

Зоологический журнал, 2022, T. 101, № 2, стр. 165-172
Пространственно-типологическая структура и организация летнего населения птиц Кыргызстана
Ю. С. Равкин a, *, Э. Давранов a, И. П. Лебяжинская b, Э. Ш. Касыбеков c, А. Б. Курманкулов c, Э. Э. Шукуров c, Н. В. Тротченко d, А. А. Жусупбаева c, С. М. Цыбулин a, М. И. Лялина a
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
b Заповедник “Приволжская лесостепь”
440031 Пенза, Россия
c Экологическое движение Кыргызской Республики “Алейне”
720071 Бишкек, Кыргызская Республика
d Институт биологии Национальной академии наук Кыргызской Республики
720071 Бишкек, Кыргызская Республика
* E-mail: zm@eco.nsc.ru
Поступила в редакцию 11.11.2020
После доработки 20.04.2021
Принята к публикации 21.04.2021
- EDN: YIGNPZ
- DOI: 10.31857/S0044513421120102
Аннотация
Представлены результаты многомерного анализа данных, собранных в летний период. Суммарная протяженность учетных маршрутов около 1700 км. Сопоставление с аналогичным исследованием, выполненным ранее по вдвое меньшему объему исходных данных, позволило оценить репрезентативность собранных материалов и уточнить представления о пространственной неоднородности населения птиц Кыргызстана. Основными факторами среды, определяющими изменчивость орнитокомплексов, служат тип растительности, облесенность и провинциальные различия рельефа и климата. С ними в целом связано 38% дисперсии матрицы сходства населения птиц. Построенные по обоим массивам данных структурные графы и иерархические классификации демонстрируют высокую степень совпадения. Различия сводятся к чуть большей оценке силы связи изменчивости населения птиц с уровнем высот местности – в первом варианте расчетов, при меньшем объеме данных, и степенью облесенности – во втором. Информативность обеих классификаций, оцененная суммарной величиной объясненной дисперсии матрицы сходства анализируемых сообществ, различается незначительно. Показано, что экспертное разделение территории по населению птиц менее информативно, чем по результатам кластерного анализа.
Орнитофауна Кыргызстана изучена сравнительно полно (Янушевич и др., 1959, 1960; Янушевич, 1961). Современные знания о населении птиц этого своеобразного и весьма интересного в зоогеографическом отношении региона в общем недостаточны для широких обобщений. Тем не менее в начале XXI века были подведены предварительные итоги изучения орнитокомплексов Кыргызстана. Для этого использованы результаты количественных учетов птиц за май, июнь, июль в 159 местообитаниях (Шукуров и др., 2004, 2005). К 2020 г. общее количество изученных в летний период пространственных вариантов населения достигло 197. Это позволило повторить анализ собранных данных, но уже включая август. Мы сочли возможным проведение расчетов в среднем за весь летний период, поскольку пространственные тренды в орнитокомплексах как в первой, так и во второй половине лета, несмотря на сезонные изменения в жизни птиц, остаются почти неизменными (Равкин, 1984; Цыбулин, 2009).
МАТЕРИАЛ И МЕТОДИКА
Принципы и методы сбора и обработки материала подробно описаны ранее (Равкин, Ливанов, 2008). Птиц учитывали на маршрутах без ограничения ширины трансекта, с последующим пересчетом встреч по средневзвешенным (гармоническим) показателям дальности обнаружения. Для обобщения материалов, составления классификации населения птиц и выявления наиболее значимых структурообразующих факторов использован один из методов кластерного анализа – программа “Факторная классификация”. С ее помощью варианты сообществ объединяют в незаданное число групп так, чтобы дисперсия матрицы сходства орнитокомплексов была учтена такой классификацией в наибольшей степени. Оценка сходства проведена по коэффициенту Жаккара для количественных признаков. Иерархия типов и подтипов классификации соответствует очередности их формирования при доразбивке групп с помощью той же программы, а надтипов – по результатам повторной агрегации типов. Связь пространственной изменчивости орнитокомплексов и неоднородности местообитаний, а также общая информативность представлений оценены с помощью линейной качественной аппроксимации выделенными градациями факторов (вариант регрессионного анализа).
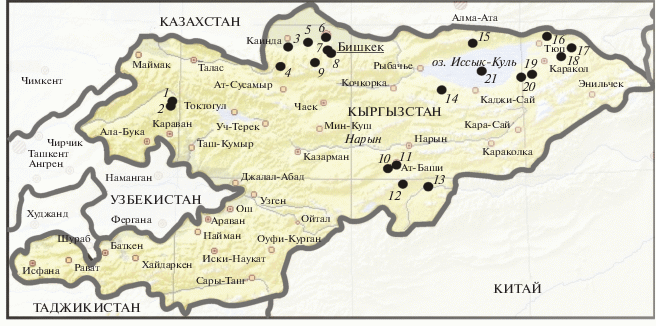
Материалы собраны за период с 15 мая по 31 августа в 1959–2006 гг. Суммарная протяженность учетных маршрутов составляет примерно 1700 км. Помимо авторов статьи, в сборе материала эпизодически принимали участие В.Н. Катаевский, С.П. Цопов, Б.К. Кумушалиев, М.Б. Старобинский, Г.Г. Воробьёв и А.Н. Остащенко. Кроме того, использованы данные Второва (1968) и Шукурова (1986). Места проведения учетных работ представлены на рис. 1. Названия видов птиц даны по: Иванов (1976), типов фауны – по: Штегман (1938) (с дополнениями).
Рис. 1.
Места и годы проведения учетов птиц: 1 – Лебяжинская И.П., 1982–1990; 2 – Лебяжинская И.П., 1988; 3, 4 – Давранов Э., 1992; 5 – Давранов Э., 2002–2003 и Катаевский В.Н., Цопов С.П., Кумушалиев Б.К., Старобинский М.Б., Воробьёв Г.Г., Остащенко А.Н., 1990; 6 – Тротченко Н.В., 2005–2006 и Жусупбаева А.А., 2004–2005; 7, 8 – Тротченко Н.В., 2005–2006; 9 – Давранов Э., 1992 и Тротченко Н.В., 2005–2006; 10 – Шукуров Э.Э., Цыбулин С.М., 2004 и Шукуров Э.Э., 2005; 11 – Шукуров Э.Э., 2005; 12, 13 – Курманкулов А.Б., 1989–1991; 14 – Шукуров Э.Д., 1986–1988 и Касыбеков Э.Ш., 1990; 15 – Касыбеков Э.Ш., 1987–1988 и Шукуров Э.Д., 1985–1988; 16 – Касыбеков Э.Ш., 1988 и Шукуров Э.Д., 1985–1988; 17, 18 – Касыбеков Э. Ш., 1987–1988 и 1986–1988; 19 – Второв П.П., 1962–1964; 20 – Касыбеков Э.Ш., 1987–1988 и Шукуров Э.Д., 1985–1988; 21 – Касыбеков Э.Ш., 1986–1988.
РЕЗУЛЬТАТЫ
Классификация орнитокомплексов
Орнитокомплексы Кыргызстана11:
I – естественных ландшафтов, садов и лесных посадок (кроме открытых околоводных)
1 – местообитаний очень низкой пригодности для птиц22 (нивальных; лидируют по обилию, % – альпийская галка 40, жемчужный горный вьюрок 28, краснобрюхая горихвостка 11, темнобрюхий улар и большая розовая чечевица по 6; плотность населения – 6 особей/км2; отмечено 15 видов / в том числе 2 фоновых; доля преобладающих по обилию типов фауны (10% и более) – 96% тибетского типа33);
2 – низкой степени пригодности местообитаний: высокогорных Аксай–Верхненарынской физико-географической провинции44, а также пустынных, полупустынных, опустыненных и галечников (рогатый жаворонок и каменка-плясунья по 19, сизый голубь, бледная завирушка и полевой жаворонок по 4; 154; 129/31; 31 монгольского типа, 19 арктического, 15 европейского, 13 средиземноморского, 10 транспалеарктов);
2.1 – высокогорных (рогатый жаворонок 48, каменка-плясунья 18, жемчужный горный вьюрок 5, горная трясогузка 4, снежный воробей 3; 39; 51/7; 48 арктического типа фауны, 23 монгольского, 15 тибетского, 10 транспалеарктов);
2.2 – среднегорных и предгорных (каменка-плясунья 19, рогатый жаворонок 15, сизый голубь и бледная завирушка по 5, полевой жаворонок 4; 269; 106/40; 32 монгольского типа фауны, 17 европейского, 15 арктического, 14 средиземноморского, 10 транспалеарктов);
3 – местообитаний низкой степени пригодности высокогорных и среднегорных скал и осыпей (кеклик 29, каменная овсянка 22, плешанка 14, индийская пеночка 9, каменка 6; 103; 29/13; 47 монгольского типа фауны, 37 средиземноморского);
4 – высокой степени пригодности альпийских и субальпийских лугов (гималайский вьюрок 27, горный конёк 10, красношапочный вьюрок 6, лесной конёк и клушица по 5; 276; 84/34; 50 тибетского типа фауны, монгольского и европейского по 15);
5 – очень высокой степени пригодности открытых местообитаний, кроме альпийских и субальпийских лугов (черноголовый чекан и скворец по 8, желтоголовая трясогузка и коноплянка по 7, горный конёк 6; 359; 147/51; 34 европейского типа фауны, 22 транспалеарктов, 15 тибетского, 12 средиземноморского);
5.1 – высокогорных лугов и степей (темнобрюхий улар 84, клушица 16; 62; 2/2; 84 тибетского типа фауны, 16 монгольского);
5.2 – среднегорных лугов (горный конёк 19, черноголовый чекан 13, полевой жаворонок 9, коноплянка и красношапочный вьюрок по 6; 306; 67/29; 30 транспалеарктов, 29 тибетского типа фауны, 21 европейского)
5.3 – среднегорных степей (скворец 13, желтоголовая трясогузка 10, черноголовый чекан и серая славка по 8, коноплянка 6; 257; 119/50; 40 европейского типа фауны, 20 транспалеарктов, 14 тибетского типа);
5.4 – предгорных степей (желчная овсянка 10, коноплянка 9, скворец, полевой и домовый воробьи по 6; 570; 79/52; 36 европейского типа фауны, 25 средиземноморского, 21 транспалеарктов);
6 – очень высокой степени пригодности облесенных и мозаичных по облесенности и закустаренности местообитаний (зарничка 9, чечевица 7, зеленая пеночка 6, черногрудый воробей и черный дрозд по 5; 617; 211/79; 33 европейского типа фауны, 19 китайского, 14 средиземноморского, 10 транспалеарктов);
6.1 – хвойных и мелколиственных лесов, стлаников и тугаев (зарничка 17, зеленая пеночка и чечевица по 10, красношапочный вьюрок и седоголовый щегол по 4; 540; 172/61; европейского и китайского типов фауны по 26, 17 сибирского);
6.2 – широколиственных лесов и садов (черный дрозд 15, желтогрудый князёк 9, серая мухоловка и большая горлица по 8, рыжешейная синица 7; 844; 129/52; 43 европейского типа фауны, средиземноморского и китайского по 19);
6.3 – лесных посадок, кустарников, лугов и лугостепей с кустарниками (черногрудый воробей 11, чечевица 7, скворец 6, береговая ласточка 5, серая славка 4; 613; 163/73; 34 европейского типа фауны, 19 средиземноморского, 16 транспалеарктов, 13 китайского типа);
II – антропогенных ландшафтов, кроме садов и лесных посадок (застроенных, рудеральных и рекреационных территорий и агроценозов).
7 – очень высокой степени пригодности (городов, исключая парки и сады, поселков, кошар, животноводческих ферм, свалок и полей; домовый и полевой воробьи 26 и 16, сизый голубь 7, майна 5, рогатый жаворонок 4; 1515; 141/66; 49 транспалеарктов, 18 средиземноморского типа фауны, 15 европейского);
7.1 – участков многоэтажной застройки городов (городская ласточка 36, домовый воробей 31, сизый голубь 17, майна 10, черный стриж 2; 4795; 23/21; 67 транспалеарктов, 18 средиземноморского типа фауны);
7.2 – участков одноэтажной застройки городов, поселков, кошар и животноводческих ферм (домовый и полевой воробьи 28 и 18, сизый голубь 6, майна и рогатый жаворонок по 5; 2052; 122/62; 49 транспалеарктов, средиземноморского и европейского типов фауны по 15);
7.3 – свалок (домовый воробей 31, майна 27, чёрный коршун и деревенская ласточка по 9, черная ворона 7; 238; 28/16; 55 транспалеарктов, 10 европейского типа фауны);
7.4 – полей зерновых (воробьи – полевой 17, черногрудый 11 и домовый 10, желчная овсянка и сизый голубь по 8; 606; 83/43; 43 транспалеарктов, 33 средиземноморского типа фауны, 18 европейского);
7.5 – клеверных полей, залежей и черных паров (серая славка 14, желчная овсянка 13, желтая трясогузка и полевой воробей по 6, теньковка 5; 163; 48/29; 31 европейского типа фауны, 30 транспалеарктов, 27 средиземноморского типа);
III – водно-околоводных местообитаний
8 – низкой степени пригодности (высокогорных и предгорных рек и их пойм; горная и маскированная трясогузки 25 и 19, серая славка 9, черногрудый воробей 6, бурая оляпка 5; 190; 46/25; 32 транспалеарктов, 17 европейского типа фауны, 13 китайского);
9 – малых озер, прудов и болот (чомга 21, лысуха 19, чибис 17, чирок-трескунок 11, ходулочник 5; 208; 71/20; 59 транспалеарктов).
9.1 – низкой степени пригодности (малых высокогорных озер; шилохвость 35, огарь 23, черношейная поганка и чомга по 10, травник 8; 48; 17/8; 35 сибирского типа фауны, 23 монгольского, 20 транспалеарктов);
9.2 – высокой степени пригодности малых среднегорных и предгорных озер, прудов и болот (чомга и лысуха по 21, чибис 18, чирок-трескунок 12, ходулочник 5; 261; 68/22; 61 транспалеарктов).
10 – очень высокой степени пригодности (крупных и средних рек на всех высотах, озера Иссык-Куль и их берегов; малый зуек 19, перевозчик 14, травник 13, горная и маскированная трясогузки 9 и 7; 3314; 50/36; 70 транспалеарктов, 10 монгольского типа фауны);
10.1 – рек (горная трясогузка 35, перевозчик 27, маскированная и белая трясогузки 18 и 5, бурая оляпка 4; 1180; 36/17; 68 транспалеарктов);
10.2 – озера Иссык-Куль (малый зуек 26, травник 18, перевозчик 9, огарь 7, шилохвость 6; 8114; 30/29; 71 транспалеарктов, 12 монгольского типа фауны).
В классификации населения птиц первой половины лета выделено 3 надтипа сообществ, 15 типов и 4 подтипа (Шукуров и др., 2004), а летнего периода в целом – 3 надтипа, 10 типов и 16 подтипов. В сумме число типов и подтипов, соответственно, равно 19 и 26, т.е. на большем материале прослежена и бóльшая дробность разделения сообществ. Состав таксонов в целом сходен, хотя на меньшем по объему материале выявлено большее влияние высот местности (поясности), а на большем количестве данных прослежено влияние облесенности. На среднелетних показателях удалось связать деление на таксоны со степенью пригодности местообитаний. О ней судили по неоднородности гидротермического режима, который определяет плотность населения птиц в сумме по всем видам. С увеличением абсолютных высот местности и соответствующим изменением соотношения тепло- и влагообеспеченности общее обилие птиц и число фоновых видов уменьшается. То же было прослежено в ранее обследованных горных регионах. Максимальные значения суммарного обилия свойственны селитебным местообитаниям из-за их высокой антропогенной кормности. В водно-околоводных местообитаниях то же прослежено для крупных и средних рек и особенно озера Иссык-Куль. В нем велико суммарное количество потребляемых птицами беспозвоночных, рыб и фитомассы.
Доля представителей тибетского, монгольского и реже арктического типа фауны по обилию больше всего на высокогорных территориях, европейского типа и транспалеарктов – в открытых и мозаичных по облесенности местообитаниях, а транспалеарктов – в селитебных участках и водно-околоводных сообществах. Представители этих типов фауны преобладают почти повсеместно. Реже в число преобладающих входят представители китайского типа фауны, которых больше в мозаичных местообитаниях. Доля средиземноморских видов, как правило, тоже велика, но без четкой направленности изменений по высотам, а участие сибирских видов – нигде не достигает 10%.
Пространственно-типологическая структура сообществ
На уровне типа населения птиц основной ряд изменений совпадает с абсолютными высотами местности и уровнем пригодности для птиц местообитаний. Этот ряд отличий определяет гидротермический режим через продуктивность и кормность биоценозов (рис. 2). Пригодность местообитаний последовательно увеличивается от нивальных местообитаний, через высокогорные Аксай-Верхненарынские и аридные среднегорно-предгорные опустыненные и галечниковые участки, скалы и осыпи, к местообитаниям высокой степени пригодности альпийско-субальпийских лугов к открытым и облесенным в той или иной степени местообитаниям (типы 1–6). В трехмерном факторном пространстве граф сходства может быть ориентирован так, что каждый из трех надтипов занимает отдельную плоскость. При этом плотность населения птиц, их общее и фоновое видовое богатство неуклонно увеличиваются с уменьшением высоты местности. Исключение составляют лишь альпийские и субальпийские луга. Здесь общее видовое богатство населения непропорционально меньше из-за малого числа обследованных участков и невысокой изменчивости этих местообитаний.
Рис. 2.
Пространственно-типологическая структура летнего населения птиц Кыргызстана. Сплошные линии – сверхпороговые связи; прерывистые – доминирующие, при отсутствии сверхпороговых; пунктир – дополнительные. Цифры рядом с ними – межклассовое сходство. Цифрами внутри значков отмечены номера таксонов классификации, индекс при них – сходство наборов, вошедших в таксон. Рядом со значками – названия местообитаний, занятых таксоном, лидирующие виды; плотность населения; и через дробь – число встреченных и фоновых видов. Стрелки – направления основных изменений (тренды) и факторы среды, коррелирующие с ними. Местообитания: а – лесные; б – кустарниковые; в – открытые, относительно бедные; г – открытые, богатые; д – поселки; е – водоемы.
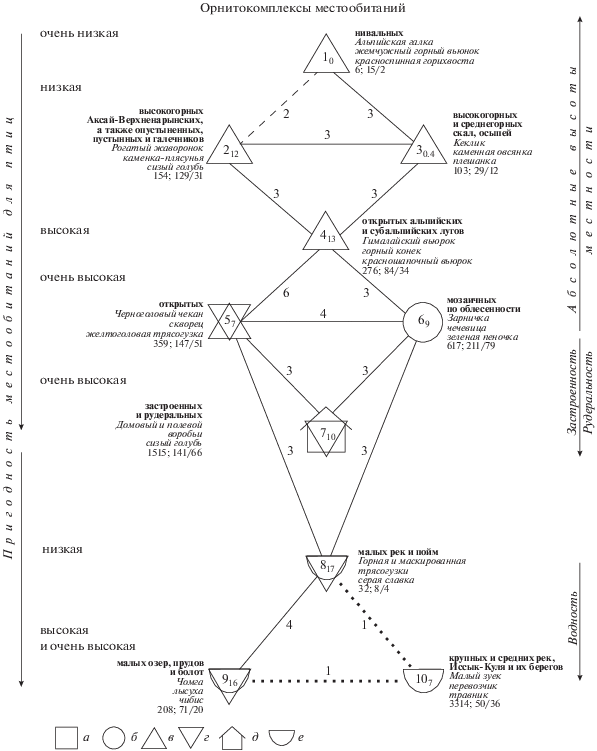
Таким образом, ряд изменений на графе связан со степенью пригодности местообитаний дифференцированно по надтипам населения птиц. В естественных ландшафтах суши неоднородность орнитокомплексов определяют абсолютные высоты местности и степень облесенности – через гидротермический режим. В антропогенных местообитаниях на уровне подтипа на степень пригодности их для птиц влияют: застроенность, этажность, рудеральность, рекреационное использование, распашка и состав возделываемых сельскохозяйственных культур. В водно-околоводных местообитаниях по структурному графу сходства сообществ прослежены различия в степени пригодности местообитаний в зависимости от размера водотоков и водоемов, а также их проточности.
Организация неоднородности орнитокомплексов
Судя по результатам индивидуальной оценки связи между изменчивостью населения и отдельных факторов среды, максимальную значимость имеют тип растительного покрова и степень облесенности местообитаний (по 22% дисперсии). Почти вдвое меньше сила связи с провинциальными различиями ландшафтов и степенью пригодности для птиц местообитаний. В четыре раза меньше связь с водностью и проточностью (табл. 1). При этом существенное приращение в информативности, оцениваемой по доле снятой ими дисперсии, дают лишь первых три фактора из всех приведенных в таблице (по 8%), четвертый фактор учитывает всего 2% дисперсии, а пятый – 1%. Последних два фактора (водность и проточность) полностью скоррелированы с предыдущими. Интегральная информативность всех факторов среды равна 41% дисперсии. Классификационные и структурные режимы учитывают несколько меньшую долю неоднородности матрицы сходства, увеличивая общую информативность всего на 5%.
Таблица 1.
Пространственно-типологическая организация летнего населения птиц Кыргызстана (доля учтенной дисперсии, %)
| Фактор, режим | Индивидуально | Нарастающим итогом |
|---|---|---|
| Тип растительности | 22 | 22 |
| Облесенность | 22 | 30 |
| Провинциальные различия рельефа и климата | 12 | 38 |
| Степень пригодности местообитаний для птиц | 10 | 40 |
| Высоты местности | 7 | 41 |
| Водность | 5 | 41 |
| Проточность | 5 | 41 |
| Режимы классификационные | 33 | – |
| структурные | 25 | – |
| Все режимы | 35 | – |
| Все факторы и режимы | – | 46 |
ОБСУЖДЕНИЕ
Шукуров (1987, 2016) экспертно (умозрительно по сходству) разделил животное население Кыргызстана на три группы в соответствии с рельефом и абсолютными высотами местности: 1 – предгорно-равнинную, 2 – котловинно-долинную и 3 – горную. Последнюю из этих групп он дополнительно подразделил на две подгруппы – среднегорную и высокогорную. Для каждой из этих групп он выделил ряды местообитаний (пустыни и полупустыни; степи; луга альпийские; субальпийские и прочие; пашни; поймы, тугаи; лесные насаждения и сады; болота, пруды, озера; реки, ручьи; кустарники; леса арчовые; еловые и пихтовые; лиственные; скалы, осыпи, галечники; города, поселки, кошары и фермы). К сожалению, учеты птиц нередко проводили по группам этих биотопов, поэтому в полном соответствии с делением Э.Д. Шукурова подразделить орнитокомплексы оказалось невозможным, хотя оба варианта (его и наш формализованный) близки. В частности, нами по сходству были объединены сообщества предгорий, котловин и долин; а также пустынь, полупустынь и степей независимо от уровня высот. Кроме того, отдельно рассмотрено население птиц оз. Иссык-Куль. Информативность такого деления обследованных орнитокомплексов по матрице коэффициентов сходства составила 33% дисперсии. Экспертное разделение летнего населения птиц Кыргызстана, судя по учтенной дисперсии, почти в 1.5 раза менее информативно, чем по результатам кластерного анализа учетных данных.
Анализ изменчивости летнего населения птиц Кыргызстана с использованием примерно вдвое большего по объему материала, собранного не только с мая по июль, но и в августе после усреднения за лето в целом, дал в общем сходные результаты. Это говорит о достаточном количестве данных по обследованной части республики. На более представительном материале прослежена несколько бóльшая дробность совокупности сообществ. Состав групп сходен, при этом на меньшем объеме данных оценка связи с высотами местности была выше, но меньше с облесенностью. Во втором случае удалось выявить связь неоднородности орнитокомплексов с пригодностью местообитаний для птиц. Ее оценивали по гидротермическому режиму и плотности их населения. Максимальные значения силы связи с пространственной неоднородностью населения птиц получены для типа растительности и степени облесённости. Общая информативность представлений об организации летнего населения птиц Кыргызстана соответствует 46% дисперсии матрицы сходства сообществ. Это в 1.5 раза меньше, чем значение, полученное при анализе населения птиц Алтая в первой половине лета, хотя первые места по значимости занимают те же факторы – тип растительности, гидротермический режим и облесенность (Цыбулин, 2009). По Восточно-Европейской и Западно-Сибирской равнинам на первых трех местах по силе и общности связи были близкие факторы – состав лесообразующих пород, облесенность и зональность (подзональность) (Равкин Е.С., Равкин Ю.С., 2005). При этом индивидуальные оценки по ним заметно выше, чем в горных регионах, особенно на Алтае. По интегральной оценке связи со всеми значимыми факторами (59%) орнитокомплексы вышеупомянутых равнин занимают промежуточное положение между Тянь-Шанем и Алтаем.
Эти различия в неоднородности населения птиц обследованных регионов обусловлены спецификой рельефа и увлажнения. Кыргызстан характеризуется максимальной расчлененностью рельефа. Поэтому периодическая смена местообитаний при экстремальной погоде для птиц не представляет труда. Вследствие этого увеличивается сходство разнородных орнитокомплексов в пределах одного и того же ущелья и уменьшается общность между сообществами типологически сходных, но пространственно разобщенных местообитаний. Аридность Тянь-Шаня заметно увеличивает итоговую неоднородность населения птиц. На Алтае расчлененность территории в целом меньше, а площади, занимаемые отдельными ландшафтами, больше. Этим затрудняются местные перекочевки птиц по высотным поясам. При значительной площади поясов требуется больше времени на перемещение птиц. Это способствует увеличению дискретности орнитокомплексов, а плавность смены ландшафтов на Алтае, напротив, увеличивает континуальность населения птиц. На Тянь-Шане неоднородность летнего населения птиц в общем больше, чем на равнинах и особенно по сравнению с орнитокомплексами гор Алтая.
ЗАКЛЮЧЕНИЕ
Итак, увеличение объема материала за счет большего числа обследованных местообитаний и расширения сроков работ на один месяц не увеличило информативности представлений о территориальной неоднородности населения птиц Кыргызстана. Это свидетельствует, во-первых, о достаточности собранных данных для решения поставленных задач. Во-вторых, при дополнительном анализе удалось связать объяснение неоднородности орнитокомплексов с пригодностью для птиц местообитаний. Показана бóльшая информативность результатов формализованной классификации сообществ птиц по сравнению с использованием экспертных (умозрительных) представлений о сходстве орнитокомплексов и связи его с неоднородностью среды. Сходство в организации населения птиц Тянь-Шаня, Алтая и равнин Северной Евразии свидетельствует о значительной степени фрактальности орнитокомплексов (самоподобию целого и его частей). Различия связаны с большей расчлененностью рельефа на Тянь-Шане и с эпизодическими местными перекочевками птиц между соседними местообитаниями и соответственно с бóльшей похожестью населения птиц отдельных ущелий, по сравнению со сходством в растительности. Это приводит к меньшему сходству орнитокомплексов типологически близких местообитаний.
Список литературы
Второв П.П., 1968. Биоэнергетика и биогеография некоторых ландшафтов Терскей-Ала-Тоо. Фрунзе: Илим. 167 с.
Иванов А.И., 1976. Каталог птиц СССР. Л.: Наука. 276 с.
Равкин Ю.С., 1984. Пространственная организация населения птиц лесной зоны (Западная и Средняя Сибирь). Новосибирск: Наука. 364 с.
Равкин Ю.С., Ливанов С.Г., 2008. Факторная зоогеография. Новосибирск: Наука. 205 с.
Равкин Е.С., Равкин Ю.С., 2005. Птицы равнин Северной Евразии. Новосибирск: Наука. 304 с.
Селянинов Г.Т., 1928. О сельскохозяйственной оценке климата // Труды метеорологии. Вып. 20. С. 165–167.
Цыбулин С.М., 2009. Птицы Алтая. Новосибирск: Наука. 236 с.
Штегман Б.К., 1938. Основы орнитогеографического деления Палеарктики. Фауна СССР. Птицы. М.-Л. Т. 1. Вып. 2. 157 с.
Шукуров Э.Д., 1986. Птицы еловых лесов Тянь-Шаня. Фрунзе: Илим. 156 с.
Шукуров Э.Д., 1987. Население наземных позвоночных животных // Атлас Киргизской ССР. Т. I. Природные условия и ресурсы. Главное управление геодезии и картографии при Совете министров СССР. М. С. 118–119.
Шукуров Э.Д., 2016. Зоогеография Кыргызстана. Бишкек. 186 с.
Шукуров Э.Э., Касыбеков Э.Ш., Шукуров Э.Д., Курманкулов А.Б., Лебяжинская И.П., Давранов Э., 2004. Классификация летнего населения птиц Тянь-Шаня // Облiк птахiв: Пiдходы, методики, результаты. Житомир. С. 139–143.
Шукуров Э.Э., Касыбеков Э.Ш., Шукуров Э.Д., Давранов Э., Курманкулов А.Б., 2005. Пространственно-типологическая структура летнего населения птиц Тянь-Шаня // Актуальные вопросы изучения птиц Сибири. Барнаул. С. 230–233.
Янушевич А.И., 1061. Биогеографический обзор птиц Киргизии // Птицы Киргизии. Т. 3. С. 5–58.
Янушевич А.И., Тюрин П.С., Яковлева И.Д., Кыдыралиев А., Семёнова Н.И., 1959, 1960. Птицы Киргизии. Т. 1 и 2. Фрунзе: Изд-во АН Киргизской ССР. 229 и 273 с.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал