

Зоологический журнал, 2022, T. 101, № 11, стр. 1300-1304
Дальние перемещения малой лесной мыши (Sylvaemus uralensis, Muridae): скорость и дистанция, выявленные при групповом мечении
Е. Б. Григоркина a, *, Г. В. Оленев a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: grigorkina@ipae.uran.ru
Поступила в редакцию 08.02.2022
После доработки 04.03.2022
Принята к публикации 17.03.2022
- EDN: ZPGNMJ
- DOI: 10.31857/S0044513422110046
Аннотация
Представлены новые сведения о миграционных паттернах (расселении и переселении) малой лесной мыши (Sylvaemus uralensis (Pallas 1811)) за пределами домашнего участка обитания, полученные при изучении дальних перемещений мелких млекопитающих в Восточно-Уральском радиационном заповеднике (Челябинская обл., Южный Урал). Мы использовали метод группового мечения животного населения родамином В. Впервые получены данные о перемещении S. uralensis на расстояние 1500 м за 14 ч в темное время суток. Это позволяет судить о возможной скорости передвижения зверьков. Учитывая, что основная активность грызунов приходится на вечерние и утренние сумерки, фактическое время вполне могло быть значительно меньшим. Мигрантами оказались самцы-сеголетки разного функционального статуса: размножающийся (май 2019 г.) и неразмножающийся – перед уходом в зиму (сентябрь 2021 г.). Таким образом, оценены ночная дистанция, скорость передвижения S. uralensis, а также выявлен эпизод внешнего загрязнения биомаркером транзитного самца (октябрь 2021 г.), посетившего площадку мечения в период расселения. Наши результаты позволяют заключить, что дальние перемещения особей за пределами обычного домашнего участка являются нормой для S. uralensis.
Нерезидентная активность мелких млекопитающих за пределами участков обитания мало изучена, что обусловлено методическими и техническими сложностями их получения (Хляп, 1980; Щипанов, Купцов, 2004; Карасева и др., 2008; Kozakiewicz et al., 2007; Калинин, 2019). Неизвестно, как осуществляются переходы животных на большие расстояния, длительность и скорость их передвижения. Важность изучения дальних перемещений мышевидных грызунов в зонах локального загрязнения напрямую связана с вопросами адаптации популяций к антропогенной среде. Миграционные процессы имеют большое значение в распространении биологических эффектов техногенного воздействия на сопредельные территории (Theodarakis et al., 2001; Ryabokon, Goncharova, 2006; Ракитин и др., 2016; Bashlykova et al., 2019).
Цель работы – изучение миграционной подвижности мелких млекопитающих за пределами домашнего участка обитания с возможной оценкой скорости их перемещения.
МАТЕРИАЛ И МЕТОДЫ
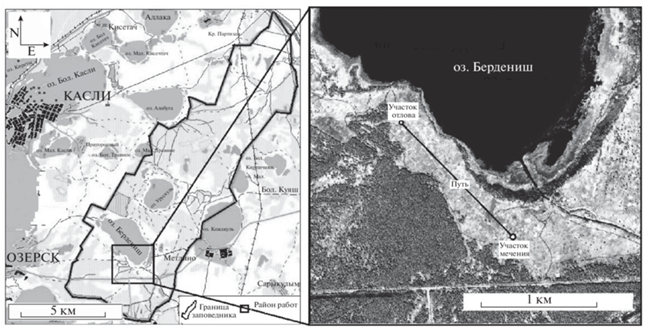
Исследования проведены в Восточно-Уральском радиационном заповеднике (Челябинская обл.) (Атлас …, 2013), который находится в лесостепной зоне Урала. Объект исследования – малая лесная мышь (Sylvaemus uralensis (Pallas 1811)) – абсолютный доминант в биотическом сообществе, доля которого в уловах существенно возросла после засухи 2010 г. (Оленев, Григоркина, 2016) и в совокупном материале сборов 2021 г. (6 отловочных сессий) составила 91%.
Для изучения миграций использован метод группового мечения родамином В (RB), который неизбирательно маркирует животное население (Lindsey, 1983; Weerakoon et al., 2013; Tolkachev, 2019; Григоркина, Оленев, 2021). В организме животных RB связывается со структурами, содержащими кератин (вибриссы, волосы, когти) (Fisher, 1999). Метка долго сохраняется и определяется по характерной желтой флуоресценции в зеленом свете при осмотре всей поверхности тела зверьков (Толкачев, Беспамятных, 2019).
Приманку с RB готовили на основе овсяных хлопьев по методике Толкачева и Беспамятных (2019), перед раскладыванием на экспериментальном участке опрыскивали нерафинированным подсолнечным маслом. На один эксперимент использовано 5 кг приманки с родамином (800 мг), которую равномерно распределяли в 17-00 на площади 1 га. Представлены материалы двух полевых экспериментов, проведенных весной 2019 и осенью 2021 гг., дизайн которых был сходен. В обоих случаях приманка была разложена на площадке мечения (55°46′20″ с.ш., 60°53′37″ в.д.) (рис. 1 ) в сухую малооблачную погоду. По данным логгеров, 22.05.2019 температура воздуха составила +15.0°C, 28.09.2021 г. – +8.0°C. Кусочки размещали преимущественно в естественных укрытиях для их большей сохранности и доступности для изучаемых животных. Этот способ оказался наиболее эффективным, поскольку зверьков с меткой отлавливали спустя 129 дней после мечения (Григоркина, Оленев, 2021; Григоркина и др., 2022). Контрольный отлов проведен на участке (55°44′56″ с.ш., 60°52′39″ в.д.), удаленном на расстояние 1500 м от площадки мечения (рис. 1 ). Использованы крючковые давилки с деревянным основанием, установленные на сутки линией (20 штук с интервалом 10 м). У добытых животных определяли вид, пол. Диагностика функционального статуса (зимовавшие-сеголетки) проведена по комплексу показателей: вес тела, состояние генеративной системы, индекс тимуса. Для возрастной диагностики учитывали одонтологические признаки (Колчева, 2009).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Весной 2019 г. (через 14 час после раскладывания приманки) на участке отлова (рис. 1 ) в выборке из 8 особей S. uralensis был обнаружен самец с четкой системной (фиксированной в тканях) меткой (табл. 1). При внешнем осмотре тушки и морфофизиологической обработке выявлены признаки поступление биомаркера с приманкой (рис. 2A, 2B). По функциональному статусу это размножающийся самец-сеголеток, родившийся в середине марта.
Таблица 1.
Характеристика самцов S. uralensis, преодолевших расстояние 1500 м в ночное время суток
| № | Дата мечения, отлова, время между ними |
Локализация родаминовой метки |
Способ получения метки | Вес тела (г) |
Вес тимуса, надпочечника, семенников (мг) |
Функциональное состояние, возраст |
|---|---|---|---|---|---|---|
| 1 | 22.05.19
23.05.19 14 ч |
Системная метка: шерсть на морде, передних лапах, в области уретры; в петлях кишечника, диафрагме |
Ел приманку | 23.0 | 10, 6–8, 226/222 |
Размножающийся сеголеток, 70 дней |
| 2 | 28.09.21 29.09.21 14 ч |
Несистемная метка: свечение частиц приманки с маркером на шерсти разных частей тела | Нюхал, коснулся приманки | 16.0 | 12, 2, 14/14 |
Неразмножающийся сеголеток, 45 дней |
Рис. 2.
Родаминовые метки у самцов-сеголеток S. uralensis, преодолевших расстояние 1500 м в ночное время суток: A, B – cистемная метка при поедании приманки с родамином В (желтое свечение) (А – на морде и лапе, B – в кишечнике); C – несистемная метка (внешнее загрязнение) при случайном соприкосновении животного с приманкой, содержащей краситель.
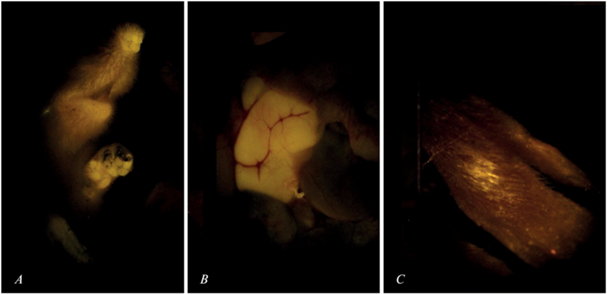
Аналогично, осенью 2021 г., на расстоянии 1500 м от площадки мечения (рис. 1 ) в улове из 7 особей S. uralensis оказался неразмножающийся самец-сеголеток, родившийся в середине августа, с несистемной (не фиксированной в тканях) меткой (табл. 1). Тип метки свидетельствует о внешнем загрязнении животного (рис. 2C) при случайном соприкосновении с приманкой или при ее обнюхивании. Признаков поедания корма с красителем не было обнаружено. Остальные мыши в обоих уловах были без меток.
Известно, что мышевидные грызуны и мелкие насекомоядные не ограничиваются передвижениями в пределах определенных границ, а время от времени расширяют знакомое пространство. При изучении подвижности – оседлости используют термин “нерезидентность”, предложенный Щипановым и Купцовым (2004) для обозначения перемещений животных за пределами участка обитания, т.к. эта активность включает экскурсии, переселения, собственно расселение и бродяжничество. Мы придерживаемся этой классификации, и обоих зверьков, преодолевших известное расстояние за определенное время, расцениваем как дальних нерезидентов (мигрантов). Ими оказались самцы-сеголетки альтернативных типов онтогенеза, которые различаются комплексом характеристик (скоростью роста, полового созревания, уровнем метаболизма, продолжительностью жизни, скоростью старения, радиорезистентностью и др.) и которые выполняют в популяции разные функции (Оленев, 2002). Зарегистрированные паттерны мы рассматриваем как переселение (breeding dispersal) (Kozakiewicz et al., 2007) и расселение (natal dispersal) – перемещение особи от места рождения до места возможного размножения (Howard, 1960). Причем осеннему зверьку присваиваем миграционный статус транзитного, прошедшего сквозь площадку мечения в ходе расселения. Максимальный суточный пробег S. uralensis по литературным источникам составляет 1.5–1.6 км (Хляп, 1980; Watts, 1970; Большаков, Баженов, 1988; Громов, 2008). Ранее эти данные позволяли оценить скорость перемещений лишь за сутки.
Таким образом, получены новые сведения о миграционных паттернах S. uralensis за пределами обычного домашнего участка обитания. Документально зафиксированные случаи дальних перемещений впервые позволяют предметно судить о скорости передвижения зверьков на расстояние 1500 м не более чем за 14 час. Учитывая, что основная активность грызунов приходится на вечерние и утренние сумерки, фактическое время вполне могло быть значительно меньшим. Используемый метод позволил не только оценить ночную дистанцию и скорость передвижения S. uralensis, но и выявить эпизод внешнего загрязнения биомаркером транзитного самца, случайно посетившего площадку мечения.
Полученные результаты исследования приводят к заключению, что дальние перемещения грызунов и землероек-бурозубок (6–9 км), которые регистрировались нами (Григоркина, Оленев, 2018, 2021) в ходе группового мечения тетрациклином и RB, не являются редкостью и могут рассматриваться как привычное явление. В зонах локального радиоактивного загрязнения следствиями дальних перемещений животных являются перенос радиационно-индуцированных эффектов (накопленного генетического груза) на сопредельные участки, изменение генетической структуры популяций и сохранение устойчивости надорганизменных систем (Lidicker, 1985; Большаков, Баженов, 1988; Theodarakis et al., 2001; Щипанов, 2002; Ryabokon, Goncharova, 2006; Ракитин и др., 2016; Bashlykova et al., 2019).
Список литературы
Атлас Восточно-Уральского и Карачаевского радиоактивных следов, включая прогноз до 2047 года, 2013. М.: ИГКЭ Росгидромета и РАН. 140 с.
Большаков В.Н., Баженов А.В., 1988. Радионуклидные методы мечения в популяционной экологии млекопитающих. М.: Наука. 157 с.
Григоркина Е.Б., Оленев Г.В., 2018. Миграции грызунов в зоне локального радиоактивного загрязнения на разных фазах динамики численности и их следствия // Известия РАН. Серия биологическая. № 1. С. 123–132.
Григоркина Е.Б., Оленев Г.В., 2021. Нерезидентная активность бурозубок (Sorex araneus, Eulipotyphla): массовое мечение // Экология. № 4. С. 316–320.
Григоркина Е.Б., Оленев Г.В., Толкачев О.В., 2022. Способ выявления точного места рождения мелких млекопитающих в исследованиях с применением группового мечения родамином // Экология. № 2. С. 153–158.
Громов В.С., 2008. Пространственно-этологическая структура популяций грызунов. М.: Товарищество научных изданий КМК. 581 с.
Калинин А.А., 2019. Последствия учетов мелких млекопитающих методом безвозвратного изъятия // Экология. № 3. С. 211–216.
Карасева Е.В., Телицына А.Ю., Жигальский О.А., 2008. Методы изучения грызунов в полевых условиях. М.: Изд-во ЛКИ. 416 с.
Колчева Н.Е., 2009. Стертость зубов как критерий возраста малой лесной мыши при анализе возрастной структуры популяций // Вестник ОГУ. Спецвыпуск. С. 77–80.
Оленев Г.В., 2002. Альтернативные типы онтогенеза цикломорфных грызунов и их роль в популяционной динамике (экологический анализ) // Экология. № 5. С. 341‒350.
Оленев Г.В. Григоркина Е.Б., 2016. Эволюционно-экологический анализ стратегий адаптации популяций грызунов в экстремальных условиях // Экология. № 5. С. 375–381.
Ракитин С.Б., Григоркина Е.Б., Оленев Г.В., 2016. Анализ микросателлитной ДНК у грызунов из зоны Восточно-Уральского радиоактивного следа и сопредельных территорий // Генетика. Т. 52. № 4. С. 453–460.
Толкачёв О.В., Беспамятных Е.Н., 2019. Новый метод детекции родаминовой метки и возможности его применения в зоологических исследованиях // Журнал СФУ. Биология. Т. 12. № 4. С. 352–365.
Щипанов Н.А., 2002. Функциональная организация популяций: возможный подход к изучению популяционной устойчивости. Прикладные аспекты (на примере мелких млекопитающих) // Зоологический журнал. Т. 81. № 9. С. 1048–1077.
Щипанов Н.А., Купцов А.В., 2004. Нерезидентность у мелких млекопитающих и ее роль в функционировании популяции // Успехи современной биологии. Вып. 124. № 1. С. 28–43.
Хляп Л.А., 1980. Основные проблемы изучения связей мелких млекопитающих с территорией // Фауна и экология грызунов. Вып. 14. С. 44–62.
Bashlykova L., Rascosha O., Starobor N., 2019. Induction of abnormal sperm heads in small mammals under chronic ionizing radiation // Turkish Journal of Zoology. V. 43. P. 540–544.
Fisher P., 1999. Review of using Rhodamine B as a marker for wildlife studies // Wildl. Soc. Bull. V. 27. P. 318–329.
Howard W.E., 1960. Innate and environmental dispersal of individual vertebrates // American Midland Naturalist. V. 63. P. 152–161.
Kozakiewicz M., Cho£uj A., Kozakiewicz A., 2007. Long-distance movements of individuals in a free-living bank vole population: an important element of male breeding strategy // Acta Theriologica. V. 52 (4). P. 339–348.
Lidicker W.Z. Jr., 1985. Dispersal. The American Society of Mammalogists. Special publication. V. 8. P. 420–454.
Lindsey G.D., 1983. Rhodamine B: a systemic fluorescent marker for studying mountain beavers (Aplodontia rufa) and other animals // Northwest Science. V. 57 (1). P. 16–21.
Ryabokon N.I., Goncharova R.I., 2006. Transgenerational accumulation of radiation damage in small mammals chronically exposed to Chernobyl fallout // Radiat. Environ. Biophys. V. 45. P. 167–177.
Theodarakis C.W., Bickham J.W., Lamb T. Medica P.A., Lyne T.B., 2001. Integration of genotoxicity and population genetic analyses in kangaroo rats (Dipodomys merriami) exposed to radionuclide contamination at the Nevada Test Site, USA // Environ. Toxicol. and Chem. V. 20. № 2. P. 317–326.
Tolkachev O., 2019. A new baiting scheme and simple method of rhodamine B detection could improve biomarking of small mammals // European Journal of Wildlife Research. V. 65. № 10. https://doi.org/10.1007/s10344-018-1243-5
Watts C.H.S., 1970. Long distance movement of bank voles and wood mice // J. Zool., Lond. V. 161. P. 247–256.
Weerakoon M.K., Price C.J., Banks P.B., 2013. Hair type, intake, and detection method influence Rhodamine B detectability // J. Wildl. Manag. V. 77. P. 306–312.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал