

Зоологический журнал, 2022, T. 101, № 2, стр. 213-227
Сравнительный морфотипический анализ первых нижнекоренных зубов восточной полевки Alexandromys fortis (Büchner 1889) островных и ископаемых популяций юга Дальнего Востока России (Приморский край)
М. А. Винокурова a, *, А. Е. Гусев a, М. П. Тиунов a
a Федеральный научный центра биоразнообразия наземной биоты
Восточной Азии ДВО РАН
690022 Владивосток, Россия
* E-mail: vinmary@mail.ru
Поступила в редакцию 15.04.2021
После доработки 02.07.2021
Принята к публикации 09.07.2021
- EDN: AMNAWU
- DOI: 10.31857/S0044513422020088
Аннотация
Проанализирована морфотипическая изменчивость антероконидной формы первого нижнего моляра ископаемых и современных (континентальные и островные популяции) восточных полевок Alexandromys fortis с территории Приморья (юг Дальнего Востока). Морфотипическая изменчивость учитывалась на основании изучения вариаций строения лабиальной и лингвальной сторон передней непарной петли и особенностей строения лингвального входящего угла 4. С помощью построенной комбинационной решетки выявлено 36 морфотипов строения непарной передней петли антероконида m1. В настоящее время наиболее распространен на континенте морфотип IIICa, преобладавший в позднем плейстоцене. Особенности морфотипического состава m1 у выделенных популяций восточной полевки на островах Дурново, Де-Ливрона, Веры (залив Петра Великого) обусловлены как фенотипическими, так и генотипическими особенностями основателей островных популяций. Оригинальность новых условий обитания инициировала высокую скорость эволюции, высвобождение скрытого разнообразия и увеличение числа редких морфотипов.
Несмотря на то, что применение методов геометрической морфометрии показало значительную изменчивость рисунка жевательной поверхности первого нижнего коренного зуба у восточноазиатских полевок рода Alexandromys (Войта и др., 2019), альтернативы использования формы этого зуба при идентификации ископаемых остатков полевок пока нет. Ископаемый материал представлен обычно отдельными зубами и определение видовой принадлежности полевок проводится преимущественно по этому наиболее информативному зубу (Semken, Wallace, 2002; Бородин, Маркова, 2014; Голенищев и др., 2018; Golenishchev et al., 2019). При этом работ, посвященных изучению внутривидовой изменчивости рисунка жевательной поверхности первого нижнего коренного зуба у восточноазиатских видов полевок, за исключением A. oeconomus (Nadachowski, 1982; Костенко, Алленова, 1989; Поздняков, Литвинов, 1994; Фадеева, 2003, 2005), не много. Разными исследователями, занимавшимися ранее изучением изменчивости этого зуба у A. fortis, было выявлено от 3 до 10 морфотипов (Мейер, 1978; Алексеева, Голенищев, 1986; Поздняков, 1993, 2010; Лисовский и др., 2018; Ковалева и др., 2021).
На наш взгляд, наиболее приемлемая методика выделения морфотипов была предложена Поздняковым (1993). Основываясь на этой методике и методе построения классификационной морфологической таблицы, который использовался ранее при изучении жевательной поверхности третьих нижних премоляров северной пищухи (Гусев, Тиунов, 2021), первоочередной задачей исследования было построение подобной таблицы для A. fortis. Необходимо отметить, что недавно была показана возможность использования особенностей морфотипической изменчивости зубов в интерпретации палеохронологических событий в истории Arvicolinae (Markova et al., 2020). Появившиеся в последнее время сведения о морфологической и генотипической изменчивости у островных популяций этого вида на Дальнем Востоке (Шереметьева, 2003, 2020) и имеющийся в нашем распоряжении ископаемый материал определили следующую задачу – изучение морфотипической изменчивости формы антероконида m1 A. fortis в островных популяциях залива Петра Великого (Японское море) и сравнение ее с вековой изменчивостью этого зуба у ископаемых популяций из позднеплейстоценовых и голоценовых отложений пещеры Медвежий Клык (Приморский край).
МАТЕРИАЛ И МЕТОДЫ
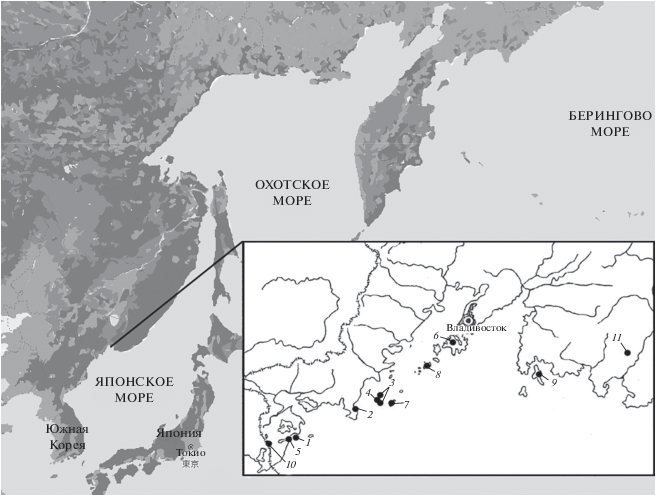
Изучение изменчивости строения жевательной поверхности первых нижних коренных зубов восточной полевки проводилось на коллекционных материалах лаборатории териологии ФНЦ Биоразнообразия ДВО РАН. Было исследовано 165 зубов из 2 локалитетов материковой части юга Приморского края (окрестности пос. Хасан и побережье бухты Астафьева), 341 зуб из выборок с островов залива Петра Великого (Русский – 26 экз., Путятина – 65 экз., Рикарда – 10 экз., Пелис – 10 экз., Веры – 116 экз., Гильдебранта – 8 экз., Де-Ливрона и Дурново – 106 экз.) и 10 экз. с о-ва Фальшивый. Ранее принадлежность островных популяций именно A. fortis была дополнительно подтверждена молекулярно-генетическими методами (Chelomina, Sheremetyeva, 2007; Шереметьева, 2020). Кроме того, был изучен позднеплейстоцен-голоценовый ископаемый материал (125 первых нижних коренных зубов) из пещеры “Медвежий Клык” на хребте Лозовой, Партизанский р-н Приморского края (рис. 1). Войта и соавторы (2019), анализировавшие этот материал, утверждают, что все эти зубы принадлежат “A. fortis, однозначно определяющейся по характерной форме антероконида и крупным размерам …” (Войта и др., 2019, стр. 333)
Рис. 1.
Места сбора материала: 1 – о-в Веры; 2 – бухта Астафьева; 3 – острова Де-Ливрона, Дурново; 4 – о-в Гильдебрандта; 5 – о-в Фальшивый; 6 – о-в Русский; 7 – о-в Большой Пелис; 8 – о. Рикорда; 9 – о-в Путятина; 10 – пос. Хасан; 11 – пещера Медвежий Клык.
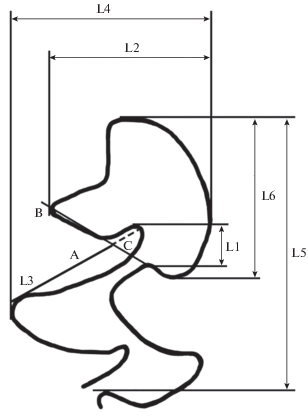
При оценке строения жевательной поверхности m1 использованы как морфотипический подход, так и анализ мерных признаков. При измерении зуба использовалось 6 промеров (рис. 2): L1 – расстояние между вершинами лингвального входящего угла 4 и лабиального входящего угла 3, L2 – ширина передней непарной петли m1, L3 – результат измерения достигается следующими последовательными действиями – а) проводится линия А от вершины лингвального выходящего угла 4 до вершины лингвального входящего угла 4, б) проводится линия В от вершины входящего лабиального угла 3 до вершины выходящего лингвального угла 5, делящая линию А на два отрезка, в) определяется отношение длины отрезка С к общей длине линии А, L4 – ширина антероконида, L5 длина антероконида, L6 – длина передней непарной петли m1.
Для расчета размерных характеристик зубов в работе использовался стандартный пошаговый дискриминантный анализ, без включения и исключения переменных. Классификация групп осуществлялась с помощью вычисления расстояния Махаланобиса и функции матричной классификации методом “складного ножа” (jackknife). Так как объем исследуемой выборки n > 100 для вычисления достоверности расстояний Махаланобиса использовалось значение уровня значимости p-level, равное 0.01 (Наследов, 2004). Все расчеты выполнялись с помощью программного обеспечения Statistica 13 и SYSTAT 12.
Все измерения производились с помощью стереомикроскопа Zeiss Stereomicroscope SteREO Discovery V.12. Затем для составления морфологической классификационной таблицы полученные изображения зубов на микроскопе обрабатывались в программе Photoshop CS6.
Морфотипическая изменчивость рассматривалась на основании изучения вариаций строения лабиальной и лингвальной сторон передней непарной петли и особенностей строения лингвального входящего угла 4, которое оценивалось на основании промера L3.
Было выделено четыре типа строения лингвального входящего угла 4, обозначенных римскими цифрами (I – II – III – V) (рис. 3).
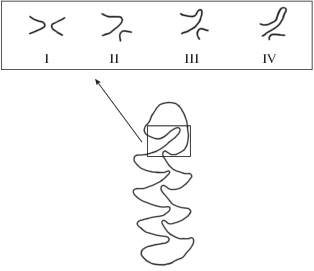
I тип. Вершины лингвального входящего угла 4 и лабиального входящего угла 3 расположены на одном уровне один напротив другого. L3 = 0.
II, III и IV типы различаются по глубине лингвального входящего угла 4.
К типу II отнесены зубы, имеющие значения L3 в пределах от 0.06 до 0.20 мм.
К типу III при значениях L3 от 0.21 до 0.38 мм.
К IV типу от 0.39 мм и выше.
Формы лингвальной и лабиальной сторон были разделены на несколько типов (рис. 4).
Рис. 4.
Форма лингвальной (слева) и лабиальной (справа) сторон головки антероконида. Пояснения в тексте.

Форма лингвальной стороны
Тип A. Лингвальная сторона без выемки.
Тип B. Лингвальная сторона имеет выемку с прямым внутренним краем.
Тип С. Лингвальная сторона имеет выемку под прямым или тупым углом.
Тип D. Лингвальная сторона имеет выемку под углом менее 90° или входящий угол.
Тип E. Лингвальная сторона имеет две выемки.
Форма лабиальной стороны
Тип a. Лабиальная сторона без выемки.
Тип b. Лабиальная сторона имеет маленькую выемку в нижней части.
Тип c. Лабиальная сторона имеет широкую выемку, захватывающую всю срединную часть, или выемку в верхней части.
Тип d. Лабиальная сторона имеет небольшой выступ в срединной части.
В итоге, описав жевательную поверхность, мы получаем комбинацию из трех состояний признаков, например, морфотип IIIBa.
Далее анализ изменчивости проводился с помощью комбинативного метода (Васильева, 1978; Большаков и др., 1980; Круковер, 1989; Поздняков, 1993).
На основе выведенных морфотипов (табл. 1) была построена матрица данных (табл. 2), которая анализировалась с помощью программы PAST – Palaeontological Statistics (версия 4.05) (Hammer, 2021) с применением метода невзвешенного попарного среднего (UPGMA), коэффициента корреляции Пирсона и бутстреп-анализа в 1000 повторениях.

Табл. 2.
Морфологические характеристики выборок m1 восточных полевок Приморья
| Морфо типы | Выборки | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| IBa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.94 | 0 | 0 | 0 | 0 | 0 |
| ICa | 7.69 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2.83 | 0 | 0 | 0 | 0 | 0 |
| ICc | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 |
| IDa | 2.56 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7.55 | 25 | 0 | 0 | 0 | 0 |
| IDb | 2.56 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.89 | 0 | 0 | 0 | 0 | 0 |
| IICa | 18 | 7.41 | 22.2 | 14.8 | 2.56 | 6.25 | 0 | 2.59 | 25.5 | 25 | 0 | 10 | 3.85 | 3.08 |
| IICb | 5.13 | 11.1 | 7.41 | 7.41 | 0 | 0 | 0 | 0.86 | 2.83 | 0 | 0 | 10 | 0 | 3.08 |
| IICc | 0 | 3.7 | 3.7 | 0 | 0 | 4.17 | 0 | 0 | 1.89 | 25 | 0 | 0 | 7.69 | 4.61 |
| IIDa | 2.56 | 3.7 | 0 | 3.7 | 0.85 | 0 | 0 | 0 | 2.83 | 0 | 0 | 0 | 0 | 3.08 |
| IIDb | 0 | 7.41 | 0 | 3.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 0 | 3.08 |
| IIDc | 0 | 0 | 0 | 0 | 0.85 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 1.54 |
| IIEc | 2.56 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 |
| IIIAa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.72 | 1.89 | 0 | 0 | 0 | 0 | 0 |
| IIIAb | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.86 | 0 | 0 | 0 | 0 | 0 | 0 |
| IIIBa | 2.56 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.94 | 0 | 0 | 0 | 0 | 0 |
| IIIBb | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3.84 | 0 |
| IIICa | 23.1 | 22.2 | 22.2 | 14.8 | 38.5 | 35.4 | 70 | 34.5 | 10.4 | 13 | 50 | 30 | 34.6 | 33.85 |
| IIICb | 15.4 | 11.1 | 29.6 | 14.8 | 14.5 | 6.25 | 10 | 2.59 | 0 | 0 | 10 | 0 | 0 | 7.69 |
| IIICc | 0 | 7.41 | 0 | 3.7 | 12 | 14.6 | 10 | 0 | 0.94 | 0 | 0 | 0 | 0 | 1.54 |
| IIICd | 0 | 0 | 0 | 3.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3.08 |
| IIIDa | 10.3 | 11.1 | 0 | 0 | 12 | 10.4 | 10 | 5.17 | 3.77 | 13 | 0 | 0 | 30.8 | 23.07 |
| IIIDb | 5.13 | 7.41 | 3.7 | 25.9 | 5.98 | 2.08 | 0 | 0.86 | 0 | 0 | 10 | 0 | 3.85 | 6.15 |
| IIIDc | 0 | 3.7 | 3.7 | 0 | 2.56 | 0 | 0 | 0.86 | 0 | 0 | 20 | 0 | 7.69 | 1.54 |
| IIIEa | 0 | 0 | 3.7 | 0 | 0 | 2.08 | 0 | 1.72 | 0 | 0 | 0 | 0 | 0 | 1.54 |
| IIIEb | 0 | 0 | 3.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.54 |
| IIIEc | 0 | 0 | 0 | 0 | 0 | 4.17 | 0 | 0 | 0 | 0 | 0 | 0 | 3.85 | 0 |
| IVAa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3.45 | 9.43 | 0 | 0 | 0 | 0 | 0 |
| IVBa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.72 | 4.72 | 0 | 0 | 0 | 0 | 0 |
| IVCa | 0 | 0 | 0 | 0 | 6.84 | 4.17 | 0 | 17.2 | 10.4 | 0 | 0 | 0 | 3.85 | 0 |
| IVCb | 0 | 0 | 0 | 3.7 | 0 | 0 | 0 | 0 | 0.94 | 0 | 0 | 0 | 0 | 0 |
| IVCc | 0 | 0 | 0 | 0 | 1.71 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| IVDa | 0 | 0 | 0 | 0 | 1.71 | 4.17 | 0 | 23.3 | 8.49 | 0 | 10 | 0 | 0 | 0 |
| IVDb | 2.56 | 0 | 0 | 0 | 0 | 2.08 | 0 | 0.86 | 1.89 | 0 | 0 | 0 | 0 | 1.54 |
| IVDc | 0 | 0 | 0 | 3.7 | 0 | 4.17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| IVDd | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.86 | 0 | 0 | 0 | 0 | 0 | 0 |
| IVEa | 0 | 3.7 | 0 | 0 | 0 | 0 | 0 | 0.86 | 0 | 0 | 0 | 0 | 0 | 0 |
Примечания. 1 – позднеплейстоценовая выборка 1, 2 – позднеплейстоценовая выборка 2, 3 – среднеголоценовая выборка, 4 – позднеголоценовая выборка, 5 – окрестности пос. Хасан, 6 – побережье бухты Астафьева, 7 – о-в Фальшивый, 8 – о-в Веры, 9 – острова Дурново, Де-Ливрона, 10 – о-в Гильдебрандта, 11 – о-в Пелис, 12 – о-в Рикорда, 13 – о-в Русский, 14 – о-в Путятина.
РЕЗУЛЬТАТЫ
Дискретность сочетаний отдельных признаков позволила построить комбинативную морфологическую классификационную таблицу и проводить многомерную классификацию отдельных зубов по определенным морфотипам (табл. 1). Классификационная таблица представляет собой прямоугольную сетку, в центре которой показаны типы строения лингвального входящего угла 4. Соответственно все морфотипы левого верхнего угла таблицы имеют I тип строения этого угла, правого верхнего – II тип, правого нижнего – III тип и левого нижнего – IV тип. В вертикальном столбце посередине таблицы показаны варианты строения лингвальной стороны головки антероконида. В горизонтальной строке посередине таблицы – варианты строения лабиальной стороны. Все варианты строения расположены в порядке возрастания сложности: от I до IV, от A до E, от a до d. В полученные от пересечения столбцов и строк ячейки помещены графические схемы рисунка передней непарной петли m1. Реальное количество морфотипов меньше теоретически ожидаемого, поэтому не все ячейки заполнены. Возможно, какие-то ячейки будут заполнены в будущем, а какие-то останутся не заполненными в связи с воздействием природных факторов, ограничивающих изменчивость.
Сочетание выделенных признаков дает 36 морфотипов строения непарной передней петли антероконида, каждый из которых имеет буквенно-цифровое обозначение. Дискриминантный анализ подтвердил правильность выделения четырех типов строения лингвального входящего угла 4 (доля правильной классификации методом “складного ножа” (jacknife) составила 97% (табл. 3, рис. 5)). Для оценки полезности канонических дискриминантных функций были рассчитаны основные статистические показатели (табл. 4). Исходя из данных таблицы, можно отметить хорошую дискриминацию групп. Первые две дискриминантные функции имеют статистическую значимость. Большое значение канонической корреляции (R = 0.94) первой дискриминантной функции свидетельствует о ее лучшей разделительной способности. В свою очередь статистическая значимость третьей дискриминантной функции превышает пороговое значение (p = 0.27), из-за чего различие типов по этой функции не может подлежать содержательной интерпретации. Наибольший вклад в разделение морфотипов внесли промеры L1, L3 и L5. При этом промер L3, который отвечает за измерение лингвального входящего угла 4, оказался наиболее важным и ценным (Wilk`s Lambda = 0.21, Partial Lambda = 0.47). Необходимо отметить, что классификационная таблица была построена на современном материале. Идентификацию морфотипов m1 ископаемых полевок проводили на основе полученной ранее таблицы. При этом не предусмотренных в таблице новых морфотипов обнаружено не было.
Табл. 3.
Оценка корректности классификации с использованием метода “складного ножа” (jacknife)
| Группа | 1 | 2 | 3 | 4 | Точность классификации, % |
|---|---|---|---|---|---|
| 1 | 17 | 0 | 0 | 0 | 100 |
| 2 | 1 | 61 | 0 | 0 | 98 |
| 3 | 0 | 3 | 184 | 6 | 95 |
| 4 | 0 | 0 | 0 | 104 | 100 |
| Всего | 18 | 64 | 184 | 110 | 97 |
Рис. 5.
Распределение исследованных первых нижних коренных зубов восточной полевки в пространстве первой (Root 1) и второй (Root 2) дискриминантных функций по результатам измерений m1.
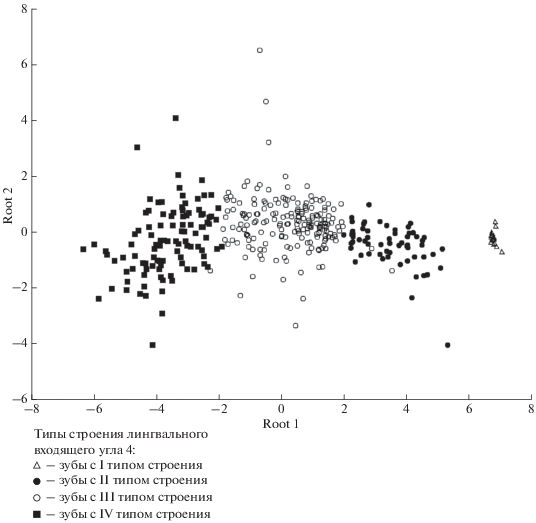
Табл. 4.
Статистические результаты для дискриминантных канонических функций
| Дискриминантная функция | Собственные значения | Каноническая корреляция R | Лямбда Уилка |
χ2 | Уровень значимости |
|---|---|---|---|---|---|
| 1 | 7.801516 | 0.942 | 0.1009 | 848.78 | p < 0.0001 |
| 2 | 0.110835 | 0.316 | 0.8878 | 44.06 | p = 0.000003 |
| 3 | 0.014066 | 0.118 | 0.9861 | 5.17 | p = 0.2705 |
Основным и наиболее часто встречающимся морфотипом в двух изученных выборках с материка является морфотип IIICa. Для него характерны средняя глубина лингвального входящего угла 4 (промер L3 от 0.16 до 0.38), широкая неглубокая выемка на лингвальной стороне передней непарной петли и отсутствие выемки на лабиальной стороне. В выборке из окрестностей пос. Хасан обнаружено 38% зубов с таким морфотипом, на побережье бухты Астафьева – 35%. Из остальных 11 морфотипов относительно часто встречающимися в Хасанской выборке были морфотипы IIICb, IIICc и IIIDa (14, 12 и 12% соответственно). В Астафьевской выборке 13 морфотипов, к основным, кроме морфотипа IIICa, можно отнести IIICс и IIIDa (около 15 и 10% соответственно). Один из восьми редких или резервных (Малеева, 1976) морфотипов обнаружен только в Хасанской выборке, другие распространены шире. В Астафьевской выборке один из редких морфотипов обнаружен еще в позднеголоценовой выборке, другой – на о-ве Русский, остальные 8 редких морфотипов распространены шире (табл. 2). Необходимо отметить, что в материковых выборках нет морфотипов из левого верхнего угла табл. 1, т.е. морфотипов с I типом строения.
Имеющийся в нашем распоряжении ископаемый материал из отложений пещеры Медвежий Клык можно разделить на 4 выборки. Позднеплейстоценовая выборка 1 – материал из 12 и 11 слоев, возраст отложений около 40–35 тыс. лет (Панасенко, Тиунов, 2010; Omelko et al., 2020). Позднеплейстоценовая выборка 2 – материал из наклонных слоев 6–9, возраст около 14–11 тыс. лет. Среднеголоценовая выборка – материал из слоя 5, возраст около 6 тыс. лет. Позднеголоценовая выборка – материал из слоев 3 и 4, возраст около 3–4 тыс. лет. В обеих позднеплейстоценовых выборках основным наиболее часто встречающимся морфотипом является морфотип IIICa. В среднеголоценовой выборке – морфотип IIICa, в позднеголоценовой выборке – морфотип IIIDb. В позднеплейстоценовой выборке 1 встречаемость более 10% отмечена также для морфотипов IICa, IIICb и IIIDa, в позднеплейстоценовой выборке 2 – IICb, IIICb и IIIDa, в среднеголоценовой – IICa и IIICa, в позднеголоценовой – IICa, IIICa и IIICb (рис. 6). Сильных колебаний количества морфотипов в ископаемых выборках не наблюдается. В позднеплейстоценовой выборке 1 зарегистрировано 13 морфотипов, в позднеплейстоценовой выборке 2–12 морфотипов, в среднеголоценовой – 9, в позднеголоценовой – 11.
Рис. 6.
Соотношение (в %) морфотипов m1 A. fortis в плейстоцен-голоценовом времени: A – позднеплейстоценовая выборка 1, B – позднеплейстоценовая выборка 2, C – среднеголоценовая, D – позднеголоценовая. Штриховкой показаны варианты морфотипов.

В большинстве выборок из изолированных островных популяций залива Петра Великого, кроме выборок из популяций островов Дурново и Де-Ливрона, наибольшей встречаемостью также характеризуется морфотип IIICa. На островах Дурново и Де-Ливрона основным наиболее часто встречающимся морфотипом является морфотип IICa (~26%). Вторым по встречаемости на этих островах является морфотип IIICa (~10%), на третьем месте морфотип IVCa (рис. 7, табл. 1). На этих островах обнаружено 19 морфотипов. Это наибольшее количество среди всех рассмотренных выборок. Из 16 редко встречающихся здесь морфотипов два морфотипа отмечены только на этих островах. Два морфотипа встречаются не только на этих островах, но еще на о-ве Веры, три морфотипа – в позднеплейстоценовой выборке 1 и один морфотип встречается в позднеголоценовой выборке.
Рис. 7.
Соотношение (в %) морфотипов m1 A. fortis в современных выборках: A – окрестности пос. Хасан; B – побережье бухты Астафьева; C – о-в Веры; D – острова Де-Ливрона, Дурново; E – о-в Русский; F – о-в Путятина. Обозначения морфотипов те же, что на рис. 6.
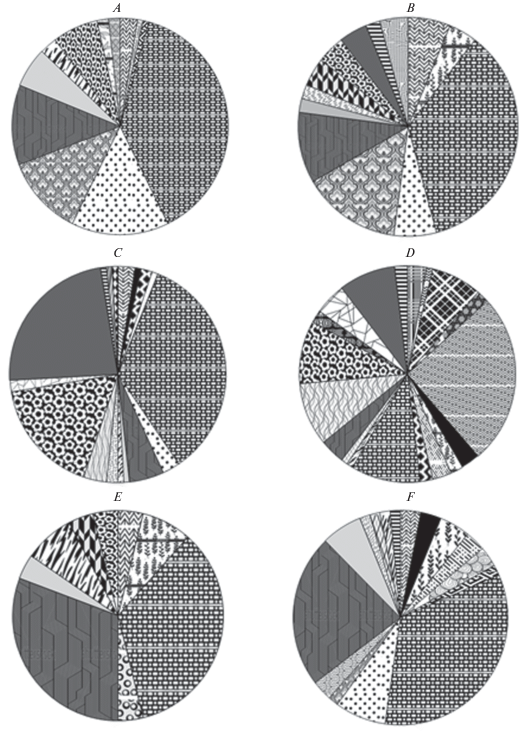
На о-ве Веры зарегистрировано 17 морфотипов строения непарной передней петли антероконида m1. Наиболее часто встречающимся морфотипом является морфотип IIICa (~34%), на втором месте по встречаемости находится морфотип IVDa (~23%), на третьем – IVCa (~17%). Из редких морфотипов только на о-ве Веры обнаружены 2 морфотипа, 2 морфотипа обнаружены на о-ве Веры и в позднеплейстоценовой выборке 1.
В остальных изученных выборках из островных изолированных популяций наиболее часто встречающимся морфотипом является морфотип IIICa. К часто встречающимся в этих выборках можно также отнести морфотипы IIICb, IIICc, IIIDa и IIDb. По одному редкому морфотипу, встречающемуся только в одной из изученных выборок, обнаружено на островах Рикорда и Русский.
ОБСУЖДЕНИЕ
Если не рассматривать выборки, количество исследованных зубов в которых явно не достаточно для анализа (о-в Гильдебранта 8 экз., о-в Фальшивый 10 экз., о-в Пелис 10 экз., о-в Рикорда 10 экз.), то, как показали наши исследования, во всех ископаемых, современных материковых и островных выборках m1 восточной полевки преобладающими по встречаемости являются небольшие группы, включающие от 2 до 4 морфотипов и составляющие от 55 до 77%. Преобладание небольшой группы морфотипов в выборках отмечено и для многих других видов полевок (Малеева, 1976; Большаков и др., 1980). Смена лидирующего по встречаемости морфотипа зарегистрирована только в среднеголоценовой, позднеголоценовой выборках и на островах Дурново и Де-Ливрона. Превалирующий по встречаемости в позднем плейстоцене морфотип IIICa сменился на морфотип IIICb с более сложно устроенной лабиальной стороной, имеющей выемку в нижней части. Необходимо отметить, что если в среднеголоценовой выборке один из основных морфотипов сменил другой, то в позднеголоценовой выборке на первое место по встречаемости выходит морфотип IIIDb, находившийся ранее в категории редких или резервных морфотипов. В настоящее время в материковых выборках снова превалирует морфотип IIICa. Несмотря на то, что во временно́й промежуток между выборками позднеплейстоценовая 1 и позднеплейстоценовая 2 на рубеже 18–20 тыс. лет происходило максимальное похолодание климата, смены лидирующего морфотипа не произошло.
Интересно, что, судя по шести рисункам жевательной поверхности первых нижних коренных зубов Alexandromys fortis, извлеченных из позднеплейстоценовых пещерных отложений на о-ве Мияко (Япония) (Kaneko, Hasegawa, 1995), на острове в тот период встречались следующие морфотипы: ICa (1 зуб), IICa (4 зуба), IICc (1 зуб). Морфотип ICa это редкий морфотип, отмеченный в позднеплейстоценовой выборке 1 и в выборке с островов Дурново и Де-Ливрона. Морфотип IICa встречается во всех ископаемых выборках, на материке и островах, входит в число основных в позднеплейстоценовой выборке 1 и лидирует по встречаемости на островах Дурново и Де-Ливрона. Широко распространенный редкий морфотип IICc отмечен в позднеплейстоценовой выборке 2, в среднеголоценовой выборке, на материке и островах. Считается, что восточная полевка мигрировала с континента на о-в Мияко в среднем плейстоцене. В современной фауне острова этот вид не встречается (Kaneko, Hasegawa, 1995; Kawamura, 2014).
Повышенное морфотипическое разнообразие на островах Путятина, Дурново и Де-Ливрона, Веры характерно и для многих других островных популяций полевок и связано с высвобождением скрытой фенотипической изменчивостью из-за инбридинга (Markova et al., 2020).
С другой стороны, постоянное прохождение “бутылочного горлышка” является нормой для восточной полевки. Так, в период депрессии, которая может продолжаться несколько лет, абсолютная численность полевок на некоторых островах измеряется несколькими десятками особей (Катин, 1989).
Среди изолированных островных популяций восточной полевки наиболее интересный набор морфотипов наблюдается на островах Дурново, Де-Ливрона и на о-ве Веры. Лидирующий по встречаемости на островах Дурново, Де-Ливрона морфотип IICa входит в число основных в позднеплейстоценовой выборке 1, в среднеголоценовой и позднеголоценовой выборках. В остальных выборках он относится к категории редких.
Острова Большой Пелис, Гильдебрандта, Дурново, Де-Ливрона входят в состав архипелага Римский-Корсаков. Отделение островов этого архипелага от материка произошло ранее, чем отделение всех остальных островов залива Петра Великого – в конце позднего плейстоцена и начале голоцена (Велижанин, 1976). Вполне вероятно, что особенности морфотипической структуры популяции островов Дурново и Де-Ливрона связаны с эффектом основателя. Необходимо отметить и наличие в этой выборке значительного количества как “простых”, так и “сложных” морфотипов. Наличие “простых” морфотипов характерно еще только для позднеплейстоценовой выборки 1. Количество “сложных” морфотипов с глубоким лингвальным входящим углом 4 на островах Дурново и Де-Ливрона достигает 35.8%, на о-ве Веры – 47.4%. В других исследованных выборках встречи таких морфотипов единичны. Повышенную встречаемость в популяции полевок особей со сложной структурой жевательной поверхности зубов связывают с более грубыми кормами (Сорокина, 2011), а условия существования восточных полевок на каменистом грунте маленьких морских островов, где мало сочных растений, значительно отличаются от условий существования на материке (Чугунов, Катин, 1984).
Если острова Дурново и Де-Ливрона находятся в изоляции не менее 10 тысяч лет (а возможно и дольше, поскольку в период отступления моря это были вершины холмов, изолированные друг от друга и от материка другими ландшафтами), то о-в Веры, отделенный от материка узким неглубоким проливом шириной 480 м, отсоединился от континента гораздо позже. Восточная полевка является единственным видом грызунов, обитающим на островах Дурново, Де-Ливрона и Гильдебрандта. На о-ве Веры, кроме восточной полевки, встречается восточноазиатская мышь Apodemus peninsulae Thomas 1907 (Чугунов, Катин, 1984). Однако у этих островов есть общие черты, отличающие их от других исследованных островов. Острова Дурново, Де-Ливрона и Веры являются самыми маленькими среди обследованных. Последняя территориальная связь у них была с самой южной частью Приморья (Хасанский р-н). Особи из выборки позднеплейстоценовой 1 существовали в период климатического оптимума среднего вюрма (43–33 тыс. лет) по интенсивности повышения температур и режиму увлажнения сходному с климатом среднего голоцена (Короткий и др., 1996). В периоды климатических оптимумов среднего вюрма и среднего голоцена происходило значительное смещение как зон растительности, так и фауны к северу относительно их современного положения (Короткий и др., 1988). Исходя из этого, можно предположить, что в качестве основателей популяций на этих островах принимали участие и южные формы восточной полевки, отличающиеся по гаплотипам (Gao et al., 2017). Гаплотипы, сходные с южными формами, распространенными в Китае, обнаружены как среди ископаемых полевок из отложений пещеры Медвежий Клык, так и в популяциях полевок с островов архипелага Римский-Корсаков (Haring et al., 2015; Шереметьева, 2020).
Определенную роль в различии материковых и островных популяций играло, вероятно, и отличие внутрипопуляционных эволюционных процессов, по-разному проходящих на этих территориях (MacArthur, Wilson, 1967). Ранее было показано, что темпы морфологической эволюции в островных популяциях млекопитающих значительно выше, чем в материковых популяциях. После начального периода ускоренных изменений (в промежутке от 21 года до 20 000 лет) скорость эволюции островных видов замедляется, приближаясь к темпам на материке (Millien, 2006). Причем млекопитающие на более мелких островах развиваются быстрее, чем млекопитающие на более крупных островах (Millien, 2011).
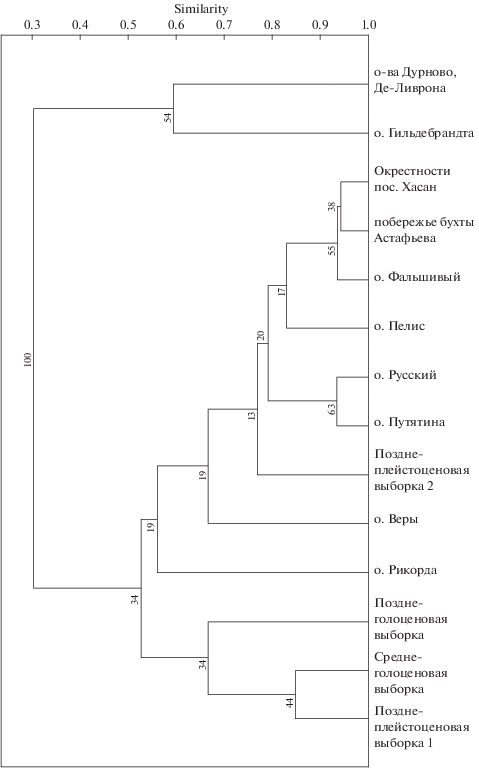
Выборки с крупных островов Русский и Путятина, отделение которых от материка произошло около 8.5–7 тыс. лет назад, схожи между собой и близки по составу морфотипов к материку (рис. 8). Условия существования восточной полевки на этих островах не так сильно отличаются от условий существования на материке.
ВЫВОДЫ
У Alexandromys fortis, как и у большинства видов полевок (Малеева, 1976), во всех выборках преобладает небольшая группа морфотипов (от 2 до 4).
Отмечено вре́менное усложнение преобладающего типа жевательной поверхности m1 восточной полевки в голоцене. В настоящее время на континенте наибольшая встречаемость отмечена у морфотипа, который превалировал в позднем плейстоцене. Изменения в частоте встречаемости морфотипов, происходившие в голоцене, могут быть связаны с возобновившимся распространением, слиянием разрозненных популяций и восстановлением численности восточной полевки (Алексеева, Голенищев, 1986) после падения ее численности в период максимального похолодания климата.
Особенности морфотипического состава m1 изолированных популяций восточной полевки на островах Дурново, Де-Ливрона и Веры обусловлены как фенотипическими и генотипическими особенностями основателей островных популяций, так и своеобразием новых условий обитания, инициировавшими высокий темп эволюции, высвобождение скрытого разнообразия и увеличение количества редких морфотипов.
Список литературы
Алексеева Э.А., Голенищев Ф.Н., 1986. Ископаемые остатки серых полевок рода Microtus из южного Приморья (пещера “Близнец”) // Труды Зоологического института АН СССР. Т. 156. С. 134–142.
Большаков В.Н., Васильева И.А., Малеева А.Г., 1980. Морфотипическая изменчивость зубов полевок. М.: Наука. 140 с.
Бородин А.В., Маркова Е.А., 2014. Ключи для определения современных и плейстоценовых полевок (Arvicolinae, Rodentia) Урала и Западной Сибири по одонтологическим признакам // Зоологический журнал. Т. 93. С. 1128–1139.
Васильева И.А., 1978. Изучение изменчивости жевательной поверхности М3 у некоторых представителей рода Microtus Schrank при их гибридизации // Физиологическая и популяционная экология животных. Вып. 5 (7). Саратов: изд-во Саратовского ун-та. С. 96–101.
Велижанин А.Г., 1976. Время изоляции материковых островов северной части Тихого океана // Доклады Академии наук СССР. Т. 231. № 1. С. 205–207.
Войта Л.Л., Голенищев Ф.Н., Тиунов М.П., 2019. Дальневосточные полевки Alexandromys (Rodencia: Cricetidae) из позднеплейстоценовых и голоценовых отложений пещеры Медвежий клык Приморского края России // Труды Зоологического института РАН. Т. 323. № 3. С. 313–346.
Голенищев Ф.Н., Войта Л.Л., Моролдоев И.В., Абрамсон Н.И., Петрова Т.В., Картавцева И.В., 2018. Новые находки муйской полевки (Rodencia: Cricetidae: Alexandromys mujanensis) в Забайкалье // Труды Зоологического института РАН. Т. 322. № 3. С. 357–384.
Гусев А.Е., Тиунов М.П., 2021. Внутривидовая изменчивость формы строения третьего нижнего предкоренного зуба северной пищухи (Ochotona hyperborea (Pallas 1811)) // Зоологический журнал. Т. 100. № 1. С. 104–114.
Катин И.О., 1989. Динамика популяций дальневосточной полевки в условиях островной изоляции // Териологические исследования на юге дальнего Востока. Владивосток: ДВО РАН СССР. С. 89–99.
Ковалева В.Ю., Поздняков А.А., Литвинов Ю.Н., Ефимов В.М., 2021. Флуктуирующая асимметрия и морфогенетические корреляции рисунков жевательной поверхности m1 серых полевок (Rodentia, Arvicolinae) // Зоологический журнал. Т. 100. № 4. С. 434–448.
Короткий А.М., Плетнев С.П., Пушкарь В.С., Гребенникова Т.А., Разжигаева Н.Г. и др., 1988. Развитие природной среды юга Дальнего Востока (поздний плейстоцен-голоцен) / Отв. ред. Краснов Е.В. АН СССР Дальневосточное отделение, Тихоокеанский институт географии. М.: Наука. 240 с.
Короткий А.М., Гребенникова Т.А., Пушкарь В.С., Разжигаева Н.Г., Волков В.Г. и др., 1996. Климатические смены на территории юга Дальнего Востока в позднем кайнозое (миоцен-плиоцен). Владивосток: Дальнаука. 58 с.
Костенко В.А., Алленова Т.В., 1989. Внутривидовая дифференциация полевки-экономки на Дальнем Востоке и история становления ареалов ее подвидовых форм // Териологические исследования на юге Дальнего Востока. Владивосток: ДВО АН СССР. С. 4–25.
Круковер А.А., 1989. Строение и морфотипическая изменчивость жевательной поверхности зубов некорнезубых полевок. Новосибирск. 39 с. (Препр. ИГиГ СО АН СССР; № 12).
Лисовский А.А., Кадетова А.А., Оболенская Е.В., 2018. Морфологическое определение видов восточноазиатских серых полевок Alexandromys (Rodentia, Cricetidae) России и сопредельных территорий // Зоологический журнал. Т. 97. № 1. С. 101–113.
Малеева А.Г., 1976. Об изменчивости зубов у полевок (Microtinae) // Труды Зоологического института АН СССР. Т. 66. С. 48–57.
Мейер М.Н., 1978. Систематика и внутривидовая изменчивость серых полевок Дальнего Востока (Rodentia, Cricetidae) // Труды Зоологического института РАН. Т. 75. С. 3–62.
Наследов А.Д., 2004. Математические методы психологического исследования. Санкт-Петербург: Речь. 392 с.
Панасенко В.Е., Тиунов М.П., 2010. Население мелких млекопитающих (Mammalia: Eulipotyphla, Rodentia, Lagomorpha) на южном Сихотэ-Алине в позднем плейстоцене и голоцене // Вестник ДВО РАН. № 4. С. 60–67.
Поздняков А.А., 1993. Морфотипическая изменчивость жевательной поверхности коренных зубов серых полевок группы “maximowiczi” (Rodentia, Arvicolidae, Microtus): опыт количественного статистического анализа // Зоологический журнал. Т. 72. Вып. 11. С. 114–125.
Поздняков А.А., Литвинов Ю.Н., 1994. Экогеографическая интерпретация морфотипической изменчивости жевательной поверхности коренных зубов полевки-экономки Microtus oeconomus (Rodentia, Arvicolidae) // Зоологический журнал. Т. 73. Вып. 2. С. 151–157.
Поздняков А.А., 2010. Морфологическое разнообразие: характеристика, структура, анализ // Труды института систематики и экологии животных СО РАН. Т. 46. С. 133–157.
Сорокина Н.В., 2011. Морфотипическая изменчивость структуры жевательной поверхности зуба М3 красной полевки (Clethrionomys rutilus Pallas, 1779) (Rodentia, Cricetidae) северной и средней тайги Тюменской области // Известия Самарского научного центра РАН. Т. 13. № 1 (5). С. 1154–1157.
Фадеева Т.В., 2003. Сравнительная морфологическая характеристика первых нижнекоренных зубов ископаемых и рецентных полевок-экономок Урала // Систематика, филогения и палеонтология мелких млекопитающих. Санкт-Петербург: ЗИН РАН. С. 221–224.
Фадеева Т.В., 2005. Пространственно-временная морфологическая изменчивость выборок первых нижнекоренных зубов полевок-экономок // Труды Зоологического института РАН. Т. 306. С. 180–201.
Чугунов Ю.Д., Катин И.О., 1984. Численность и распределение по биотопам грызунов на островах Дальневосточного государственного морского заповедника // Животный мир Дальневосточного морского заповедника. Владивосток: ДВНЦ АН СССР. С. 107–121.
Шереметьева И.Н., 2003. Морфологическая изменчивость дальневосточной полевки (Microtus fortis, Rodencia, Cricetidae), с островов Залива Петра Великого (Японское море) // Зоологический журнал. Т. 82. № 9. С. 1138–1143.
Шереметьева И.Н., 2020. Сравнение гаплотипов мтДНК дальневосточной полевки и современных гаплотипов у островных популяций // Генетика. Т. 56. № 7. С. 848–854.
Chelomina G.N., Sheremetyeva I.N., 2007. Genetic variation of Microtus fortis pelliceus in mainland and island populations of the Russian Far East based on random amplified polymorphic DNA markers // Mammal Study. V. 32. P. 63–74.
Gao J., Yue L., Jiang X., Ni L., Ashraf M.A. et al., 2017. Phylogeographic Patterns of Microtus fortis (Arvicolinae: Rodentia) in China Based on Mitochondrial DNA Sequences. [Электронный ресурс] Режим доступа:
http://researcherslinks.com/current-issues/Phylogeographic-Patterns-Microtus-fortis-Arvicolinae-Rodentia-China-Based-%20Mitochondrial-DNA-Sequences/20/1/496/html
Дата обновления: 12.04.2021.
Golenishchev F., Malikov V., Petrova T., Bodrov S., Abramson N., 2019. Toward assembling a taxonomic, puzzle Case study of Iranian gray voles of the subgenus Microtus (Rodentia, Cricetidae) // Mammalian Biology. V. 94. P. 98–105.
Hammer Ø., 2021. Paleontological Statistics. Version 4.0. Reference Manual [Электронный ресурс] Режим доступа: https://www.nhm.uio.no/english/research/infrastructure/past/downloads/past4manual.pdf Дата обновления: 12.04.2021.
Haring E., Voyta L.L., Daubl B., Tiunov M.P., 2015. Comparison of genetic and morphological characters in fossil teeth of grey voles from the Russian Far East (Rodentia: Cricetidae: Alexandromys) // Mammalian Biology. V. 80. P. 496–504.
Kaneko Y., Hasegawa Y., 1995. Some fossil arvicolid rodents from the Pinza-Abu Cave, Miyako Island, the Ryukyu Islands, Japan // Bull. Biogeogr. Soc Japan. V. 50. № 1. P. 23–37.
Kawamura Y., 2014. Research on Quaternary mammal faunas in Japan and adjacent East areas: a review of my research // The Quaternary Research (Daiyonki Kenkyu). V. 53. № 3. P. 119–142.
MacArthur R.H., Wilson E.O., 1967. The theory of island biogeography. Princeton-New Jersey: Princeton University Press. 203 p.
Markova E., Bobretsov A., Borodin A., Rakitin S., Sibiriyakov P. et al., 2020. The effects of population bottlenecks on dental phenotype in extant arvicoline rodents: Implications for studies of the quaternary fossil record // Quaternary Science Reviews. V. 228. P. 1–20.
Millien V., 2006. Morphological evolution is accelerated among island mammals // PLoS Biology. V. 4. № 4. P. 1863–1868.
Millien V., 2011. Mammals evolve faster on smaller islands // Evolution. V. 65. № 7. P. 1935–1944.
Nadachowski A., 1982. Late Quaternary rodents of Poland with special reference to morphotype dentinion analysis of voles. Warszawa-Krakow: Panstwowe wydawnictwo naukowe. 110 p.
Omelko V.E., Kuzmin Y.V., Tiunov M.P., Voyta L.L, Burr G.S., 2020. Late Pleistocene and Holocene small mammal (Lipotyphla, Rodentia, Lagomorpha) remains from Medvezhyi Klyk Cave in the Southern Russian Far East // Proceedings of the Zoological Institute RAS. V. 324. № 1. P. 124–145.
Semken H.A.Jr., Wallace S.C., 2002. Key to arvicoline (“microtine” rodent) and arvicoline-like first lower molars recovered from late Wisconsinan and Holocene archaeological and paleontological sites of eastern North America // Journal of Archaeological Science. V. 29. P. 23–31.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал