

Зоологический журнал, 2022, T. 101, № 12, стр. 1337-1349
Восточно-азиатский вселенец Thermocyclops taihokuensis (Harada 1931) и аборигенный Thermocyclops oithonoides (Sars 1863) (Crustacea, Cyclopoida): сравнительный анализ морфологии двух родственных видов из водоемов европейской части России
В. И. Лазарева a, *, С. М. Жданова a, **
a Институт биологии внутренних вод имени И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: lazareva_v57@mail.ru
** E-mail: zhdanova83@gmail.com
Поступила в редакцию 30.04.2022
После доработки 13.05.2022
Принята к публикации 02.06.2022
- EDN: HRHXQC
- DOI: 10.31857/S0044513422120078
Аннотация
Детали строения тела половозрелых копепод (самки и самцы) двух морфологически близких видов рода Thermocyclops – вселенца T. taihokuensis из Шатского водохранилища (Тульская обл.) и нативного T. oithonoides из оз. Плещеево (Ярославская обл.) – исследованы с целью уточнения известных морфологических признаков и выявления новых, позволяющих надежно различить эти виды при совместном обитании. Строение копепод анализировали с использованием программ визуализации и измерения наблюдаемых в микроскопе объектов AxioVision Rel. 4.8 и ScopePhoto 3.0. Рисунки тонких структур выполнены с фотографий. Показано, что различение данных двух видов основывается преимущественно на качественных характеристиках (форма элемента, его расположение и т.п.). Не выявлено таксономически значимой разницы в измеряемых параметрах элементов четвертой, пятой и шестой (у самцов) пар ног, каудальных ветвей и их щетинок, обычно используемых для идентификации видов рода. Установлено, что для идентификации самок этих видов наиболее важны форма семяприемника, апикальных щетинок каудальных ветвей, апикальных шипов эндоподита четвертой пары ног (Р4) и положение боковой щетинки на каудальных ветвях. Выявлены межвидовые различия в вооружении выростов интеркоксальной пластинки, орнаментации коксоподита и интеркоксальной пластинки Р4, а также вооружении 14-го членика антеннул самок. Предложено использовать эти характеристики как дополнительные для идентификации видов. Показано, что самцы T. taihokuensis существенно крупнее таковых T. oithonoides, они различаются по положению боковой щетинки каудальных ветвей, вооружению Р4 и длине средних апикальных щетинок. Предложен ключ для определения шести распространенных в европейской части России видов рода Thermocyclops.
Основной ареал Thermocyclops taihokuensis (Harada 1931) (syn. T. asiatiсus (Kiefer 1932) охватывает Восточную и Центральную Азию (Рылов, 1948; Mirabdullayev, Kuzmetov, 1997; Mirabdullayev et al., 1997, 2003; Guo, 1999; Monchenko, 2008; Dela Paz et al., 2016). Вид обитает также в водоемах Казахстана и Узбекистана (Степаньянц и др., 2015; Калымбетова, 2017; Mirabdullayev, Kuzmetov, 1997; Mirabdullayev et al., 2003; Monchenko, 2008). В 2000-х годах он обнаружен в Северо-Восточном и Среднем Каспии (Шарапова, 2014; Степаньянц и др., 2015; Monchenko, 2008). До 2010 г. его не находили севернее Арала (47° с.ш.) и западнее устья р. Урал (52° в.д.) (Mirabdullayev et al., 1997; Степаньянц и др., 2015; Monchenko, 2008).
В начале 2010-х годов T. taihokuensis проник в бассейны рек Волги и Дона. Его регистрировали в Волго-Ахтубинской пойме (Нечаев, 2016), а с 2012 г. – в Цимлянском водохранилище (р. Дон) (Вехов и др., 2014). Во второй половине 2010-х годов (2016–2019 гг.) этот вид заселил уже четыре района бассейна р. Волги, удаленных друг от друга на расстояние 270–1000 км (Жихарев и др., 2019; Lazareva et al., 2022). Вселенец обитает на незарегулированном участке Нижней Волги от с. Черный Яр до г. Астрахань (Нижняя Волга), в озеровидной части Чебоксарского водохранилища, на Волжском и Волго-Камском плесах Куйбышевского водохранилища (Средняя Волга) и в верховье бассейна р. Ока (Шатское водохранилище). В 2018–2019 гг. впервые выявлено местообитание копеподы T. taihokuensis в р. Дон выше и ниже Цимлянского водохранилища, а также в водоемах Волго-Донского канала (Лазарева, Сабитова, 2021; Лазарева, 2022). На сегодня северная (56° с.ш., 46° в.д.) граница распространения T. taihokuensis расположена в устье р. Сура (Средняя Волга), западная (54° с.ш., 38° в.д.) – в Шатском водохранилище.
В европейской части России ареал T. taihokuensis перекрывается с таковым обычного для таежной зоны вида этого рода T. oithonoides (Sars 1863). Еще Рылов (1948) отмечал, что эти два вида чрезвычайно сходны по строению ног и вооружению каудальных ветвей. Заметим, что в ряде определителей фауны европейской части России отсутствует описание T. taihokuensis (Монченко, 1974; Определитель …, 2010). А основные признаки T. oithonoides в определительном ключе видов рода Thermocyclops фактически повсеместно сведены как раз к особенностям оснащения фуркальных ветвей (Монченко, 1974; Определитель …, 2010). Лишь в определителе рыб и беспозвоночных Каспийского моря (Степаньянц и др., 2015) идентификационный ключ для T. taihokuensis включает описание семяприемника и особенности вооружения дистального членика антеннулы самки.
Целью работы было выявление признаков, позволяющих надежно отличить вселенца T. taihokuensis от морфологически близкого аборигенного вида T. oithonoides при их совместном обитании.
МАТЕРИАЛ И МЕТОДЫ
Для анализа строения тела использовали половозрелых копепод (самки и самцы). Особи T. taihokuensis (12 самок и 8 самцов) отловлены в 2019 г. в Любовском пруду-охладителе Новомосковской ГРЭС на Шатском водохранилище (Тульская обл.), T. oithonoides (14 самок и 10 самцов) – в 2014 г. в оз. Плещеево (Ярославская обл.). Рачков в полевых условиях отлавливали сетью Джеди с диаметром входного отверстия 12 см и ситом с ячеей 74 мкм, а также мерным ведром с последующей концентрацией через сито с ячеей 74 мкм, фильтровали 100–200 л воды.
Сборы фиксировали 4% формалином и просматривали в лаборатории под стереомикроскопом StereoDiscovery-12 (Carl Zeiss, Jena). Таксономическую идентификацию копепод рода Thermocyclops проводили с использованием определительных ключей и описаний видов, приведенных в работах (Рылов, 1948; Монченко, 1974; Степаньянц и др., 2015; Guo, 1999; Mirabdullayev et al., 2003; Monchenko, 2008).
Фотографии выполнены камерой AxioCam MRс5 (микроскоп StereoDiscovery V.12) при увеличении 60×, NikonDS-Fi1 (микроскоп Nikon Eclipse 80i) при увеличении 20–100× и цифровым видеоокуляром Hayear HY-500B (микроскоп Микромед 3 U3) при увеличении 40–100×. Длину тела рачков измеряли с помощью окулярного микрометра 50/10 мм Stemi при увеличении 25–50×. Детали строения копепод анализировали с использованием программ визуализации и измерения наблюдаемых в микроскопе объектов AxioVision Rel. 4.8 и ScopePhoto 3.0. Рисунки тонких структур выполнены методом копирования их фотографий с уточнением деталей по снимкам с различным увеличением (до 100×).
В работе проанализированы 13 относительных меристических признаков (табл. 1). Абсолютные значения показателей не обсуждали, поскольку они сильно варьируют в зависимости от размера особей. Всего измерено 20 взрослых особей T. taihokuensis и 24 особи T. oithonoides. Анализировали также ряд качественных морфологических характеристик, в частности, форму боковых ветвей семяприемника, внутреннего шипа дистального членика четвертой пары ног (Р4) и апикальных щетинок каудальных ветвей; положение боковой щетинки на каудальных ветвях, орнаментацию коксоподита и интеркоксальной пластинки Р4, вооружение выступов интеркоксальной пластинки Р4, вооружение базиподита первой пары ног (Р1) и дистальных члеников (14 и 17) антеннулы самки.
Таблица 1.
Меристические параметры, использованные при идентификации видов рода Thermocyclops
| № | Описание параметра | Аббревиатура параметра |
|---|---|---|
| 1 | Отношение длины дистального членика эндоподита Р4 к его ширине | Long. end. P4/lat. end. P4 |
| 2 | Соотношение длины внешнего и внутреннего дистальных шипов эндоподита Р4 | Long. sp. ext. end. P4/long. sp. int. end. P4, % |
| 3 | Соотношение длины шипа и дистальной щетинки Р5 | Long. sp. P5/long. set. maj. Р5 |
| 4 | Отношение длины щетинки проксимального членика Р5 к длине щетинки дистального членика Р5 | Long. set. min. Р5/long. set. maj. Р5, % |
| 5 | Отношение длины шипа Р6 самца к длине средней щетинки Р6 | Long. sp. P6/long. set. med. P6 |
| 6 | Отношение длины шипа Р6 самца к длине крайней щетинки Р6 | Long. sp. P6/long. set. ext. P6 |
| 7 | Отношение длины фуркальных ветвей к их ширине (фуркальный индекс) | Long. furc./lat. furc. |
| 8 | Отношение длины дорзальной щетинки к длине внешней апикальной щетинки фуркальных ветвей | Long. set. dors./long. set. apic. ext. |
| 9 | Отношение длины дорзальной щетинки к длине внутренней апикальной щетинки фуркальных ветвей | Long. set. dors./long. set. apic. int. |
| 10 | Отношение длины внутренней апикальной щетинки к длине внешней апикальной щетинки фуркальных ветвей | Long. set. apic. int./long. set. apic. ext. |
| 11 | Отношение длины внутренней из средних апикальных щетинок к длине фуркальных ветвей | Long. set. apic. med. int./long. furc. |
| 12 | Отношение длины внешней средней апикальной щетинки к длине внутренней апикальной щетинки фуркальных ветвей | Long. set. apic. med. ext./long. set. apic. int. |
| 13 | Отношение расстояния от нижнего края фуркальных ветвей до боковой щетинки фурки к длине фуркальных ветвей | Sp. furc. II/long. furc. |
РЕЗУЛЬТАТЫ
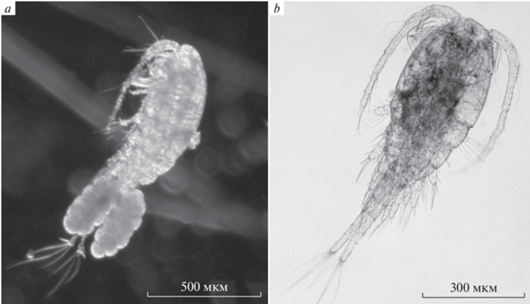
Вселенец T. taihokuensis в изученном водоеме характеризовался сравнительно крупными размерами тела, длина тела самки 0.95 ± 0.01, самца 0.86 ± 0.03 мм. От других видов рода самки вселенца даже при небольшом увеличении (20–25×) хорошо отличались широким веером волнистых апикальных щетинок каудальных ветвей и сдвинутой на спинную сторону их боковой щетинкой (рис. 1a). Аборигенный T. oithonoides значительно мельче – самка 0.75 ± 0.08, самец 0.62 ± 0.03 мм (т.е. почти в 1.5 раза мельче). Вооружение каудальных ветвей T. oithonoides типично для рода и представлено прямыми щетинками (рис. 1b).
Промеры таксономически значимых элементов четвертой, пятой и шестой (у самцов) пар ног, фуркальных ветвей и их щетинок показали, что большинство параметров вселенца и аборигенного вида различаются не значительно. Заметные различия выявлены по признакам № 3 и 5 у самцов, № 13 у самок и № 4 у обоих полов (табл. 2). Однако в большинстве случаев это можно выявить только при сравнении средних значений, измеренных не менее чем у 10 особей.
Таблица 2.
Морфометрические параметры Thermocyclops taihokuensis и T. oithonoides
| № | Параметр | T. taihokuensis | T. oithonoides | ||
|---|---|---|---|---|---|
| самка | самец | самка | самец | ||
| 1 | Long. end. P4/lat. end. P4 | 3.3 ± 0.1 | 3.6 ± 0.2 | 3.0 ± 0.1 | 3.0 ± 0.1 |
| 2 | Long. sp. ext. end. P4/long. sp. int. end. P4, % | 31 ± 1 | 36 ± 1 | 28 ± 1 | 32 ± 1 |
| 3 | Long. sp. P5/long. set. maj. Р5 | 1.07 ± 0.05 | 1.09 ± 0.04 | 1.08 ± 0.05 | 1.47 ± 0.05 |
| 4 | Long. set. min. Р5/long. set. maj. Р5, % | 62 ± 3 | 84 ± 19 | 84 ± 4 | 113 ± 6 |
| 5 | Long. sp. P6/long. set. med. P6 | – | 1.8 ± 0.12 | – | 1.3 ± 0.04 |
| 6 | Long. sp. P6/long. set. ext. P6, % | – | 48 ± 2 | – | 41 ± 1 |
| 7 | Long. furc./lat. furc. | 3.0 ± 0.1 | 2.8 ± 0.1 | 3.1 ± 0.1 | 3.0 ± 0.1 |
| 8 | Long. set. dors./long. set. apic. ext. | 2.0 ± 0.1 | 2.8 ± 0.2 | 2.6 ± 0.1 | 2.8 ± 0.2 |
| 9 | Long. set. dors./long. set. apic. int., % | 62 ± 2 | 65 ± 2 | 70 ± 2 | 71 ± 3 |
| 10 | Long. set. apic. int./long. set. apic. ext. | 3.2 ± 0.1 | 4.4 ± 0.2 | 3.8 ± 0.2 | 4.0 ± 0.2 |
| 11 | Long. set. apic. med. int./long. furc. | 3.7 ± 0.1 | 4.2 ± 0.2 | 3.5 ± 0.1 | 3.4 ± 0.1 |
| 12 | Long. set. apic. med. ext./long. set. apic. int. | 1.16 ± 0.02 | 1.11 ± 0.02 | 1.08 ± 0.04 | 1.08 ± 0.02 |
| 13 | Sp. furc. II/long. furc., % | 37 ± 0.3 | 43 ± 0.3 | 48 ± 1 | 39 ± 7 |
Так, у самцов T. taihokuensis шип и щетинка дистального членика пятой пары ног (Р5) почти равны, тогда как у T. oithonoides шип длиннее щетинки в среднем в 1.5 раза. У обоих полов T. taihokuensis щетинка проксимального членика Р5 относительно короткая, она составляет ~60% длины дистальной щетинки у самок и почти 85% у самцов. У T. oithonoides щетинка проксимального членика Р5 длинная, она достигает у самок ~85% дистальной, а у самцов ее длина даже превышает (113%) таковую щетинки дистального членика Р5. Длина шипа рудиментарной шестой пары ног (Р6) вселенца почти вдвое превышает таковую средней щетинки и достигает ~50% крайней щетинки. Тогда как у нативного вида шип Р6 только на ~30% длиннее средней щетинки и составляет ~40% крайней. Различия между видами по этим признакам невелики и не позволяют четко отличить вселенца от нативного вида.
Расстояние от нижнего края каудальных ветвей до боковой щетинки фурки самок и самцов T. taihokuensis составляет ~40% длины каудальных ветвей, также у самцов T. oithonoides. Однако у самок нативного вида боковая щетинка расположена близко к середине каудальных ветвей и почти перпендикулярно их поверхности (рис. 2d). Заметим, что у вселенца боковая щетинка сильно сдвинута на спинную сторону ветвей, в дорсо-вентральном положении абдомена она не заметна на фотографиях (рис. 1a, 2c).
Рис. 2.
Важные для идентификации качественные признаки самок Thermocyclops taihokuensis (a, c) и T. oithonoides (b, d): a, b – семяприемник; c, d – форма апикальных щетинок и положение боковой щетинки фуркальных ветвей.
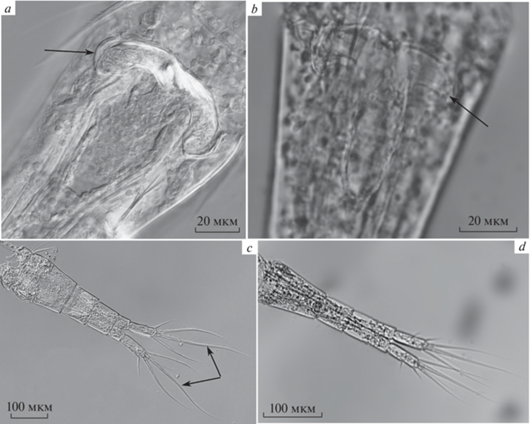
Важными для различения вселенца и нативного вида являются также ряд качественных морфологических признаков. В первую очередь это форма семяприемника и апикальных щетинок каудальных ветвей самки. Боковые ветви семяприемника у T. taihokuensis длинные, их края сильно загнуты вниз и назад (рис. 2a). У T. oithonoides боковые выросты семяприемника слабо изогнуты или прямые, они расположены перпендикулярно продольной оси генитального сегмента (рис. 2b). Внутренняя средняя апикальная щетинка каудальных ветвей самок вселенца S-образно изогнута (рис. 2c), у самок нативного вида все апикальные щетинки прямые (рис. 2d). Дополнительный отличительный признак самцов T. taihokuensis – очень длинные и тонкие средние апикальные щетинки каудальных ветвей, которые на концах сильно согнуты на брюшную сторону.
У представленных двух видов хорошо различается строение Р4. Внутренний шип дистального членика эндоподита Р4 вселенца прямой и толстый, он вооружен крепкими зубцами, редко и симметрично расположенными на обоих его краях (рис. 3a). У нативного вида этот шип грациозно изогнут в основании и сильно отклонен вовнутрь от продольной оси членика (рис. 3b). Выросты интеркоксальной пластинки Р4 T. taihokuensis несут 5–6 широких в основании треугольных шипов, неровно расположенных по всей нижней поверхности выростов (рис. 3c, 4a). У T. oithonoides на этих выростах находятся 6–7 тонких, сравнительно длинных и когтеобразно изогнутых шипов (рис. 3d, 4b), хорошо различимых только при большом (100×) увеличении.
Рис. 3.
Вооружение ног четвертой пары Thermocyclops taihokuensis (a, c, e) и T. oithonoides (b, d, f): a, b – форма внутреннего шипа дистального членика Р4; c, d – вооружение выступов интеркоксальной пластинки Р4; e, f – орнаментация щетинками поверхности интеркоксальной пластинки Р4.
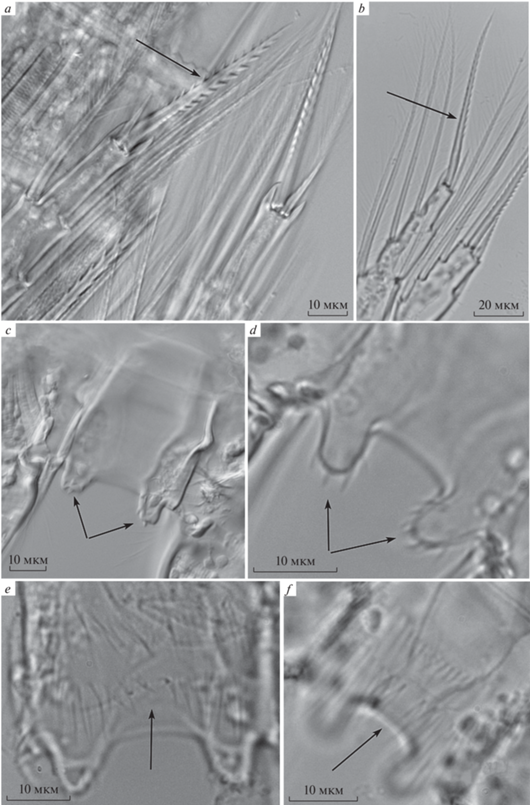
Рис. 4.
Орнаментация коксоподита и интеркоксальной пластинки Р4 Thermocyclops taihokuensis (a) и T. oithonoides (b).
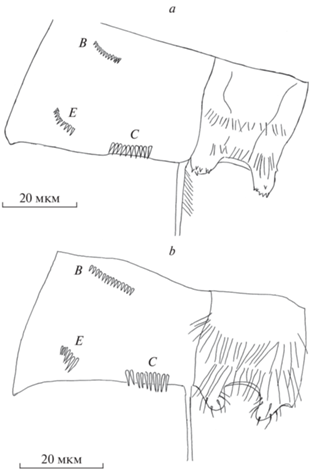
Интеркоксальная пластинка обоих видов покрыта двумя рядами густо расположенных волосков. У вселенца они заметно короче и толще (рис. 3e), чем у нативного вида (рис. 3f). Кроме того, у T. oithonoides в нижнем ряду волоски более длинные, они достигают нижнего края выступов пластинки. Орнаментация коксоподита Р4 представлена тремя группами шипиков, по аналогии с обозначениями таких групп у видов рода Cyclops (Einsle, 1993) их можно обозначить как В, C и Е (рис. 4). Количество шипиков в группах B и C у обоих видов одинаково и составляет 12–14 и 9–11 соответственно. Но в группе В нативного вида шипики удлиненные они частично выходят за край коксоподита (рис. 4b). В группе Е на коксоподите Р4 T. taihokuensis насчитывается 10–12 шипиков, постепенно уменьшающихся в направлении внешнего края ног (рис. 4a). У T. oithonoides шипики в группе Е сильно различаются по размеру, а их количество (6–7) почти вдвое меньше, чем у вселенца (рис. 4b).
Обнаружены различия в вооружении дистальных члеников антеннул самок вселенца и аборигенного вида. Так, у T. taihokuensis на апикальном крае дистального членика антеннулы, две из 5 щетинок очень длинные, они как минимум на 30–50% длиннее остальных (рис. 5a). На 14-м членике антеннулы присутствует еще более длинная щетинка, достигающая конца дистального членика или немного выходящая за него. У T. oithonoides на конце дистального членика антеннулы также расположены две удлиненные щетинки, однако они короче, чем у вселенца. Наиболее длинная из пяти щетинок не более чем на 30% длиннее трех коротких (рис. 5b). Напротив, щетинка на 14-м членике антеннулы заметно длиннее по сравнению с таковой вселенца, она выходит далеко за внешний край дистального членика и часто даже перекрывает короткие щетинки на его конце.
Рис. 5.
Вооружение дистальных члеников антеннулы самки и базиподита Р1 Thermocyclops taihokuensis (a, c) и T. oithonoides (b, d). А1 – стрелками помечены длинные щетинки на дистальном и 14-м члениках антеннулы самок, Р1 – стрелками помечены волоски и щетинка на внутреннем крае базиподита Р1.

Базиподит первой пары ног (Р1) обоих видов с внутренней стороны покрыт тонкими волосками. Выявлены различия в длине щетинки, расположенной на внутренней лопасти базиподита Р1 (рис. 5). В частности, у T. taihokuensis внутренний край базиподита Р1 несет щетинку, достигающую основания дистального членика эндоподита (enp3) Р1 (рис. 5c). Тогда как у аборигенного T. oithonoides эта щетинка очень длинная, она достигает середины дистального членика эндоподита (enp3) (рис. 5d).
Итак, по нашим наблюдениям, для идентификации самок T. taihokuensis и их отличия от T. oithonoides наиболее важны форма семяприемника, апикальных щетинок фурки, апикальных шипов эндоподита Р4, а также положение боковой щетинки фуркальных ветвей и вооружение выростов интеркоксальной пластинки Р4. Дополнительными признаками могут быть орнаментация коксоподита и интеркоксальной пластинки Р4, а также вооружение дистальных члеников антеннул самок. Самцы вселенца крупные и грациозные, длина их тела обычно больше, чем даже у самок T. oithonoides. От нативного вида самцов вселенца отличал общий габитус тела, положение боковой щетинки фуркальных ветвей и очень длинные, согнутые на брюшную строну средние апикальные щетинки фурки.
Определительная таблица видов, обитающих в европейской части России
1(4) Семяприемник самки молотоообразный, его боковые выросты слабо изогнуты или прямые, расположены перпендикулярно продольной оси генитального сегмента
2(3) Внутренний апикальный шип дистального членика эндоподита Р4 отогнут вовнутрь, его длина превышает таковую внешнего в 3–3.5 раза. Боковая щетинка каудальных ветвей самки расположена строго латерально и близко к середине их наружного края ……………………… T. oithonoides (Sars 1863)
3(2) Внутренний апикальный шип дистального членика эндоподита Р4 сравнительно короткий, его длина превышает таковую внешнего в 2–3 раза. Боковая щетинка смещена к задней трети каудальных ветвей ………… T. crassus (Fischer 1853)
4(1) Боковые ветви семяприемника длинные, их края сильно изогнуты вниз и назад Внутренний шип дистального членика эндоподита Р4 прямой и толстый, вооружен крепкими зубцами, редко и симметрично расположенными на обоих его краях. Боковая щетинка сильно сдвинута на спинную сторону каудальных ветвей. Внутренняя средняя апикальная щетинка каудальных ветвей самки S-образно изогнута …………… T. taihokuensis (Harada 1931)
5(4) Внутренний шип дистального членика эндоподита Р4 слегка изогнут и асимметрично вооружен шипиками на внешней поверхности и тонкими сетулами на внутренней. Внутренняя средняя апикальная щетинка каудальных ветвей самки прямая ………………………………… T. vermifer Lindberg 1935
6(4) Боковые ветви семяприемника удлинненные, их края немного опущены вниз и загнуты назад
7(8) Внутренний апикальный шип дистального членика эндоподита Р4 сравнительно короткий, приблизительно вдвое длиннее внешнего, слегка изогнут и с обеих сторон вооружен крепкими зубчиками. Боковая щетинка каудальных ветвей смещена к их дистальному концу. Дорсальная щетинка каудальных ветвей короткая, вдвое короче внутренней крайней апикальной щетинки ………………………………………… T. rylovi (Smirnov 1928)
8(7) Внутренний апикальный шип дистального членика эндоподита Р4 прямой и очень короткий, на 10–20% короче внешнего. Выросты интеркоксальной пластинки Р4 слабовыраженные, почти не выходят за край пластинки. Боковая щетинка расположена в задней трети каудальных ветвей ….……....................T. dybowskii (Lande 1890)
ОБСУЖДЕНИЕ
Для водоемов России указывают шесть видов рода Thermocyclops: T. oithonoides, T. crassus, T. dybowskii (Lande 1890), T. vermifer Lindberg 1935, T. rylovi (Smirnov 1928) и T. taihokuensis (Рылов, 1948; Степаньянц и др., 2015). В европейской части России широко распространены первые два вида. Данные последних лет о распространении T. taihokuensis указывают на тот факт, что в Средней и Нижней Волге, а также на р. Дон от Цимлянского водохранилища до дельты этого недавнего вселенца также можно отнести к обычным и локально даже многочисленным видам (Лазарева, Сабитова, 2021; Lazareva et al., 2022).
Последнее переописание T. taihokuensis сделано Монченко (Monchenko, 2008) из солоноватых водоемов восточного побережья Среднего Каспия (Туркменистан), в которых этот вид представлен очень мелкими особями (в среднем <0.7 мм). Длина тела половозрелых самок T. taihokuensis в Шатском водохранилище и водохранилищах р. Волги существенно больше (в среднем 1.0 ± ± 0.02 мм) (Lazareva et al., 2022). Это близко к размерам рачков из Казахстана (0.99 мм), Монголии (0.96 мм), Узбекистана и Китая (0.94 мм), пойменных водоемов р. Амур (0.92 мм) (Рылов, 1948; Mirabdullayev, Kuzmetov, 1997; Mirabdullayev et al., 1997; Guo, 1999). В типовом местообитании на Тайване длина самок варьирует в пределах 0.80–0.88 мм (Mirabdullayev, Kuzmetov, 1997). Особи из бассейна р. Волги несколько крупнее тайваньских и значительно (в 1.5 раза) крупнее рачков из Туркмении. Наиболее крупные особи вселенца (в среднем >1 мм) обнаружены в Чебоксарском и Куйбышевском водохранилищах – самых северных его местообитаниях (Lazareva et al., 2022). Однако такие же крупные (до 1.1 мм) представители T. taihokuensis отмечены в водоемах Филиппин (Dela Paz et al., 2016).
Описание и рисунки обоих видов, в том числе форма семяприемника самок, даны в сводках (Рылов, 1948; Степаньянц и др., 2015; Mirabdullayev et al., 2003). Подробное описание с рисунками T. taihokuensis приведены также в статьях (Mirabdullayev, Kuzmetov, 1997; Mirabdullayev et al., 1997; Guo, 1999; Monchenko, 2008; Dela Paz et al., 2016). Сопоставление литературных характеристик T. oithonoides и T. taihokuensis показывает, что ряд признаков, особенно мелкие детали вооружения ног и каудальных ветвей, авторы характеризуют по-разному. Часто таксономические признаки, указанные для одного вида, отсутствуют в описании другого. Детали строения, различимые только на большом увеличении (60–100×), оказались не точны в сравнительно старых описаниях видов (Рылов, 1948; Монченко, 1974; Mirabdullayev, Kuzmetov, 1997; Mirabdullayev et al., 1997; Guo, 1999). Это связано, вероятно, с еще недавним отсутствием оптики высокого разрешения.
Соотношение длины щетинок каудальных ветвей, которое часто используют для идентификации видов рода Thermocyclops, у обоих исследованных видов близки или заметно перекрываются (Монченко, 1974; Mirabdullayev et al., 2003; Monchenko, 2008; Степаньянц и др., 2015). То же выявлено в настоящей работе (табл. 2). К тому же у каждого из видов отмечены региональные вариации этих характеристик, что еще более усложняет их сопоставление. Наши индексы каудальных щетинок аборигенного T. oithonoides из оз. Плещеево (табл. 2) почти идентичны указанным в работе Монченко (1974) для особей из бассейна р. Днепр. Напротив, у T. taihokuensis из Шатского водохранилища они заметно отличаются в большую или меньшую сторону от указанных для популяций Дальнего Востока (Рылов, 1948) и Центральной Азии (Mirabdullayev et al., 2003; Monchenko, 2008). Вероятно, миграция вселенца к северо-западу от исходных местообитаний сопровождается изменением строения его тела.
В качестве важного признака T. taihokuensis в некоторых работах рассматривают длинную щетинку на конце дистального членика антеннул самки, которая вдвое длиннее прочих (Степаньянц и др., 2015; Mirabdullayev et al., 2003). Мы выяснили, что этот признак не пригоден для различения вселенца и аборигенного вида. Дистальный членик антеннулы обоих видов несет не одну, а две длинные щетинки. У T. taihokuensis одна из них длиннее трех более коротких на 30–50%. У T. oithonoides самая длинная из них тоже на ~30% длиннее трех коротких щетинок. Более существенные различия наблюдаются по параметрам мощной и длинной щетинки 14-го членика антеннулы (рис. 5a–5b). У T. taihokuensis эта щетинка достигает конца антеннулы или немного выходит за него, а у T. oithonoides – далеко выходит за конец антеннулы и перекрывает часть коротких щетинок на нем.
Во многих определительных ключах видов рода Thermocyclops России в качестве основного отличительного признака T. oithonoides указано расположение боковой щетинки, близкое к середине наружного края каудальных ветвей (Рылов, 1948; Определитель …, 2010; Степаньянц и др., 2015). По нашим данным это справедливо только для самок, у самцов боковая щетинка сдвинута к задней трети фурки (табл. 2) – тоже отмечено для этого вида в сводке (Монченко, 1974). Для разграничения T. taihokuensis и T. oithonoides важным является тот факт, что боковая щетинка у обоих полов вселенца сильно сдвинута на спинную сторону каудальных ветвей. Тогда как у аборигенного T. oithonoides она обычно прикреплена к их боковой поверхности почти перпендикулярно, что делает эту щетинку хорошо заметной.
Форма семяприемника самки – по-видимому, один из самых важных современных критериев идентификации представителей рода Thermocyclops мировой фауны (Mirabdullayev et al., 2003; Chaicharoen et al., 2011; Dela Paz et al., 2016). Семяприемники вселенца и аборигенного вида хорошо различаются (Рылов, 1948; Mirabdullayev et al., 2003; Степаньянц и др., 2015). Описание этих структур для обоих видов на русском языке и их рисунки приведены в сводках (Рылов, 1948; Степаньянц и др., 2015), а в одной из наших работ даны фотографии семяприемника T. taihokuensis (Lazareva et al., 2022). К сожалению, в определительном ключе рода Thermocyclops в книге (Определитель …, 2010), широко используемом российскими гидробиологами, этот признак не применяется. В сопроводительных иллюстрациях даны лишь рисунки семяприемников самок разных видов без объяснения, на что нужно обратить внимание.
Особенности вооружения плавательных ног четвертой пары традиционно используют для идентификации представителей рода Thermocyclops во всех таксономических работах (Рылов, 1948; Монченко, 1974; Определитель …, 2010; Степаньянц и др., 2015; Mirabdullayev, Kuzmetov, 1997; Mirabdullayev et al., 1997, 2003; Guo, 1999; Monchenko, 2008; Chaicharoen et al., 2011; Dela Paz et al., 2016). Соотношение длины внутреннего (медиального) и внешнего (латерального) апикальных шипов дистального членика эндоподита (enp3) P4 T. taihokuensis сильно варьирует. В разных популяциях показатель изменяется от 2.6–3.9 (Mirabdullayev et al., 2003; Monchenko, 2008) до 3.0–4.5 (Mirabdullayev et al., 1997; Guo, 1999; Dela Paz et al., 2016). Наши данные (2.8–3.2) близки к указанным в работах (Mirabdullayev et al., 2003; Monchenko, 2008). Также вариабельно соотношение длины апикальных шипов эндоподита Р4 (3–5) у нативного T. oithonoides (Монченко, 1974; Степаньянц и др., 2015; Mirabdullayev et al., 2003), в популяции из оз. Плещеево 3.1–3.6 (табл. 2). Эта характеристика не пригодна для разграничения указанных двух видов, поскольку вариации соотношения длины шипов у них перекрываются. В данном случае следует обратить внимание на форму внутреннего шипа каждого из видов. У T. taihokuensis он мощный, прямой и сильно зазубренный, а у T. oithonoides – более тонкий и в основании изгибающийся вовнутрь от вертикальной оси эндоподита (рис. 3a–3b).
Существуют различные мнения по поводу орнаментации коксоподита Р4, а также количества и формы шипов на выростах интеркоксальной пластинки обоих видов. Так, для T. taihokuensis указывают наличие маленьких зубчиков или мелких шипиков на высоких полушаровидных выростах интеркоксальной пластинки Р4 (Mirabdullayev et al., 1997; Monchenko, 2008; Dela Paz et al., 2016), также крепких зубчиков на хорошо развитых выростах (Степаньянц и др., 2015). Число зубчиков у разных авторов варьирует от 3–6 (Mirabdullayev et al., 1997; Guo, 1999; Mirabdullayev et al., 2003), до 5–7 (Определитель …, 1995). Нами установлено, что выступы интеркоксальной пластинки Р4 вселенца несут 5–6 сравнительно крупных и крепких, широких в основании треугольных шипов (рис. 3c, 4a). Вооружение тоже высоких выростов интеркоксальной пластинки Р4 аборигенного вида T. oithonoides характеризуют как 5–6 или более мелких волосообразных шипиков (Рылов, 1948), слабые многочисленные зубчики (Степаньянц и др., 2015) или несколько тонких зубчиков (Монченко, 1974; Mirabdullayev et al., 2003). На большом (100×) увеличении удалось выявить, что по всему свободному краю выступов расположены 6–7 тонких, сравнительно длинных когтеобразых шипов (рис. 3d, 4b). Таким образом, вселенец хорошо отличается от аборигенного вида, как по форме, так и по количеству шипов на выступах интеркоксальной пластинки Р4.
Орнамент из шипиков на каудальной поверхности коксоподита Р4 обоих видов представлен тремя их группами. Для T. taihokuensis все три, а именно: 7–12 шипиков у медиально-дистального края, косой ряд из 9–15 шипиков в латеральной части дистального края и ряд мелких шипиков вблизи проксимального обода коксоподита Р4, показаны в описании вида в ряде работ (Guo, 1999; Dela Paz et al., 2016). В иллюстрациях к другим статьям показаны 1–2 группы (Степаньянц и др., 2015; Mirabdullayev et al., 2003) или орнаментация совсем не отражена (Определитель …, 1995; Mirabdullayev et al., 1997; Monchenko, 2008), также и для T. oithonoides (Монченко, 1974; Определитель …, 2010; Степаньянц и др., 2015; Mirabdullayev et al., 2003). Мы установили, что все три группы шипиков у обоих видов расположены одинаково. Однако в группе Е – косой ряд в латеральной части дистального края коксоподита Р4, у этих двух видов число и размер шипиков различны (рис. 4). Это дает возможность использовать данный показатель как вспомогательный для отличия вселенца от аборигенного вида.
Для обоих видов как особенность вооружения базиподита Р1 часто указывают длинную щетинку на внутреннем его крае, которая достигает начала дистального членика эндоподита (Монченко, 1974; Mirabdullayev et al., 2003; Monchenko, 2008). Мы выяснили, что щетинка на базиподите Р1 у T. taihokuensis короче, чем у T. oithonoides. У T. taihokuensis она достигает начала третьего (дистального) сегмента эндоподита Р1, а у T. oithonoides – середины этого сегмента (рис. 5). Однако, возможно, что это особенности только двух исследованных локальных популяций.
Для обоих видов указывают почти одинаковую длину придатков дистального сегмента Р5, разница в длине не более 10–30% (Монченко, 1974; Степаньянц и др., 2015; Guo, 1999; Mirabdullayev et al., 2003; Monchenko, 2008). Этому полностью соответствуют наши данные для самок, но мы обнаружили, что у самцов T. oithonoides шип Р5 в 1.5 раза длиннее апикальной щетинки. Мы также выявили, что у самок и самцов вселенца укорочена проксимальная щетинка Р5, она составляет 60–85% длины апикальной щетинки (табл. 2). В то же время, у аборигенного T. oithonoides проксимальная щетинка Р5 достигает 85–110% длины апикальной. Это может служить дополнительным признаком, позволяющим различить самцов исследованных видов. У самцов вселенца проксимальная щетинка Р5 короче дистальной, у самцов нативного вида она длиннее дистальной щетинки.
Для таксономической идентификации самцов копепод часто используют строение рудиментарной Р6 (Монченко, 1974; Определитель …, 2010; Mirabdullayev et al., 2003). Из анализа литературных данных следует, что для T. taihokuensis и T. oithonoides соотношение длинной латеральной щетинки и шипа Р6 варьируют примерно в одинаковых пределах (2–2.5 и 1.5–2.5 соответственно) (Монченко, 1974; Степаньянц и др., 2015; Mirabdullayev et al., 2003; Monchenko, 2008). По нашим данным, различия между видами по этому признаку не превышают 30% и не позволяют отличить самцов вселенца от аборигенного вида.
Палеарктический планктонный вид T. oithonoides обычен в малых и больших водоемах, а также в пелагиали обширных водохранилищ на крупных реках России (Рылов, 1948; Монченко, 1974; Степаньянц и др., 2015; Лазарева и др., 2018; Lazareva, 2020). Вселенца из восточной Азии T. taihokuensis характеризуют также как планктонный вид, в Центральной Азии он населяет преимущественно малые водоемы (пруды, рисовые поля и лужи) (Mirabdullayev et al., 2003), но также многочислен в Капшагайском водохранилище на р. Или в Казахстане (Шарапова, 2011; Калымбетова, 2017). Вид населяет не только пресные, но и солоноватые воды, известен из Каспийского моря (Шарапова, 2014; Степаньянц и др., 2015; Monchenko, 2008). В европейской части России T. taihokuensis обитает в крупных пресноводных водохранилищах Средней Волги, преимущественно в литорали и устьевых областях притоков, а также в незарегулированной части р. Волги (Lazareva et al., 2022). Вселенец крайне многочислен в пелагиали Цимлянского водохранилища на р. Дон, а также в малых водохранилищах Волго-Донского канала (Лазарева, Сабитова, 2021). Есть сведения о почти полном вытеснении вселенцем аборигенного T. crassus в Цимлянском водохранилище (Вехов и др., 2014), однако в водохранилищах р. Волги подобного не отмечено ни по отношению к T. crassus, ни по отношению к T. oithonoides (Lazareva et al., 2022).
ЗАКЛЮЧЕНИЕ
Сравнение двух морфологически близких видов T. taihokuensis и T. oithonoides из водоемов центра европейской части России позволило уточнить особенности их строения и обнаружить новые признаки, позволяющие отличить вселенца от аборигенного вида. Выявлено, что различение данных двух видов основывается преимущественно на качественных характеристиках (форма элемента, его расположение и т.п.). Установлено, что для идентификации самок T. taihokuensis и T. oithonoides наиболее важны форма семяприемника, апикальных щетинок каудальных ветвей, апикальных шипов эндоподита Р4 и положение боковой щетинки каудальных ветвей. В качестве дополнительных признаков впервые предложены вооружение выростов интеркоксальной пластинки Р4, орнаментация коксоподита и интеркоксальной пластинки Р4, а также 14-го членика антеннул самок. Показано, что самцы T. taihokuensis существенно крупнее таковых T. oithonoides, они различаются также по положению боковой щетинки каудальных ветвей и длине средних апикальных щетинок. Не выявлено таксономически значимых различий в измеряемых параметрах элементов четвертой, пятой и шестой (у самцов) пар ног, каудальных ветвей и их щетинок.
Список литературы
Вехов Д.А. Науменко А.Н., Горелов В.П., Голоколенова Т.Б., Шевлякова Т.П., 2014. Современное состояние и использование водных биоресурсов Цимлянского водохранилища (2009–2013 гг.) // Рыбохозяйственные исследования на водных объектах Европейской части России. СПб.: Гос. науч.-исслед. ин-т озерного и речного рыбного хоз-ва (ГосНИОРХ). С. 116–145.
Жихарев В.С., Гаврилко Д.Е., Шурганова Г.В., 2019. Находка тропического вида Thermocyclops taihokuensis (Harada, 1931) (Copepoda: Cyclopoida) в Европейской части России // Поволжский экологический журн. № 2. С. 264–270.
Калымбетова М.Т., 2017. Современное состояние зоопланктона Шардаринского водохранилища // Вестник сельскохозяйственной науки Казахстана. № 5–6. С. 80–83.
Лазарева В.И., 2022. Распространение некоторых понто-каспийских и чужеродных копепод (Crustacea, Copepoda) в планктоне водоемов бассейна реки Дон // Российский журнал биологических инвазий. № 3. С. 79–98. https://doi.org/10.35885/1996-1499-15-3-79-98
Лазарева В.И., Сабитова Р.З., Быкова С.В., Жданова С.М., Соколова Е.А., 2018. Распределение летнего зоопланктона в каскаде водохранилищ Волги и Камы // Труды Ин-та биологии внутренних вод РАН. Вып. 83 (86). С. 62‒84. https://doi.org/10.24411/0320-3557-2018-10030
Лазарева В.И., Сабитова Р.З., 2021. Зоопланктон Цимлянского водохранилища и канала Волга-Дон // Зоологический журнал. Т. 100. № 4. С. 1‒15. https://doi.org/10.31857/S0044513421040115
Монченко В.I., 1974., Щелепноротi циклопоподiбнi. Циклопи (Cyclopidae). Киев: Наукова думка. 452 с. (Фауна України; Т. 27, вип. 3).
Нечаев Д.Ю. 2016., Фаунистическое разнообразие планктонных беспозвоночных Волго-Ахтубинской поймы // Материалы Всерос. молодежной гидробиол. конференции “Перспективы и проблемы современной гидробиологии”, пос. Борок, Ин-т биологии внутренних вод им. И.Д. Папанина РАН, 10–13 ноября 2016 г. Ред. И.С. Турбанов, Я.С. Климова, С.Ю. Синельников. Ярославль: Филигрань. С. 117–119.
Определитель пресноводных беспозвоночных России и сопредельных территорий, 1995. Т. 2. Ракообразные. Ред. В.Р. Алексеев. СПб.: Зоологический ин-т РАН. 629 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России, 2010. Т. 1. Зоопланктон. Ред. В.Р. Алексеев. М.: Товарищество научных изданий КМК. 495 с.
Рылов В.М., 1948. Фауна СССР. Ракообразные. Cyclopoida пресных вод. Т. 3. Вып. 3. М.-Л.: АН СССР. 318 с.
Степаньянц С.Д., Хлебович В.В., Алексеев В.Р., Данелия М.Е., Петряшев В.В., 2015. Определитель рыб и беспозвоночных Каспийского моря. Т. 2. Стрекающие, гребневики, многощетинковые черви, веслоногие ракообразные и мизиды. СПб.–М.: Товарищество научных изданий КМК. 244 с.
Шарапова Л.И., 2011. Интегральная оценка экологического состояния зоопланктоценозов Капшагайского водохранилища // Вестник Казахского национального ун-та (КазНУ). Сер. биологическая. № 5. С. 105–109.
Шарапова Л.И., 2014. Зоопланктон Северо-Восточного Каспия. Приложение 2 // Мониторинг окружающей природной среды Северо-Восточного Каспия при освоении нефтяных месторождений. Алматы: АджипККО. С. 228–229.
Chaicharoen R., Sanoamuang L., Hołyńska M., 2011. A Review of the Genus Thermocyclops (Crustacea: Copepoda: Cyclopoida) in Cambodia // Zoological Studies. V. 50. № 6. P. 780–803.
Einsle U., 1993. Crustacea, Copepoda: Calanoida und Cyclopoida. Stuttgart: Gustav Fischer Verlag. 209 p.
Dela Paz E.S.P., Holyn’ska M.K., Papa R. S., 2016. Mesocyclops and Thermocyclops (Copepoda, Cyclopidae) in the major visayas islands (Central Philippines) // Crustaceana. V. 89. № 6–7. P. 787–809. https://doi.org/10.1163/15685403-00003547
Guo X., 1999. The genus Thermocyclops Kiefer, 1927 (Copepoda: Cyclopidae) in China // Hydrobiologia. V. 403. P. 87–95.
Lazareva V.I., 2020. Long-Term Changes in the Composition and Abundance of the Zooplankton Community in Kama River Reservoirs // Inland Water Biology. V. 13. № 2. P. 214–229. https://doi.org/10.1134/S199508292002025X
Lazareva V.I., Zhdanova S.M., Sabitova R.Z., 2022. The Spread of East Asian Copepod Thermocyclops taihokuensis (Harada, 1931) (Crustacea, Copepoda) in the Volga River Basin // Inland Water Biology. V. 15. № 2. P. 139‒148. https://doi.org/10.1134/S1995082922010059
Mirabdullayev I.M., Kuzmetov A.R., 1997. The Genus Thermocyclops (Crustacea: Copepoda) in Uzbekistan (Central Asia) // International Review of Hydrobiology. V. 82. № 2. P. 201–212.
Mirabdullayev I.M., Sharapova L.I., Turemuratova G.I., 1997. A Far East Cyclopoid, Thermocyclops taihokuensis (Harada, 1931) in Central Asia // Turkish Journal of Zoology. V. 21. P. 175–178.
Mirabdullayev I.M., Reid J. W., Ueda H., 2003. Genus Thermocyclops Kiefer, 1927 // Copepoda: Cyclopoida genera Mesocyclops and Thermocyclops. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 20. H. Ueda & J.W. Reid eds. Leiden: Backhuys Publishers. P. 214–302.
Monchenko V.I., 2008. Redescription of the Oriental Thermocyclops taihokuensis (Harada, 1931) (Copepoda: Cyclopoida) from its westernmost population // Zoology in the Middle East. V. 43. № 1. P. 99–104. https://doi.org/10.1080/09397140.2008.10638274
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал