

Зоологический журнал, 2022, T. 101, № 2, стр. 202-212
Применение модели эпигенетического ландшафта для изучения структуры морфотипической изменчивости (на примере M3 скальных полевок (Alticola, Rodentia, Arvicolinae))
А. А. Поздняков *
Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
* E-mail: pozdnyakov61@gmail.com
Поступила в редакцию 30.11.2020
После доработки 06.05.2021
Принята к публикации 14.05.2021
- EDN: LGUJQL
- DOI: 10.31857/S0044513421120096
Аннотация
Изучена морфотипическая изменчивость жевательной поверхности третьего верхнего моляра шести видов скальных полевок. Выделено 49 морфотипов. На основе учета морфотипов на левой и правой сторонах челюсти для изученных видов построены модели эпигенетического ландшафта, которые оказались сходными. Основу модели составляют два морфотипа, преобладающих по частоте. Наибольшая доля асимметричных особей у большинства видов (за исключением тувинской полевки) имеет именно эти морфотипы. Остальные морфотипы в рамках модели эпигенетического ландшафта можно рассматривать как промежуточные, фиксирующие более длинные переходы между двумя основными морфотипами.
Индивидуальное развитие К. Уоддингтон пояснял с помощью модели эпигенетического ландшафта. В этой модели развитие интерпретировалось как скатывание шарика по системе долин, символизирующих развитие признаков от зиготы до конечного состояния. Типичный путь развития какого-либо признака К. Уоддингтон обозначил термином креод (creode). Он охарактеризован двумя параметрами. Во-первых, это креодный профиль, описывающий крутизну креода от начального до конечного состояния. Вполне очевидно, что на разных участках (фазах развития) эта крутизна может быть различной. На участках с высокой крутизной за определенный период происходит много изменений, тогда как на участках с небольшой крутизной за тот же период происходит гораздо меньше изменений. Во-вторых, это форма поперечного сечения долины в эпигенетическом ландшафте. Она характеризует способность системы возвращаться к нормальному развитию при возмущениях, переводящих развитие со дна долины на склон. Чем больше крутизна склона, тем эффективнее работают регуляционные механизмы, возвращая развитие (шарик) на дно долины. В данном случае можно говорить об интенсивности, степени или крутизне канализации (Waddington, 1957).
Согласно модели эпигенетического ландшафта К. Уоддингтона переход на альтернативные траектории развития, заканчивающиеся нетипическими фенотипами, возможен в особых локусах эпигенетического ландшафта, символизируемых невысокими склонами долин.
Необходимо также учитывать, что модель эпигенетического ландшафта Уоддингтона полагалась в ее прямом применении к развитию конкретного индивида. Соответственно, на основе этой модели ничего нельзя сказать о возможных эволюционных изменениях, так как для этого необходимо анализировать популяционную или видовую изменчивость.
Однако данную модель очень легко проинтерпретировать как иллюстрирующую структуру популяционной (видовой) изменчивости (Васильев, 2005). Так, с этой точки зрения система долин отражает развитие разных вариантов (модальностей) одного признака. Скатывание шарика отражает развитие данного варианта у конкретного индивида. Наиболее часто реализуемый вариант представляет собой креод; теоретически – это самая глубокая долина. Другие (нетипичные) варианты отражены в менее глубоких долинах.
В случае, когда мы анализируем коллекционный материал, представленный, как правило, взрослыми и полувзрослыми особями, то на основе этого анализа мы можем говорить лишь о срезе модели эпигенетического ландшафта К. Уоддингтона (рис. 1). Тогда модель можно упростить до кривой линии, на которой углубления представляют срез долины (эпигенетической траектории), а возвышения – это гребни, разделяющие долины. В рамках этой трансформированной модели переход развития с этой траектории на другую можно представить как переход шарика из одной долины в другую под действием возмущающих факторов. Также полагается линейность расположения траекторий развития на срезе.
Рис. 1.
Модель эпигенетического ландшафта К. Уоддингтона (по: Waddington, 1957, с изменениями). A, B – глубина долины по отношению к разным склонам; C – крутизна склона.
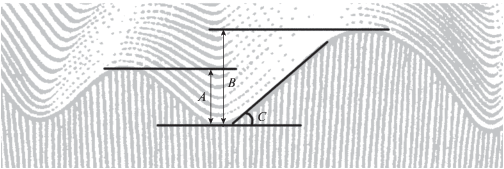
В таком случае возможность шарика перейти в другую долину (реализовать иной фенотип) определяется двумя параметрами: глубиной долины и крутизной склона (рис. 1). К сожалению, формализовать модель (соотнести эти параметры с какими-то свойствами реальных организмов) невозможно, в том числе и по причине различной глубины долины по отношению к разным склонам. Также совершенно необязательно линейное расположение траекторий развития на срезе, и никак нельзя исключать возможность описания их расположения в пространстве большей мерности.
В качестве примера можно указать на изменчивость структуры перемычек овального и круглого отверстий, расположенных на крылоклиновидной кости черепа, у прометеевой полевки (Prometheomys schaposchnikowi Satunin 1901), которая визуализируется в форме двухмерной сети (Васильев, Васильева, 2009, с. 258). Это обстоятельство косвенно свидетельствует в пользу утверждения, что параметры модели эпигенетического ландшафта необходимо устанавливать, основываясь на анализе изменчивости конкретных признаков.
Вполне очевидно, что трансформированная модель эпигенетического ландшафта приложима не ко всем признакам, а лишь к тем, модальности которых обусловлены эпигенетическими факторами. В частности, как считается, асимметрия выражения многих билатеральных признаков обусловлена именно эпигенетическими причинами (Berry, Searle, 1963; Berry, Berry, 1967; Wiig, Lie, 1979; Alberch, 1980; Pankakoski, Hanski, 1989; Васильев, 2005; Поздняков, 2007, 2011).
Жевательная поверхность щечных зубов полевок представляет собой один из таких билатеральных признаков. Значительная изменчивость первого нижнего и третьего верхнего щечных зубов, а также большая доля особей с неодинаковым выражением морфотипов на левой и правой сторонах челюсти свидетельствуют в пользу нестабильности развития этих структур. Также можно полагать значимое влияние эпигенетических факторов на их развитие. В таком случае модель эпигенетического ландшафта вполне применима для описания их изменчивости.
В настоящей статье на основе анализа выражений морфотипов третьего верхнего щечного зуба (M3) на разных сторонах челюсти скальных полевок предпринята попытка построить модель эпигенетического ландшафта, описывающую морфотипическую изменчивость данного признака.
МАТЕРИАЛЫ И МЕТОДИКА
В работе использованы материалы из коллекций Института систематики и экологии животных СО РАН, Новосибирск, и Зоологического музея Московского государственного университета, Москва. Исследована морфотипическая изменчивость М3 шести видов скальных полевок Alticola s. str. Изучены взрослые и полувзрослые особи с полностью сформированной плоской жевательной поверхностью. Всего исследовано 2058 особей. Общая характеристика изменчивости приведена в табл. 1.
Таблица 1.
Характеристика изменчивости М3 скальных полевок
| Вид | n | Nm | Ns | Pas |
|---|---|---|---|---|
| A. argentatus | 363 | 21 | 42 | 17.1 |
| A. strelzowi | 792 | 33 | 65 | 23.0 |
| A. semicanus | 386 | 19 | 38 | 19.9 |
| A. tuvinicus | 136 | 27 | 46 | 25.2 |
| A. olchonensis | 240 | 15 | 31 | 38.7 |
| A. barakshin | 141 | 10 | 20 | 19.9 |
Жевательная поверхность M3 полевок характеризуется значительной изменчивостью. Для ее описания использована авторская методика, основанная на учете количества замкнутых полей и количества выступающих углов на задней непарной петле (ЗНП). Подробно методика описана в работе, касающейся географической изменчивости серых полевок (Поздняков, 1993), и была использована при анализе географической изменчивости скальных полевок (Литвинов и др., 2000; Поздняков и др., 2004), поскольку принципиальных различий в строении щечных зубов у представителей этих двух родов нет. Всего выделено 49 морфотипов жевательной поверхности М3 скальных полевок (рис. 2).

Морфотипы левой и правой сторон верхней челюсти могут быть представлены как одинаковыми, так и разными вариантами, что позволяет говорить о симметричных и асимметричных парах (или сочетаниях) вариантов. Общую долю асимметричных пар в выборке отражает показатель асимметрии.
На основании асимметричных сочетаний можно установить связи между разными морфотипами. Поскольку выделение морфотипов осуществлялось по трем критериям (количеству замкнутых треугольников, количеству выступающих углов на лабиальной стороне ЗНП и количеству выступающих углов на лингвальной стороне ЗНП), то в пространстве этих трех параметров структура связей между морфотипами может быть выражена следующим образом (рис. 3).
Рис. 3.
Структура связей между морфотипами M3 A. strelzowi. Схема построена по направлениям, отражающим увеличение количества 1) замкнутых треугольников и выступающих углов на 2) лабиальной и 3) лингвальной сторонах ЗНП.
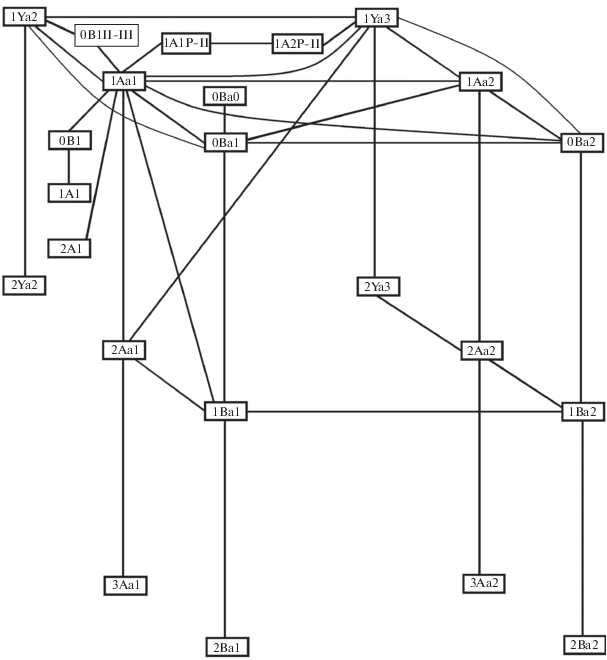
По указанным направлениям морфотипы располагаются в плоскостях, отражающих изменение соответствующего параметра. Так, в случае увеличения количества замкнутых треугольников морфотипы образуют три плоскости: первая: 1Ya2, 1Ya3, 2Ya2 и 2Ya3, вторая: 1Aa1, 2Aa1, 3Aa1, 1Aa2, 2Aa2 и 3Aa2, третья: 0Ba0, 0Ba1, 1Ba1, 2Ba1, 0Ba2, 1Ba2 и 2Ba2 (рис. 3).
В случае увеличения количества выступающих углов на лабиальной стороне ЗНП также можно выделить три плоскости: первая: 1Ya2, 1Ya3, 1Aa1, 1Aa2, 0Ba1, 0Ba2, вторая: 2Ya2, 2Ya3, 2Aa1, 2Aa2, 1Ba1, 1Ba2, третья: 3Aa1, 3Aa2, 2Ba1, 2Ba2 (рис. 3).
В случае увеличения количества выступающих углов на лингвальной стороне ЗНП можно выделить две плоскости: первая: 1Ya2, 2Ya2, 1Aa1, 2Aa1, 3Aa1, 0Ba1, 1Ba1, 2Ba1, вторая: 1Ya3, 2Ya3, 1Aa2, 2Aa2, 3Aa2, 0Ba2, 1Ba2, 2Ba2 (рис. 3).
Остальные морфотипы (1A1P-II, 1A2P-II, 0B1II-III, 1A1, 2A1, 0B1) по количеству замкнутых треугольников относятся к четырем разным группам, и они не формируют четко выраженные плоскости.
Если же связи между морфотипами выстроить “экономным” способом, то получается почти плоская картина (рис. 4).
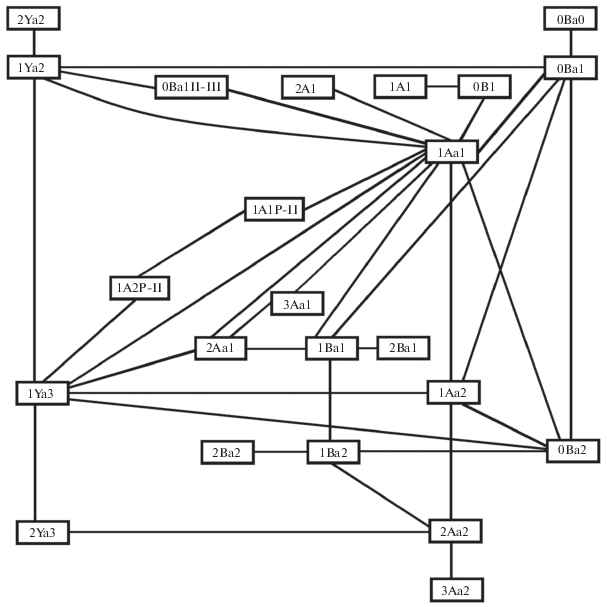
Полученную картинку можно интерпретировать как срез эпигенетического ландшафта. Элемент эпигенетического ландшафта – морфотип – представляет собой реализацию морфологической структуры в ходе индивидуального развития. Соответственно, линия формирования моляра с определенным морфотипом (процесс становление морфотипа рисунка жевательной поверхности) в ходе индивидуального развития представляет собой морфогенетическую траекторию. На основании структуры связей между морфотипами (рис. 3 и 4) можно принять, что морфогенетические траектории на срезе эпигенетического ландшафта рисунка жевательной поверхности M3 скальных полевок располагаются не линейно, как моделировал К. Уоддингтон, и не в трехмерном пространстве, как это можно было предположить на основании критериев выделения морфотипов, а в плоскости.
В таком случае в отношении рисунка жевательной поверхности M3 можно принять, что морфогенетическая траектория представляет собой не долину, как это полагается в модели эпигенетического ландшафта К. Уоддингтона, а тоннель. В качестве количественного показателя, характеризующего морфогенетическую траекторию, можно предложить диаметр тоннеля. С этим показателем можно прямо сопоставить частоту особей в выборке с одинаковыми морфотипами на разных сторонах челюсти.
В идеальных условиях, как предполагается, моляры на разных сторонах челюсти должны быть одинаковыми, т.е. в контексте модели эпигенетического ландшафта можно сказать, что осуществление обоих зубов (их морфотипов) проходит по одной и той же морфогенетической траектории (тоннелю). Случаи, когда на разных сторонах челюсти встречены различные морфотипы, в контексте модели эпигенетического ландшафта можно объяснить тем, что в онтогенезе развитие одного из зубов стало осуществляться по другой морфогенетической траектории, т.е. в процессе развития произошел переход на другую траекторию. В рамках модели этот переход между траекториями можно представить как соединяющий их тоннель. В отношении модели следует заметить, что расстояние между морфогенетическими траекториями не имеет значения. Так, если развитие представить как прохождение шарика по тоннелю (морфогенетической траектории), то при попадании шарика в тоннель между траекториями он обязательно достигнет иной траектории. В таком случае, поскольку мы приняли, что частота особей соотносится с диаметром тоннеля морфогенетической траектории, то и диаметр тоннеля между морфогенетическими траекториями необходимо соотносить с частотой особей с асимметричным сочетанием морфотипов на разных сторонах челюсти.
Чтобы разгрузить схемы, из них исключили все морфотипы, зафиксированные менее чем у пяти особей в одинаковых вариантах на разных сторонах челюсти (в симметричном состоянии), за исключением тувинской и ольхонской полевок, у которых исключили морфотипы, зафиксированные в симметричном состоянии единично.
РЕЗУЛЬТАТЫ
У гобийской полевки (Alticola barakshin Bannikov 1947) наибольшая доля особей с одинаковыми морфотипами на разных сторонах челюсти имеет морфотип 1Aa0 (рис. 5А). По сравнению с преобладающими по частоте морфотипами у других изученных видов скальных полевок у гобийской полевки этот морфотип наиболее простой. Второй по встречаемости морфотип – это 1Aa1. Способ усложнения – появление зубца на лингвальной стороне ЗНП. Наибольшая доля особей с асимметричным проявлением морфотипов – с парой морфотипов 1Aa0 и 1Aa1.
Рис. 5.
Структура связей между морфотипами M3: А – A. barakshin, Б – A. strelzowi, В – A. semicanus, Г – A. olchonensis, Д – A. argentatus, Е – A. tuvinicus.
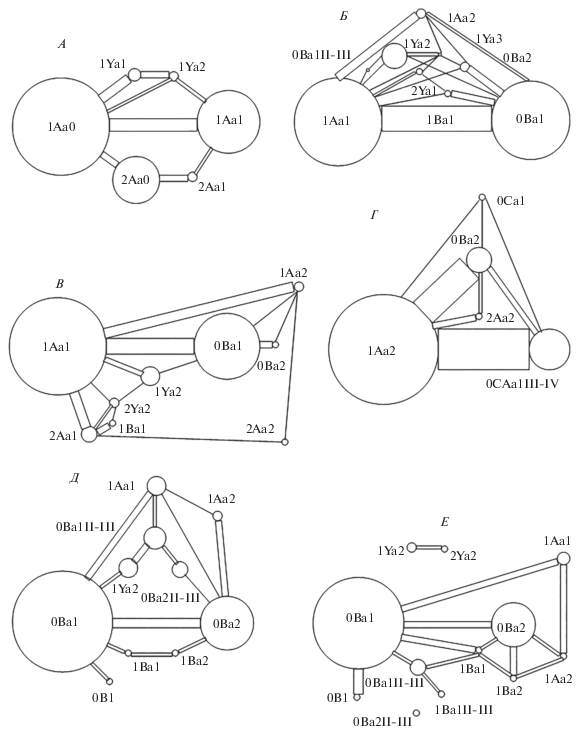
У плоскочерепной (A. strelzowi Kastschenko 1901) и хангайской (A. semicanus Allen 1924) полевок у наибольшей доли особей представлен морфотип 1Aa1 (рис. 5Б, 5В), у которого по сравнению с 1Aa0 имеется дополнительный выступающий зубец на лингвальной стороне ЗНП. У обоих этих видов второй по встречаемости морфотип 0Ba1, который получается из 1Aa1 путем замыкания треугольника на лабиальной стороне ЗНП. Также у обоих этих видов наибольшая доля особей с асимметричными морфотипами имеет именно эту пару морфотипов.
У ольхонской полевки (A. olchonensis Litvinov 1960) наибольшая доля особей имеет морфотип 1Aa2 (рис. 5Г), у которого, по сравнению с предыдущими видами, добавляется еще один зубец на лингвальной стороне ЗНП. Второй по встречаемости морфотип – это 0Ca1III-IV. Способ усложнения – смыкание полосок репаративного дентина после четвертого зубца с образованием замкнутого поля, в котором имеются зубцы как с лабиальной, так и с лингвальной стороны ЗНП. Наибольшая доля асимметричных особей именно с этими морфотипами. Но также со значительной частотой представлен и морфотип 0Ba2.
У серебристой (A. argentatus Severtzov 1879) и тувинской (A. tuvinicus Ognev 1950) полевок наибольшая доля особей имеет морфотип 0Ba1 (рис. 5Д, 5Е), который получается из морфотипа 1Aa1 путем замыкания лабиального зубца на ЗНП. У обоих этих видов второй по встречаемости морфотип 0Ba2. Способ усложнения – появление еще одного зубца на лингвальной стороне ЗНП.
ОБСУЖДЕНИЕ
Описываемая в данной статье модель эпигенетического ландшафта отличается от модели К. Уоддингтона не только изобразительными особенностями, но и направленностью объяснения. Собственно, модель эпигенетического ландшафта К. Уоддингтона направлена на объяснение стабильности индивидуального развития. В этой модели склоны долин рассматриваются как имеющие разную крутизну и высоту. Соответственно, регуляция развития возможна, если склоны имеют среднюю крутизну. Моделируется, что различные возмущения переводят развитие со дна долины на склон. При достаточной высоте склона и его крутизне развитие снова возвращается на дно долины. Таким образом, развитие восстанавливается после нарушения. Если же склоны крутые и высокие, то развитие оказывается детерминированным (Уоддингтон, 1947).
Развитие в этой модели представляется как движение по сложно разветвленной системе долин, в которой долины могут как разветвляться, так и сливаться. В случае, если долины разветвляются на одном уровне, то выбор направления развития может осуществляться случайным образом в результате незначительного колебания условий, в которых осуществляется развитие. Если же одна из долин располагается выше другой, то переход в эту долину возможен в случае возмущения, достаточного для преодоления порога.
Наша модель направлена на объяснение морфологического разнообразия и его структуры. Собственно, срез системы тоннелей – это совокупность реализованных вариантов строения (в нашем случае – морфотипов) на видовом уровне. В данном случае тоннели отображают траектории развития вариантов, а креодом будет называться траектория развития с наиболее часто реализуемым вариантом.
Траектории развития не изолированы друг от друга, поскольку в процессе развития возможен переход с одних траекторий на другие. Такие переходы регистрируются на особях, у которых на разных сторонах челюсти формируются различные морфотипы. Частота таких асимметричных особей отражает вероятность взаимных переходов между данными траекториями. Срез траекторий развития с указанием переходов между ними отражает структуру изменчивости, т.е. популяционный или видовой (в нашем случае) эпигенетический ландшафт (рис. 5).
К нашей модели очень близки схемы, отражающие структуру изменчивости перемычек круглого и овального отверстий у прометеевой полевки и морфотипов рисунка жевательной поверхности M3 лемминговидной полевки (Alticola lemminus Miller 1899) (Васильев, Васильева, 2009). Различия заключаются в том, что на схемах А.Г. Васильева указано количество фенов (антимер), тогда как на наших схемах – количество особей. Также графика наших схем масштабирована пропорционально количеству особей, тогда как графика схем А.Г. Васильева – не масштабирована. В целом, в выразительном отношении различия между схемами А.Г. Васильева и нашими непринципиальные.
Однако важнейшим моментом общетеоретических представлений Васильева (2005, 2008, 2009; Васильев, Васильева, 2009) является попытка синтеза эпигенетической теории и мерономии С.В. Мейена. В этом контексте мерон интерпретируется им как множество возможных (вероятных) состояний (модусов) признака. Разные таксоны могут характеризоваться одинаковым спектром модусов какого-либо признака, но могут различаться по частоте представленности этих модусов. Согласно А.Г. Васильеву, концепцию мерона можно объединить с моделью эпигенетического ландшафта11. Это объединение позволяет указать на процессуальный момент: “мерон представляет собой морфогенетический закон трансформации структуры, порождающий характерное полиморфное множество” (Васильев, 2004, с. 19).
На этой теоретической основе схемы, отражающие изменчивость структуры перемычек круглого и овального отверстий у прометеевой полевки и морфотипов рисунка жевательной поверхности M3 лемминговидной полевки, он интерпретирует как визуализацию мерона (Васильев, Васильева, 2009, с. 257, 259), но не визуализацию эпигенетического ландшафта.
В качестве примера визуализации эпигенетического ландшафта он приводит “проекцию векторов, исходно вычисленных для каждой особи по главным компонентам 107 гомологичных фенов неметрических признаков” трех видов хомяков (Васильев, Васильева, 2009, с. 278).
Но указанный пример отражает оценку межвидовой дивергенции, выполненную на билатеральных признаках, изменчивость которых обусловлена эпигенетическими причинами.
Вполне очевидно, что модель – это представление определенного аспекта реальности, позволяющее получить ответы на изучаемые вопросы. Соответственно, выбор формы модели должен определяться поставленными вопросами.
Материалом исследования во всех указанных случаях является популяционная и видовая изменчивость билатеральных признаков, модальности которых обусловлены эпигенетическими причинами. На исследуемом коллекционном материале зафиксированы конечные состояния признаков, являющиеся результатом определенной траектории развития. В первом случае (изменчивость перемычек круглого и овального отверстий у прометеевой полевки и морфотипов рисунка жевательной поверхности M3 полевок) визуализируются конечные состояния траекторий развития разных вариантов одного признака. Во втором случае (фены неметрических признаков черепа хомяков) визуализируются конечные состояния обобщенных траекторий развития особей, полученных в результате статистического анализа большого комплекса билатеральных признаков. Все эти примеры допустимо рассматривать как примеры визуализации эпигенетического ландшафта. Проблема заключается в том, на какие вопросы мы можем получить ответы с помощью этих визуализаций.
Так, во втором случае мы получаем оценку межвидовых дистанций, которую можно получить не только на эпигенетических, но и на молекулярных, морфометрических и еще каких-нибудь признаках. А в первом случае мы визуализируем структуру эпигенетических связей между вариантами какого-либо признака, которую можно получить только описанным способом.
Итак, с помощью предложенной модели выясняется, что структура эпигенетического ландшафта у всех изученных видов скальных полевок сходна. Ее основу составляют два морфотипа, преобладающих по частоте. Наибольшая доля асимметричных особей у большинства видов (за исключением тувинской полевки) именно с этими морфотипами. Остальные морфотипы в рамках модели эпигенетического ландшафта можно рассматривать как промежуточные, фиксирующие более длинные переходы между двумя основными морфотипами. Иными словами, между двумя основными морфотипами существует несколько путей перехода, а в некоторых случаях, как у плоскочерепной полевки (рис. 5Б), таких путей довольно много. Более длинные пути распознаются посредством одного или нескольких морфотипов, через которые они проходят.
Сопоставляя модели эпигенетического ландшафта изученных видов скальных полевок, отношения между двумя основными морфотипами можно трактовать как показывающие направление изменения эпигенетического ландшафта, т.е. возможную смену креода другим путем развития. В морфотипическом отношении это будет выражаться в смене одного преобладающего в данный момент времени по частоте морфотипа другим морфотипом. В таком случае можно сказать, что основная тенденция заключается в увеличении количества зубцов на лингвальной стороне ЗНП: 1Aa0 (A. barakshin) → 1Aa1 (A. strelzowi; A. semicanus) → 1Aa2 (A. olchonensis); 0Ba1 → 0Ba2 (A. tuvinicus; A. argentatus). Дополнительная тенденция заключается в замыкании полей: 1Aa1 → 0Ba1 (A. strelzowi; A. semicanus); 1Aa2 → 0Ca1III-IV (A. olchonensis). В основном тренде усложнения морфотипов отсутствует появление дополнительных зубцов на лабиальной стороны ЗНП.
Согласно молекулярным данным филогенетические отношения между изученными видами скальных полевок реконструируются следующим образом. Раньше всех обособляется гобийская полевка, затем серебристая (Lebedev et al., 2007), затем плоскочерепная (Литвинов и др., 2015). Остальные три вида по разным методикам дают разные соотношения, но последней обособляется тувинская полевка, которая состоит в сестринском отношении или с ольхонской, или с хангайской полевкой (Литвинов и др., 2015). Таким образом, положение изученных видов скальных полевок в ряду усложнения морфотипов диссонирует с последовательными этапами их филогенеза, опирающимися на молекулярные данные.
Согласно палеонтологическим данным наиболее древние остатки полевок, отнесенных к роду Alticola, найдены в среднем плейстоцене Китая, провинция Хэбэй (Zheng, Cai, 1991). Обнаружено несколько щечных зубов, среди которых хорошо сохранившийся M3 относится к морфотипу 0Ya1.
На территории России, согласно палеонтологическим данным (Галкина, Оводов, 1975; Alexeeva, 1998; Оводов и др., 1992; Деревянко и др., 2001; Agadjanian, Serdyuk, 2005), скальные полевки встречаются в отложениях, начиная с верхнего плейстоцена, причем видовая идентификация остатков затруднена. Принимая во внимание аллопатрическое распространение современных видов скальных полевок (Rossolimo, Pavlinov, 1992), их формирование следует рассматривать как процесс географической дифференциации в пределах ареала предковой формы. Учитывая также влияние температурных условий среды обитания полевок на их морфотипическую изменчивость (Поздняков, 2003, 2004; Montuire, Brunet-Lecompte, 2004), можно предположить, что различия в структуре эпигенетического ландшафта M3 изученных видов скальных полевок, выражающиеся в первую очередь в преобладающих по частоте морфотипах, обусловлены различиями в условиях обитания.
Следует подчеркнуть, что эти различия выражаются на фоне эволюционной тенденции к усложнению очертания жевательной поверхности M3. Иными словами, спектр морфотипической изменчивости скальных полевок при формировании современных видов на разных территориях определялся парой доминирующих по частоте морфотипов. Причем во всех случаях преобладающий по частоте морфотип оказался проще второго по частоте морфотипа. Однако эта тенденция к усложнению не является жестко детерминированной. Так, изменчивость включает спектр морфотипов, которые могут быть как сложнее, так и проще морфотипа, преобладающего по частоте.
В контексте модели эпигенетического ландшафта все эти морфотипы отражают окольные пути переходов между двумя доминирующими морфотипами, но сама структура эпигенетического ландшафта оказывается неповторимой, что выявляется при сравнении видов с одинаковыми доминирующими морфотипами (A. strelzowi и A. semicanus; A. argentatus и A. tuvinicus) (рис. 5). Указанные пары видов в филогенетическом отношении относительно далеки. Но филогенетически близкие виды (A. semicanus, A. olchonensis и A. tuvinicus) как раз различаются по доминирующим морфотипам.
Если основываться на широко распространенном представлении, что эволюция выражается в постепенной дивергенции признаков, в том числе и эпигенетических, то по отношению к другим видам такое достаточно сильное различие филогенетически близких видов (A. semicanus, A. olchonensis и A. tuvinicus) по составу преобладающих по частоте морфотипов оказывается необъяснимым.
Но в данном случае различия в частотах морфотипов связаны с различиями в эпигенетических ландшафтах. Есть теоретические основания полагать, что перестройка эпигенетического ландшафта при видообразовании носит не постепенный, а пороговый характер. Соответственно, следствием таких перестроек эпигенетического ландшафта будут резкие изменения в частотах морфотипов. Поскольку фактического материала по визуализации эпигенетического ландшафта для близких видов животных крайне мало, то в отношении различий эпигенетических ландшафтов близких видов можно высказать лишь предварительные суждения, носящие предположительный характер. Необходимо накопление фактического материала, чтобы высказать определенные утверждения.
ЗАКЛЮЧЕНИЕ
Модель эпигенетического ландшафта К. Уоддингтона нередко упоминается как в работах, посвященных теоретическим проблемам эволюционной теории (Шишкин, 1987), так и в работах, посвященных изучению изменчивости (Васильев, 2005; Васильев, Васильева, 2009).
Способ визуализации модели эпигенетического ландшафта, разработанный А.Г. Васильевым, перспективен, но не используется другими исследователями.
С помощью описанной в данной статье усовершенствованной методики визуализации модели эпигенетического ландшафта изучена морфотипическая изменчивость M3 скальных полевок и установлены некоторые общие закономерности.
Так, структура изменчивости определяется двумя морфотипами, преобладающими по частоте. Между этими морфотипами имеется не только прямой переход, который можно выявить по особям, у которых на разных сторонах челюстей имеются различные морфотипы, но и выявлено несколько дополнительных переходов, опосредованных другими морфотипами и потому более длинных.
Модель эпигенетического ландшафта, изложенная в настоящей статье, может быть применена для исследования популяционной и видовой изменчивости любых вариабельных дискретных билатеральных признаков, обладающих в той или иной степени асимметрией проявления на разных сторонах (фолидоз головы рептилий, рисунок на крыльях насекомых, жилкование крыльев насекомых и т.п.).
Список литературы
Васильев А.Г., 2004. Феногенетическая изменчивость и популяционный онтогенез // Ученые записки НТГСПА. С. 13–23.
Васильев А.Г., 2005. Эпигенетические основы фенетики. На пути к популяционной мерономии. Екатеринбург: Академкнига. 640 с.
Васильев А.Г., 2008. Популяционная мерономия и проблема визуализации эпигенетического ландшафта // XXII Любищевские чтения. Т. 2. Ульяновск: Ульянов. гос. пед. ун-т. С. 6–15.
Васильев А.Г., 2009. Феногенетическая изменчивость и популяционная мерономия // Журнал общей биологии. Т. 70. № 3. С. 195–209.
Васильев А.Г., Васильева И.А., 2009. Гомологическая изменчивость морфологических структур и эпигенетическая дивергенция таксонов: основы популяционной мерономии. М.: Товарищество научных изданий КМК. 511 с.
Галкина Л.И., Оводов Н.Д., 1975. Антропогеновая териофауна пещер Западного Алтая // Систематика, фауна, зоогеография млекопитающих и их паразитов. Новосибирск: Наука. С. 165–180.
Деревянко А.П., Агаджанян А.К., Кулик Н.А., Мартынович Н.В., Оводов Н.Д., Постнов А.В., Сердюк Н.В., Чевалков Л.М., 2001. Основные результаты изучения многослойного памятника Усть-Канская пещера в 1998–2001 гг. // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 7. С. 109–114.
Литвинов Н.И., Литвинов Ю.Н., Поздняков А.А., 2000. Морфологическая изменчивость изолированных популяций ольхонской полевки Alticola olchonensis (Rodentia, Arvicolidae) с островов Байкала // Зоологический журнал. Т. 79. № 1. С. 73–79.
Литвинов Ю.Н., Абрамов С.А., Чертилина О.В., Симонов Е.П., Лопатина Н.В., 2015. Генетическая дифференциация скальных полевок в горно-степных районах северо-востока Внутренней Азии // Изв. Иркут. гос. ун-та. Серия: Биология. Экология. Т. 12. С. 23–30.
Мейен С.В., 1974. О соотношении номогенетического и тихогенетического аспектов эволюции // Журнал общей биологии. Т. 35. № 3. С. 353–364.
Оводов Н.Д., Мартынович Н.В., Поздняков А.А., Орлова Л.А., 1992. Млекопитающие и птицы в окрестностях Красноярского края на рубеже плейстоцен–голоцен. Красноярск: ИАЭ СО РАН. С. 197–200.
Поздняков А.А., 1993. Морфотипическая изменчивость жевательной поверхности коренных зубов серых полевок группы “maximowiczi” (Rodentia, Arvicolidae): опыт количественного статистического анализа // Зоологический журнал. Т. 72. Вып. 11. С. 114– 125.
Поздняков А.А., 2003. Морфотипическая изменчивость серых полевок (Rodentia, Arvicolidae, Microtus) в связи с температурными условиями среды // Успехи современной биологии. Т. 123. № 2. С. 187–194.
Поздняков А.А., 2004. Билатеральная асимметрия морфотипов жевательной поверхности коренных зубов полевки-экономки Microtus oeconomus Pallas (Rodentia, Arvicolidae) // Успехи современной биологии. Т. 124. № 4. С. 371–377.
Поздняков А.А., 2007. Структура морфотипической изменчивости серых полевок (Microtus: Rodentia, Arvicolidae) с точки зрения эпигенетической теории эволюции // Успехи современной биологии. Т. 127. № 4. С. 416–424.
Поздняков А.А., 2011. Структура морфологической изменчивости (на примере морфотипов жевательной поверхности первого нижнего коренного зуба серых полевок) // Журнал общей биологии. Т. 72. № 2. С. 127–139.
Поздняков А.А., Абрамов С.А., Лопатина Н.В., Литвинов Ю.Н., 2017. Морфотипическая изменчивость плоскочерепной полевки (Alticola strelzowi, Rodentia, Arvicolinae) // Зоологический журнал. Т. 96. № 10. С. 1267–1278.
Поздняков А.А., Литвинов Ю.Н., Лопатина Н.В., 2004. Эколого-морфологическая изменчивость алтайских популяций плоскочерепной полевки Alticola strelzowi Kastschenko (Rodentia, Cricetidae, Arvicolinae) // Сибирский экологический журнал. № 4. С. 579–587.
Уоддингтон К.Х., 1947. Организаторы и гены. М.: Госиздат. 240 с.
Шишкин М.А., 1987. Индивидуальное развитие и эволюционная теория // Эволюция и биоценотические кризисы. М.: Наука. С. 76–124.
Agadjanian A.K., Serdyuk N.V., 2005. The history of mammalian communities and paleogeography of the Altai Mountains in the Paleolithic // Paleontological Journal. V. 39. № Suppl. 6. P. 645–821.
Alberch P., 1980. Ontogenesis and morphological diversification // Am. Zool. V. 20. P. 653–667.
Alexeeva N.V., 1998. Paleoecology of Transbaikal Allophaiomys faunas // Paludicola. V. 2. № 1. P. 16–19.
Berry A.C., Berry R.J., 1967. Epigenetic variation in the human cranium // J. Anat. V. 101. P. 361–379.
Berry R.J., Searle A.G., 1963. Epigenetic polymorphism of the rodent skeleton // Proc. Zool. Soc. Lond. V. 140. P. 577–615.
Lebedev V.S., Bannikova A.A., Tesakov A.S., Abramson N.I., 2007. Molecular phylogeny of the genus Alticola (Cricetidae, Rodentia) as inferred from the sequence of the cytochrome b gene // Zool. Scripta. V. 36. P. 547–563.
Montuire S., Brunet-Lecomte P., 2004. Relation between climatic fluctuation and morphological variability in Microtus (Terricola) grafi (Arvicolinae, Rodentia) from Bacho Kiro (Bulgaria, Upper Pleistocene) // Lethaia. V. 37. P. 71–78.
Pankakoski E., Hanski I., 1989. Metrical and non-metrical skull traits of the common shrew Sorex araneus and their use in population studies // Ann. Zool. V. 26. P. 433–444.
Rossolimo O.L., Pavlinov I.J., 1992. Species and subspecies of Alticola s. str. (Rodentia: Arvicolidae) // Prague Studies in Mammalogy (eds I. Horacek, V. Vohralik). Prague. P. 149–176.
Waddington C.H., 1957. The strategy of the genes. L.: George Allen & Unwin Ltd. 262 p.
Wiig Ø., Lie R.W., 1979. Metrical and non-metrical skull variations in Norwegian wild mink (Mustela vison Schreber) // Zool. Scripta. V. 8. P. 297–300.
Zheng S., Cai B., 1991. Fossil micromammals from the Donggou section of Dongyaozitou, Yuxian County, Hebei Province // XIII International Congress of INQUA (2–9 August 1991, Beijing, China). Beijing. P. 100–131.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал