., 2020. Морфологическая характеристика первых нижних коренных зубов (m1) узкочерепных полевок Microtus (Stenocranius) gregalis Pallas, 1779 из неоплейстоценовых местонахождений Южного Предуралья) // Геологический вестник. № 2. С. 39–44.
Baca M., Nadachowski A., Lipecki G., Mackiewicz P., Marciszak A., Popović D., Socha P., Stefaniak K., Wojtal P., 2017. Impact of climatic changes in the Late Pleistocene on migrations and extinctions of mammals in Europe: four case studies // Geological Quarterly. V. 61. P. 291– 304.
Cooper A., Turney C., Hughen K.A., Brook B.W., McDonald H.G., Bradshaw C.J., 2015. Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover // Science. V. 349. P. 602–606.
Dansgaard W., Johnsen S.J., Clausen H.B., Dahl-Jensen D., Gundestrup N.S., Hammer C.U., Hvidberg C.S., Steffensen J.P., Sveinbjornsdottir A.E., Jouzel J., Bond G., 1993. Evidence for general instability of past climate from a 250-kyr ice-core record // Nature. V. 364. P. 218–220.
Danukalova G., Yakovlev A., Osipova E., Alimbekova L., Yakovleva T., Kosintsev P., 2011. Biostratigraphy of the Late Upper Pleistocene (Upper Neopleistocene) to Holocene deposits of the Belaya River valley (Southern Urals, Russia) // Quaternary International. V. 231 (1–2). P. 28–43.
Danukalova G., Kosintsev P., Yakovlev A., Yakovleva T., Osipova E., Kurmanov R., van Kolfschoten T., Izvarin E., 2020. Quaternary deposits and biostratigraphy in caves and grottoes located in the Southern Urals (Russia) // Quaternary International. V. 546. P. 84–124.
Gimranov D.O., Kosintsev P.A., 2020. Quaternary large mammals from the Imanay Cave // Quaternary International. V. 546. P. 125‒134.
Kosintsev P.A., Bachura O.P., 2013. Late Pleistocene and Holocene mammal fauna of the Southern Urals // Quaternary International. V. 284. P. 161–170.
Maul L.C., Markova A.K., 2007. Similarity and regional differences in Quaternary arvicolid evolution in Central and Eastern Europe // Quaternary International. V. 160. P. 81–99.
Pacher M., Stuart A.J., 2009. Extinction chronology and palaeobiology of the cave bear Ursus spelaeus // Boreas. V. 38. P. 189–206.
Ponomarev D., Puzachenko A., Isaychev K., 2015. Morphotypic variability of masticatory surface pattern of molars in the recent and Pleistocene Lemmus and Myopus (Rodentia, Cricetidae) of Europe and Western Siberia // Acta Zoologica. V. 96. P. 14–29.
Rasmussen S.O., Bigler M., Blockley S.P., Blunier T., Buchardt S.L., Clausen H.B., Cvijanovic I., Dahl-Jensen D., Johnsen S.J., Fischer H., Gkinis V., Guillevic M., Hoek W.Z., Lowe J.J., Pedro J.B., Popp T., Seierstad I.K., Steffensen J.P., Svensson A.M., Vallelonga P., Vinther B.M., Walker M.J.C., Wheatley J.J., Winstrup M., 2014. A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: refining and extending the INTIMATE event stratigraphy // Quaternary Science Reviews. V. 106. P. 14–28.
Sommer R.S., Nadachowski A., 2006. Glacial refugia of mammals in Europe: evidence from fossil records // Mammal Review. V. 36. P. 251– 265.
Sommer R.S., Zachos F.E., 2009. Fossil evidence and phylogeography of temperate species: “glacial refugia” and postglacial recolonization // Journal of Biogeography. V. 36. P. 2013–2020.
Stewart J.R., 2008. The progressive effect of the individualistic response of species to Quaternary climate change: an analysis of British mammalian faunas // Quaternary Science Reviews. V. 27. P. 2499–2508.
Stuart A.J., Lister A.M., 2007. Patterns of Late Quaternary megafaunal extinctions in Europe and northern Asia // Courier Forschingsinstitut Senckenberg. V. 259. P. 289– 299.
Wohlfarth B., Veres D., Ampel L., Lacourse T., Blaauw M., Preusser F., Andrieu-Ponel V., Keravis D., Lallier-Verges E., Björck S., Davies S., de Beaulieu J.L., Risberg J., Hormes A., Kasper H.U., Possnert G., Reille M., Thouveny N., Zander A., 2008. Rapid ecosystem response to abrupt climate changes during the last glacial period in Western Europe, 40–16 kyr BP // Geology. V. 36. P. 407–410.
Yakovlev A., Danukalova G., Kosintsev P., Alimbekova L., Morozova (Osipova) E., 2006. Biostratigraphy of the Late Palaeolithic site of “Bajslan-Tash cave” (the Southern Urals) // Quaternary International. V. 149 (1). P. 115–121.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал