

Зоологический журнал, 2022, T. 101, № 3, стр. 336-348
Циркумполярная изменчивость черепа песца (Vulpes lagopus, Carnivora, Canidae) с уточнением таксономического положения островных популяций
О. Г. Нанова *
Научно-исследовательский Зоологический музей Московского государственного университета
имени М.В. Ломоносова
125009 Москва, ул. Большая Никитская, 2, Россия
* E-mail: nanova@mail.ru
Поступила в редакцию 20.12.2020
После доработки 17.02.2021
Принята к публикации 22.02.2021
- EDN: XEPJCJ
- DOI: 10.31857/S0044513422010081
Аннотация
Исследована циркумполярная изменчивость размера и формы краниума песца (V. lagopus L. 1758). Представленные данные включают экземпляры с материковой части Северной Америки, Евразии и большинство нативных островных популяций (о-в Гренландия, о-в Исландия, архипелаг Шпицберген, о-в Св. Лаврентия, о-в Св. Матвея, о-ва Прибылова (о-в Сент-Пол, о-в Сент-Джордж), Командорские о-ва (о-в Беринга, о-в Медный)), а также экземпляр интродуцированной популяции о-ва Атка, Алеутские о-ва. Полученные результаты по структуре краниологической изменчивости сопоставлены с ранее опубликованными данными по структуре генетической изменчивости песца. Показано, что все материковые песцы, а также песцы о-ва Св. Лаврентия, о-ва Св. Матвея, о-ва Исландия, архипелага Шпицберген сходны по размеру и форме черепа. Песцы о-ва Гренландия достоверно мельче материковых песцов, но идентичны им по форме черепа. Наиболее сильно ото всех остальных песцов по форме черепа отличаются песцы Командорских о-вов. С помощью анализа MCLUST мы показали, что командорские песцы могут быть определены с высоким уровнем достоверности (p ≥ 0.95). Морфологическая специфичность командорских песцов согласуется с их генетической обособленностью. Морфологическая специфичность песцов о-ва Гренландия и о-вов Прибылова выражена сильнее, чем их генетическая обособленность. На основе полученных результатов проведена таксономическая ревизия вида. Все материковые песцы, включая евразийскую и североамериканскую материковые части ареала, а также песцы о-ва Св. Лаврентия, о-ва Св. Матвея, архипелага Шпицберген, о-ва Исландия относятся к единому номинативному подвиду V. l. lagopus L. 1758. Песцы о-ва Гренландия принадлежат отдельному мелкому подвиду материкового песца V. l. groenlandicus Bechstein 1799. Песцы о-вов Прибылова V. l. pribilofensis Merriam 1903 также являются подвидами материкового песца. Популяция каждого из двух о-вов Прибылова – о-ва Сент-Пол и о-ва Сент-Джордж – должна быть в дальнейшем выделена в отдельный подвид, учитывая уровень морфологических различий между ними. Песцы Командорских о-вов могут рассматриваться как отдельный вид V. beringensis Merriam 1902 с двумя подвидами V. b. semenovi Ognev 1931 (о-в Медный) и V. b. beringensis Merriam 1902 (о-в Беринга), как ранее предложил Огнев (1931). Песцы о-ва Атка классифицируются как командорские песцы. Это свидетельствует о том, что на о-в Атка песцы были интродуцированы с Командорских о-вов. Учитывая снижение численности командорских песцов, эта информация важна для природоохранной деятельности и сохранения генофонда командорских песцов.
Песец (V. lagopus L. 1758) имеет циркумполярный ареал, населяет также арктические и субарктические острова. Изоляция островных песцов обусловлена отсутствием ледового покрова в акватории в зимний период.
Для песцов выделено большое число подвидов на основании изучения внешних признаков (в основном окраса и размерных характеристик): номинативный V. l. lagopus L. 1758 (Сибирь), V. l. ungava Merriam 1903 (Квебек, Канада), V. l. innuitus Merriam 1902 (Аляска, США), V. l. groenlandicus Bechstein 1799 (о-в Гренландия), V. l. fuliginosus Bechstein 1799 (о-в Исландия), V. l. spitzbergenensis Barrett-Hamilton and Bonhote 1898 (архипелаг Шпицберген), V. l. hallensis Merriam 1900 (о-в Холл, о-в Св. Матвея), V. l. pribilofensis Merriam 1903 (о-ва Прибылова: о-в Сент-Пол, о-в Сент-Джордж), V. l. beringensis Merriam 1903 (о-в Беринга, Командорские о-ва), V. l. semenovi Ognev 1931 (о-в Медный, Командорские о-ва). Командорские песцы были выделены Огневым (1931) в отдельный вид Alopex beringensis Merriam 1902 c двумя подвидами c каждого острова соответственно. Тем не менее степень морфологической обособленности командорских песцов на фоне общей географической изменчивости песцов не изучена. Кроме того, в девятнадцатом веке песцы были интродуцированы на Алеутские о-ва (Bailey, 1993). В настоящий момент интродуцированные популяции сохранились на нескольких островах Алеутской гряды.
Молекулярно-генетические исследования (Dalen et al., 2005; Geffen et al., 2007) показали, что генетическая структура песца на ареале обусловлена характером оледенения акваторий. Было показано, что материковые песцы имеют слабовыраженную генетическую структуру на всем циркумполярном ареале. Краниологические данные говорят в пользу мономорфности песцов на материковой части ареала (Огнев, 1931; Цалкин, 1944; Гептнер, Наумов, 1967; Пузаченко, Загребельный, 2008; Vibe, 1967; Daitch, Guralnick, 2007).
Несмотря на большое количество морфологических исследований песца, детально его циркумполярная изменчивость изучена не была. Как правило, исследовали либо только размерную изменчивость черепа (Vibe, 1967; Frafjord, 1993), либо выборки не включали все островные популяции песцов (Daitch, Guralnick, 2007; Szuma, 2011), либо авторы были сосредоточены на описании особенностей отдельных тихоокеанских популяций (Цалкин, 1944; Загребельный, Пузаченко, 2006; Нанова, 2008, 2009, 2010; Pengilly, 1981; Nanova, Prôa, 2017; Nanova et al., 2017; Martin-Serra et al., 2019).
Задачей данной работы является исследовать изменчивость размера и формы черепа песцов на всем циркумполярном ареале, уделяя особое внимание островным популяциям, и провести таксономическую ревизию подвидовой структуры песца с учетом результатов данного исследования, а также результатов предыдущих морфологических и генетических исследований.
МАТЕРИАЛ И МЕТОДЫ
В работе использован 31 промер черепа и нижней челюсти (рис. 1): кондилобазальная длина черепа (CBL), длина лицевой части (FL), длина мозговой камеры (BL), длина верхнего зубного ряда с клыком (TOOL), длина верхнего ряда малых премоляров (PRML), длина верхнего ряда моляров с хищным зубом (MOLL), длина рострума – расстояние до подглазничного отверстия (ROSTL), скуловая ширина (ZYGW), ширина мозговой коробки (BRCW), расстояние между заглазничными отростками (SORW), ширина рострума на уровне клыков (ROW), мастоидная ширина (MASTW), расстояние между слуховыми барабанами (BULLW), расстояние между мыщелками (CONDW), длина слухового барабана (BULL), высота мозговой коробки в основании саггитального гребня (без слухового барабана) (BRCH), высота черепа в области глазниц (FAH), высота скуловой дуги (ZYGH), высота рострума на уровне второго предкоренного зуба (ROH), заглазничная ширина (PORW), длина нижней челюсти (MANL), длина нижнего зубного ряда с клыком (TOOL2), длина нижнего ряда премоляров (PrmL2), длина нижнего ряда моляров (MolL2), длина проксимального отростка нижней челюсти (RAML), высота проксимального отростка нижней челюсти (MANH1), высота венечного отростка нижней челюсти (MANH2), расстояние от ангулярного отростка нижней челюсти до сочленовного (MANH3), высота нижней челюсти на уровне первого моляра (MANH4).
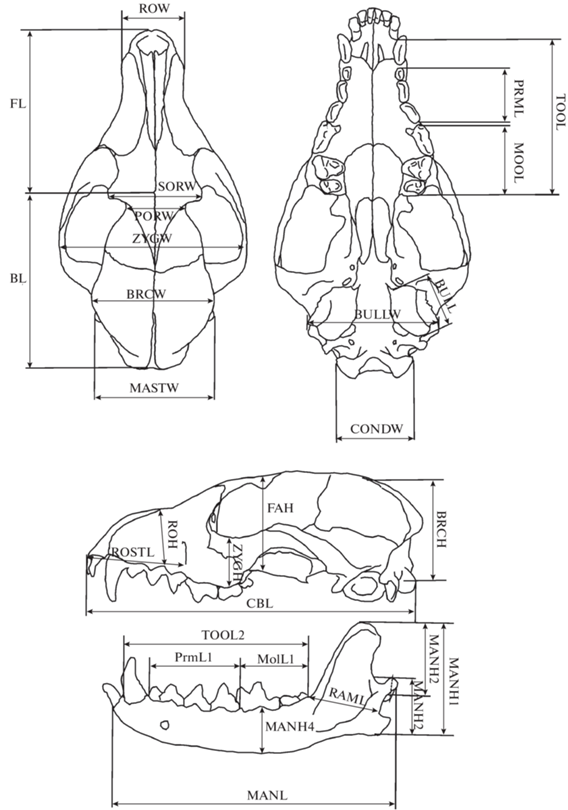
Черепа измеряли электронным штангенциркулем Sylvac. Измеряли только черепа взрослых животных. Возраст определяли, основываясь на степени стертости зубов и облитерации швов (Цалкин, 1944; Загребельный, 2000). Пол животных принимали по этикеточным данным. Ранее с помощью краниометрических исследований было показано, что половые различия черепа у песцов (Нанова, 2009, 2010) сводятся к размерным (самцы немного крупнее самок), при этом вариабельность самцов и самок одинакова. В задачи данной работы не входил анализ половых различий, поэтому при анализе данных мы объединяли самцов и самок.
Всего измерили 326 черепов из следующих локалитетов (рис. 2):
Рис. 2.
Карта локалитетов, включенных в работу. Обозначения локалитетов: 1 – п-ов Чукотка; 2 – о-в Диксон; 3 – Архангельская область; 4 – Северная Европа, материк; 5 – Баффинова Земля; 6 – Провинция Ньюфаундленд и Лабрадор; 7 – мыс Пойнт Барроу; 8 – Андреевская одиночка; 9 – Северо-Западные территории; 10 – о-в Св. Лаврентия; 11 – о-в Гренландия; 12 – архипелаг Шпицберген; 13 – о-в Исландия; 14 – о-в Сент-Пол, о-ва Прибылова; 15 – о-в Сент-Джордж, о-ва Прибылова; 16 – о-в Св. Матвея; 17 – о-в Атка, Алеутские о-ва; 18 – о-в Беринга, Командорские о-ва; 19 – о-в Медный, Командорские о-ва.
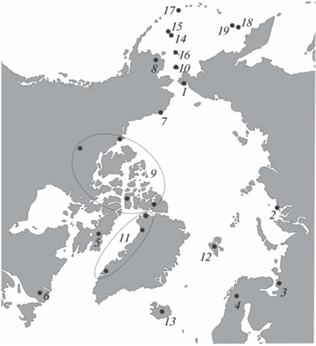
– Евразия, материковая часть: 1 – п-ов Чукотка, 39 экз. (20 самцов, 19 самок); 2 – о-в Диксон, 40 экз. (20 самцов, 20 самок); 3 – Архангельская обл. (20 самцов, 19 самок), 39 экз.; 4 – Северная Европа, материк (Норвегия), 2 экз. (1 самец, 1 самка);
– Северная Америка, материковая часть: 5 – Баффинова Земля, Канада, 19 экз. (6 самцов, 13 самок); 6 – Провинция Ньюфаундленд и Лабрадор, Канада, 10 экз. (4 самца, 1 самка, 5 пол не определен); 7 – мыс Пойнт Барроу, Аляска, США, 17 экз. (9 самцов, 8 самок); 8 – Андреевская одиночка, Аляска, США, 5 экз. (5 пол не определен); 9 – Северо-Западные территории, Канада, 7 экз. (2 самца, 2 самки, 3 пол не определен);
– 10 – о-в Св. Лаврентия, Аляска, США, 12 экз. (7 самцов, 5 самок);
– 11 – о-в Гренландия, Дания, 25 экз. (10 самцов, 4 самки, 11 пол не определен);
– 12 – архипелаг Шпицберген, Норвегия, 2 экз. (пол не определен);
– 13 – о-в Исландия, 2 экз. (пол не определен);
– о-ва Прибылова: 14 – о-в Сент-Пол, 20 экз. (5 самцов, 6 самок, 9 пол не определен); 15 – о-в Сент-Джордж, 5 экз. (2 самца, 1 самка, 2 пол не определен);
– 16 – о-в Св. Матвея, Аляска, США, 1 экз. (самка);
– 17 – о-в Атка, Алеутские о-ва, 1 экз. (пол не определен);
– Командорские о-ва: 18 – о-в Беринга, 40 экз. (20 самцов, 20 самок); 19 – о-в Медный, 40 экз. (20 самцов, 20 самок).
Исследованные черепа хранятся в коллекциях следующих музеев: Зоологический музей МГУ, Москва, РФ; Зоологический Институт РАН, Санкт-Петербург, РФ; Смитсоновский музей Естественной Истории, Вашингтон, США; Американский музей естественной истории, Нью-Йорк, США; Музей естествознания, Берлин, Германия.
В качестве переменной, характеризующей размер животного, выбрана кондилобазальная длина черепа CBL, т.к. длина черепа хорошо коррелирует с размерными характеристиками тела животного (такими как линейные размеры тела, масса тела, Gould, 1966; Emerson, Bramble, 1993; Fitch, 2000). Выборки сравнили между собой по значению CBL с мощью t-теста Стьюдента (тест проводили только для выборок с количеством экземпляров не менее 5).
Далее промеры были логарифмированы для линеаризации аллометрического эффекта (Huxley, 1932; Pimentel, 1979; Reyment, 1991; Klingenberg, 1996) и сопоставимости изменчивости разных выборок (Wayne et al., 1986). Для удаления изометрического размера из данных все промеры делили на центроидный размер соответствующего экземпляра (Нанова, 2008). Статистический анализ проводили на двух типах данных: логарифмированные данные и данные с корректированной изометрической изменчивостью.
Для оценки того, насколько хорошо могут быть идентифицированы особи исследуемых популяций без использования информации о географическом происхождении экземпляров, проведен анализ MCLUST. Этот анализ основан на общей линейной модели, где данные рассматриваются как смесь многомерно-нормальных распределений, каждое из которых имеет свою ковариационную структуру (Лебедев, Лисовский, 2008; Banfield, Raftery, 1993; Fraley, Raftery, 1998, 1999, 2002, 2002а; Nanova, 2014). В отличие от дискриминантного анализа в MCLUST не требуются сведения о групповой принадлежности экземпляра, а по структуре данных оценивается число кластеров и их состав. Таким образом, анализ MCLUST позволяет работать с материалами, когда видовая принадлежность экземпляра неизвестна. Для выбора оптимальной модели, описывающей структуру данных, используется Байесов информационный критерий (BIC). MCLUST оценивает вероятность отнесения каждого экземпляра к выделенным им кластерам. Экземпляр считался достоверно идентифицированным, если вероятность его отнесения к одному из кластеров была равна или выше 0.95 (p ≥ 0.95). Неопределенность классификации каждого экземпляра рассчитывалась как единица минус максимальная вероятность отнесения его к одному из кластеров. Общая неопределенность классификации равна сумме неопределенностей всех экземпляров, деленная на число экземпляров в выборке. MCLUST провели для двух типов данных, используемых в данном исследовании – логарифмированных и корректированных на центроидный размер экземпляра.
Для выявления того, каким образом структурирована изменчивость в исследуемых данных, проводили анализ главных компонент. Для оценки морфологических дистанций рассчитывали квадратичные дистанции Махаланобиса между центроидами географических выборок (рассчитывали только для выборок с количеством экземпляров не менее 5). Для визуализации дистанций между центроидами выборок использовали кластерный анализ, метод UPGMA, на матрице квадратичных дистанций Махаланобиса. Анализ главных компонент, расчет дистанций Махаланобиса и кластерный анализ провели на двух типах данных – логарифмированных и корректированных на центроидный размер экземпляра.
Для оценки степени согласованности краниологической и генетической изменчивости с помощью теста Мантеля сравнили матрицы морфометрических и генетических дистанция между популяциями. В качестве морфометрических дистанций использовали квадратичные дистанции Махаланобиса, рассчитанные на логарифмированных данных. Генетические дистанции взяли из работы Geffen et al., 2007. Использовали средние попарные дистанции (DA) и дистанции φST, рассчитанные на контрольном регионе mtDNA. Для сопоставления выбрали 10 локалитетов сбора черепов и генетических проб, наиболее близких друг другу географически в данном исследовании и исследовании Geffen et al. (2007) соответственно. В анализ вошли следующие выборки: п-ов Чукотка, о-в Диксон, Архангельская обл., Баффинова Земля, мыс Пойнт Барроу, о-в Гренландия, о-в Сент-Пол, о-в Сент-Джордж, о-в Беринга, о-в Медный. Поскольку в статье Geffen et al. (2007) приведены отдельные генетические дистанции для разных частей Гренландии (восточная, южная, западная и северная), мы рассчитали среднее значение этих дистанций для Гренландии в целом.
Все расчеты были проведены в программах Statistica 12, Past 4.03 и программной среде R 4.0.3, пакет MCLUST 5.4.7.
РЕЗУЛЬТАТЫ
Сравнение длин черепа
Средние значения кондилобазальной длины черепа CBL для каждой популяции приведены в табл. 1. Все материковые популяции (п-ов Чукотка, о-в Диксон, Архангельская обл., Баффинова Земля, Провинция Ньюфаундленд и Лабрадор, мыс Пойнт Барроу, Андреевская одиночка, Северо-Западные территории) не различаются между собой по значению CBL на уровне значимости p = 0.01. Песцы о-ва Св. Лаврентия не отличаются по значению CBL от материковых песцов (p > > 0.01). Значения CBL песцов архипелага Шпицберген, о-ва Исландия, о-ва Св. Матвея близки к значениям CBL материковых песцов, однако мы не смогли протестировать достоверность этого сходства ввиду малого объема выборок из этих островных локалитетов. Значения CBL песцов Гренландии достоверно меньше CBL материковых песцов (p < 0.01). Песцы Командорских о-вов и о-вов Прибылова имеют достоверно большие значения CBL (p < 0.01) по сравнению с материковыми песцами. При этом между собой эти четыре тихоокеанские островные популяции (о-в Беринга, о-в Медный, о-в Сент-Пол, о-в Сент-Джордж) не различаются (p > 0.01) по значениям CBL. Значение CBL песца о-ва Атка близко к значениям CBL крупных Тихоокеанских популяций (Командорские о-ва, о-ва Прибылова).
Таблица 1.
Значения кондилобазальной длины черепа CBL песцов для исследуемых локалитетов
| Локалитет | Код локалитета | N | Mean | Min | Max | Std. Dev. |
|---|---|---|---|---|---|---|
| п-ов Чукотка | 1 | 39 | 121.51 | 110.00 | 130.00 | 4.60 |
| о-в Диксон | 2 | 40 | 121.35 | 113.00 | 129.00 | 4.22 |
| Архангельская обл. | 3 | 39 | 122.10 | 114.00 | 130.00 | 4.15 |
| Северная Европа, материк | 4 | 2 | 122.65 | 121.43 | 123.87 | 1.73 |
| Баффинова Земля | 5 | 19 | 120.16 | 113.59 | 125.59 | 4.18 |
| Провинция Ньюфаундленд и Лабрадор | 6 | 10 | 123.26 | 118.93 | 127.69 | 2.46 |
| мыс Пойнт Барроу | 7 | 17 | 122.94 | 117.93 | 127.14 | 3.04 |
| Андреевская одиночка | 8 | 5 | 120.17 | 118.02 | 123.23 | 2.00 |
| Северо-Западные территории | 9 | 7 | 119.33 | 109.39 | 126.64 | 5.23 |
| о-в Св. Лаврентия | 10 | 12 | 122.63 | 116.43 | 130.12 | 4.36 |
| о-в Гренландия | 11 | 25 | 113.06 | 105.31 | 123.07 | 4.28 |
| архипелаг Шпицберген | 12 | 2 | 121.52 | 116.70 | 126.34 | 6.82 |
| о-в Исландия | 13 | 2 | 119.75 | 117.50 | 121.99 | 3.17 |
| о-в Сент-Пол, о-ва Прибылова | 14 | 20 | 127.77 | 117.97 | 135.93 | 4.97 |
| о-в Сент-Джордж, о-ва Прибылова | 15 | 5 | 125.28 | 121.51 | 129.67 | 3.32 |
| о-в Св. Матвея | 16 | 1 | 119.55 | – | – | – |
| о-в Атка, Алеутские о-ва | 17 | 1 | 132.40 | – | – | – |
| о-в Беринга, Командорские о-ва | 18 | 40 | 128.53 | 114.00 | 138.00 | 5.14 |
| о-в Медный, Командорские о-ва | 19 | 40 | 126.53 | 117.58 | 137.00 | 4.91 |
Анализ MCLUST
Логарифмированные данные
MCLUST выявил в структуре данных два кластера. Значение BIC равно 32 980.6, суммарная неопределенность классификации равна 0.625. К первому кластеру относятся все экземпляры с Командорских о-вов (о-в Беринга, о-в Медный, всего 80 экз.) и 1 экз. с о-ва Атка, ко второму кластеру относятся все экземпляры с остального ареала песца (выборки 1–16, всего 245 экз.). В первом кластере 78 экз., т.е. 96.3% экземпляров от общего состава кластера относятся к данному кластеру с вероятностью p ≥ 0.95. Во втором кластере 244 экз., т.е. 99.6% экземпляров от общего состава кластера относятся к данному кластеру с вероятностью p ≥ 0.95.
Корректированные на изометрический размер данные
MCLUST выявил в структуре данных два кластера. Значение BIC равно 93293.7, суммарная неопределенность классификации равна 1.173. К первому кластеру относятся все экземпляры с Командорских о-вов (о-в Беринга, о-в Медный) и экземпляр с о-ва Атка, ко второму кластеру относятся все экземпляры с остального ареала песца (выборки 1–16). В первом кластере 78 экз., т.е. 96.3% экземпляров от общего состава кластера относятся к данному кластеру с вероятностью p ≥ 0.95. Во втором кластере 242 экз., т.е. 98.8% экземпляров от общего состава кластера относятся к данному кластеру с вероятностью p ≥ 0.95.
Анализ главных компонент
Логарифмированные данные
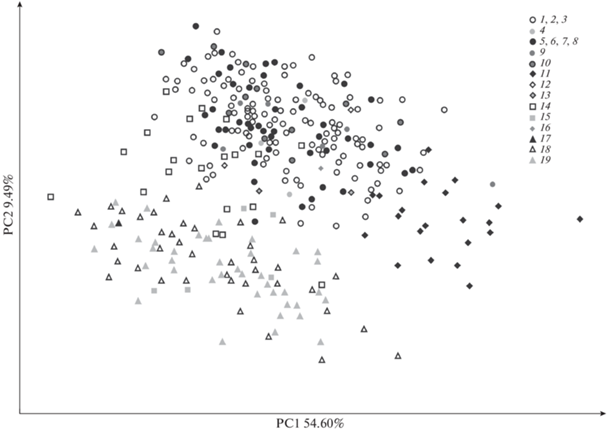
90% общей изменчивости приходится на первые 14 осей главных компонент из 31-й. На первую главную компоненту (PC1) приходится 54.60% общей изменчивости, на вторую главную компоненту (PC2) – 9.49%, на третью главную компоненту (PC3) – 4.67%.
Выборки материковых песцов, собранных в Евразии и Северной Америке, а также выборка песцов о-ва Св. Лаврентия полностью перекрываются между собой в пространстве первых двух главных компонент (рис. 3). Экземпляры, собранные на архипелаге Шпицберген, о-ве Исландия и о-ве Св. Матвея, также попадают в распределение материковых песцов в пространстве первых двух главных компонент. Песцы из Гренландии отличаются от материковой группы песцов по PC1, берущей на себя в основном размерную изменчивость. Все материковые песцы и песцы Командорских о-вов образуют две большие группы, расходящиеся по PC1 и PC2. Песцы о-ва Сент-Пол (о-ва Прибылова) частично перекрываются и с материковыми, и с командорскими песцами в плоскости двух первых главных компонент. Песцы о-ва Сент-Джордж (о-ва Прибылова) перекрываются с командорскими песцами и песцами о-ва Сент-Пол.
Рис. 3.
Распределение экземпляров в пространстве первых двух главных компонент (PC1–PC2) для логарифмированных признаков. Номера локалитетов такие же как на рис. 2.
Корректированные на изометрический размер данные
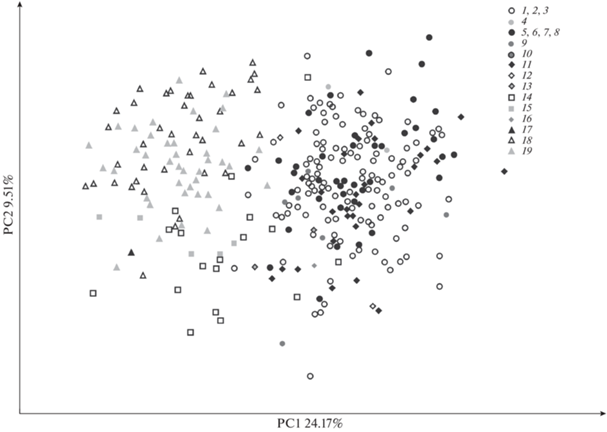
90% общей изменчивости приходится на первые 20 осей главных компонент из 31-й. На первую главную компоненту (PC1) приходится 24.17% общей изменчивости, на вторую главную компоненту (PC2) – 9.51%, на третью главную компоненту (PC3) – 7.24%.
Все материковые выборки песца Евразии и Северной Америки перекрываются между собой в пространстве первых двух главных компонент и представляют собой единую группу (рис. 4). В распределение экземпляров этой материковой группы попадают также песцы архипелага Шпицберген, о-ва Исландия и о-ва Св. Матвея. После удаления размерной изменчивости из данных песцы о-ва Гренландия также полностью перекрываются с материковыми песцами в пространстве PC1– PC2. После удаления размерной изменчивости песцы Командорских о-вов по-прежнему представляют собой отдельную группу, отличающуюся от группы материковых песцов по PC1 и PC2. Песцы о-вов Прибылова после корректировки изометрической изменчивости занимают область между командорскими и материковыми песцами, частично перекрываясь с материковыми песцами.
Рис. 4.
Распределение экземпляров в пространстве первых двух главных компонент (PC1–PC2) для корректированных на изометрический размер признаков. Номера локалитетов такие же как на рис. 2.
Анализ межгрупповых дистанций
Логарифмированные данные
Дерево, построенное на основе квадратичных дистанций Махаланобиса между центроидами отдельных выборок, приведено на рис. 5. Все материковые песцы объединены в один кластер. В этот же кластер входят песцы о-ва Св. Лаврентий и о-ва Гренландия. Все эти выборки равноудалены друг от друга, и дистанции между ними составляют 6.42–25.10. Отдельный кластер, хорошо обособленный от материковых песцов, составляют песцы о-вов Прибылова (о-в Сент-Пол и о-в Сент-Джордж). Дистанции между песцами о-ва Сент-Пол и материковыми песцами составляют 18.6–29.62. Дистанции между песцами о-ва Сент-Джордж и материковыми песцами составляют 43.97–77.67. Стоит отметить, что песцы с двух о-вов Прибылова достаточно сильно удалены друг от друга (квадратичная дистанция Махаланобиса между о-вом Сент-Пол и о-вом Сент-Джордж составляет 27.23), значительно больше, чем две популяции командорских песцов удалены друг от друга и значительно больше, чем гренландские песцы удалены от материковых песцов. Песцы Командорских о-вов (о-в Беринга и о-в Медный) представляют собой отдельный, наиболее удаленный ото всех остальных выборок песцов кластер. Дистанции между выборками материковых песцов и командорскими песцами составляют 36.02–59.44, при этом распределение дистанций одинаково для о-ва Беринга и о-ва Медный. Дистанция между песцами о-ва Беринга и о-ва Медный существенно меньше и составляет 14.99. Дистанции между командорскими песцами и песцами о-ва Сент-Пол составляют 33.64–38.38. Дистанции между командорскими песцами и песцами о-ва Сент-Джордж составляют 59.18–62.77.
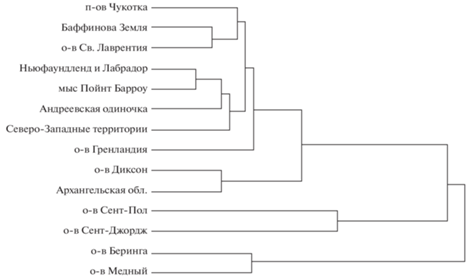
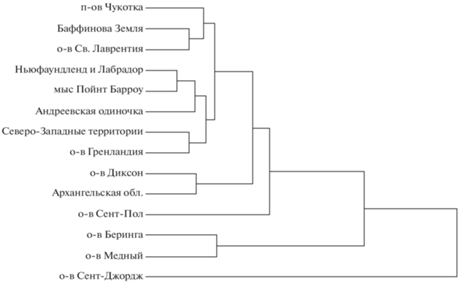
Корректированные на изометрический размер данные
Дерево, построенное на основе квадратичных дистанций Махаланобиса между отдельными выборками на корректированных данных, приведено на рис. 6. Все материковые (включая о-в Св. Лаврентия и о-в Гренландия) песцы объединены в один кластер. Дистанции между этими выборками составляют 6.28–30.22. После удаления изометрической изменчивости из данных о-в Сент-Пол слабее отличается от материковых песцов, тем не менее представляет собой внешнюю ветвь по отношению к материковой популяции. Дистанции между песцами о-ва Сент-Пол и материковыми песцами составляют 17.85–33.72. После удаления изометрической изменчивости из данных о-в Сент-Джордж, напротив, сильнее отличается от материковых песцов. Дистанции между песцами о-ва Сент-Джордж и материковыми песцами составляют 54.55–81.57. О-ва Прибылова не представляют собой единого кластера, дистанция между о-вами Сент-Пол и Сент-Джордж составляет 33.66. Песцы Командорских о-вов представляют собой единый кластер, хорошо обособленный как от материковых песцов, так и от песцов о-вов Прибылова. Дистанция между песцами о-ва Беринга и о-ва Медный – 14.56. Дистанции между выборками материковых и командорских песцов составляют 37.29–52.28. Дистанции между командорскими песцами и песцами о-ва Сент-Пол – 27.92–34.98. Дистанции между командорскими песцами и песцами о-ва Сент-Джордж – 60.50–63.72.
Сравнение матриц морфологических и генетических дистанций
Тест Мантеля показал отсутствие достоверной корреляции между матрицами морфологических и генетических дистанций. Так, коэффициент корреляции между матрицей квадратичных дистанций Махаланобиса и матрицей попарных дистанций (DA) R = 0.33, уровень значимости p = = 0.078. Коэффициент корреляции между матрицей квадратичных дистанций Махаланобиса и матрицей φST дистанций R = 0.31, уровень значимости p = 0.051.
ОБСУЖДЕНИЕ
Результаты анализа краниологической изменчивости песца показали, что все материковые песцы Северной Америки и Евразии, а также песцы о-ва Св. Лаврентия, о-ва Св. Матвея, архипелага Шпицберген, о-ва Исландия имеют сходные размеры и форму черепа. По форме черепа песцы о-ва Гренландия идентичны с материковыми песцами, однако мельче их. Песцы Командорских о-вов и о-вов Прибылова значительно крупнее материковых песцов. При этом между собой животные этих четырех тихоокеанских популяций не различаются по размерам черепа, но хорошо различаются по его форме. Наиболее специфичны и удалены ото всех остальных популяций песцов по форме черепа песцы Командорских о-вов (о-в Беринга и о-в Медный). Песцы Командорских о-вов настолько специфичны по форме черепа, что несмотря на сходные с прибыловскими песцами размеры (а размерная изменчивость часто вносит наибольший вклад в изменчивость между группами), классифицируются как отдельная группа. Этот факт мы продемонстрировали с помощью анализа MCLUST, часто использующегося для установления границ между близкими видами (Cordeiro-Estrela et al., 2008; Nanova, 2014). При этом командорские песцы определяются по черепу относительно материковых песцов с высокой вероятностью, характерной для видового уровня изменчивости. Череп песца с о-ва Атка однозначно классифицируется как принадлежащий командорским песцам.
Настолько полно, как в данном исследовании, изменчивость черепа песца, особенно его формы, ранее не исследовалась. Сравнивая результаты нашей работы с результатами предыдущих работ, выполненных на разных частях ареала, можно прийти к выводу, что они в основном хорошо согласуются между собой. Наиболее подробно изучалась размерная изменчивость песцов. Большие размеры тихоокеанских популяций песцов – песцов Командорских о-вов и о-вов Прибылова – были отмечены многократно (Цалкин, 1944; Огнев, 1931; Гептнер, Наумов, 1967; Vibe, 1967). Мелкие размеры песцов о-ва Гренландия также ранее были отмечены некоторыми авторами (Vibe, 1967; Frafjord, 1993). Наши данные подтвердили эти размерные различия. Предположение о том, что песцы с ближайших к о-ву Гренландия островов (например, о-ва Ellesmer) близки морфологически к гренландским песцам, на наших данных не подтвердилось. Обсуждаемые некоторыми авторами размерные различия между разными материковыми песцами (Frafjord, 1993) скорее всего отражают годичные флуктуации или особенности выборок.
Изменчивость формы черепа песцов исследовалась не так интенсивно. Подробно изучены различия между командорскими и материковыми песцами (Цалкин, 1944; Загребельный, Пузаченко, 2006; Пузаченко, Загребельный, 2008; Нанова, 2008, 2009, 2010; Nanova, Prôa, 2017; Nanova et al., 2017; Martin-Serra et al., 2019). Наиболее сильно особенности формы черепа выражены у песца о-ва Медный, у этих песцов по сравнению с материковыми песцами укорочен рострум, более широко расставлены зубные ряды, характерны очень крупные зубы и широко расставленные скуловые дуги. В работе Pengilly (1981) показано, что прибыловские песцы значительно более крупные и одновременно более мелкозубые по сравнению с материковыми песцами. Мелкозубость песцов с о-ва Сент-Пол позже была подтверждена в работе Daitch, Guralnick, 2007. Pengilly (1981) также показал, что песцы с о-ва Св. Матвея не отличаются от материковых песцов по характеристикам черепа, что мы подтвердили в нашей работе.
В работе Daitch, Guralnick (2007) исследована изменчивость формы первого верхнего моляра с помощью методов геометрической морфометрии. Авторы показали низкий уровень изменчивости формы этого зуба на материке. При этом песцы о-ва Беринга и о-ва Сент-Пол наиболее сильно отличались по форме зуба от материковых песцов и друг от друга (о-в Медный и о-в Сент-Джордж не были включены в их анализ). Исследование полиморфизма дентальных признаков (Szuma, 2011; Gimranov, 2021) показало высокую специфичность песцов Командорских о-вов по сравнению с материковыми песцами. Однако в исследовании Szuma (2011) песцы о-ва Беринга и о-ва Медный при анализе были объединены в одну выборку, что, учитывая большие морфологические различия песцов этих двух островов, делать не следует. В работе Gimranov (2021) автор показал, что песцы о-ва Медный наиболее сильно отличаются от материковых песцов, при этом качественные признаки зубов имеют смесь архаичных и продвинутых черт. По мнению автора найденные различия свидетельствуют о долгой истории изоляции командорских популяций песцов.
Отсутствие географической изменчивости черепа песца на материковой части ареала согласуется с отсутствием выраженной циркумполярной генетической структуры у песцов (Dalen et al., 2005; Geffen et al., 2007). Материковый песец является широко мигрирующим видом. Песцы могут проходить тысячи километров, мигрируя из Евразии в Северную Америку и обратно (Сдобников, 1940; Гептнер, Наумов, 1967; Fuglei, Tarooux, 2019). Основным фактором для миграции является наличие ледового покрова в зимнее время. Острова, расположенные в оледеневающей части океанов, доступны для материковых песцов (Dalen et al., 2005; Geffen et al., 2007). Песцы архипелага Шпицберген, о-ва Св. Лаврентия, о-ва Св. Матвея не являются изолированными популяциями, т.к. эти острова лежат в областях зимнего оледенения поверхности моря и населены материковыми песцами. Песцы о-ва Исландия очень близки генетически и по размеру и по форме черепа к материковым песцам. Песцы о-ва Гренландия близки генетически и морфологически к материковым песцам, при этом генетические дистанции между гренландскими и материковыми песцами несколько больше, чем между разными локалитетами материковых песцов. По форме черепа песцы Гренландии не отличаются от материковых песцов, но достоверно мельче их. Песцы Командорских о-вов наиболее удалены ото всех остальных песцов генетически (разница в генетических дистанциях между материком и Командорскими о-вами в 10 раз больше, чем между разными материковыми локалитетами). Это хорошо согласуется с морфологической спецификой командорских песцов. Песцы о-вов Прибылова значительно ближе генетически к материковым песцам (разница в генетических дистанциях между материком и о-вами Прибылова в 2 раза больше, чем между разными материковыми локалитетами). При этом песцы о-ва Сент-Джордж более удалены генетически от материковых песцов, чем песцы о-ва Сент-Пол. Можно сказать, что морфологическая специфика прибыловских песцов выражена сильнее, чем генетическая. Генетически песцы о-вов Прибылова удалены от песцов Командорских о-вов так же, как и все материковые песцы.
Отсутствие генетической структурированности и структурированности морфологической изменчивости на материке, несоответствие между генетической и морфологической удаленностью песцов о-вов Прибылова и песцов о-ва Гренландия обусловили полученный нами результат по низкому уровню корреляции между генетической и краниологической изменчивостью у песцов на циркумполярном ареале.
Проведенное нами исследование позволило сделать следующие таксономические выводы. Все материковые песцы, включая евразийскую и североамериканскую материковые части ареала, а также включая о-в Св. Лаврентия, о-в Св. Матвея, архипелаг Шпицберген, о-в Исландия, относятся к единому подвиду V. l. lagopus L. 1758. Младшими синонимами для этого названия являются V. l. ungava, V. l. innuitus, V. l. fuliginosus, V. l. spitzbergenensis, V. l. hallensis. Песцы о-ва Гренландия являются отдельным мелким подвидом песца V. l. groenlandicus. Песцы о-вов Прибылова V. l. pribilofensis Merriam 1903 также являются подвидами материкового песца, при этом популяция каждого из двух островов – о-ва Сент-Пол и о-ва Сент-Джордж – в дальнейшем должна быть выделена в отдельный подвид. Структура морфологической изменчивости песцов о-вов Прибылова – разброс индивидуальной изменчивости, половой диморфизм, особенности формы черепа, влияние снижения численности на особенности черепа – нуждается в дальнейшем подробном изучении на большем материале.
Песцы Командорских о-вов имеют глубокие генетические и морфологические отличия от остальных песцов и поэтому могут рассматриваться как отдельный вид V. beringensis Merriam 1902 с двумя подвидами V. b. semenovi Ognev 1931 (о-в Медный) и V. b. beringensis Merriam 1902 (о-в Беринга), как ранее было предложено Огневым (1931). Данный вывод также подтверждается тем, что уровень краниологических различий между командорскими и материковыми песцами соответствует межвидовому уровню изменчивости близких видов лис рода Vulpes и лисоподобных хищников рода Urocyon (Нанова, 2021).
Песцы о-ва Атка наиболее вероятно были интродуцированы с Командорских о-вов. Анализ большего числа экземпляров этой популяции позволит точно установить, с какого именно из двух островов была взята предковая популяция. Ранее с помощью генетических методов для другого острова из Алеутской гряды – Shemya Island (White, Spraker, 2009) – было подтверждено родство с командорскими популяциями песца. Выявление происхождения интродуцированных популяций песцов Алеутских о-вов – важная природоохранная задача, учитывая то, что численность песцов Командорских о-вов (Загребельный, 2000; Goltsman et al., 1996, 2005; Shienok et al., 2017), особенно на о-ве Медный, и песцов о-вов Прибылова (White, 2017) значительно сократилась. Кроме того, детальное сравнительное исследование формы черепа интродуцированных и нативных популяций песцов поможет понять особенности эволюции краниума на начальных этапах заселения островов.
Структура ареала песца и его изменчивость являются хорошей моделью для проверки островного правила (“island rule”; Lomolino, 1985). Островное правило объясняет закономерности изменения размеров животных на островах. Есть работы, связывающие тенденцию к увеличению или уменьшению размеров тела на островах, с изначальным размерным классом вида либо его таксономической принадлежностью (Foster, 1964; Lomolino, 1985; Adler, Levins, 1994). Пример песца наглядно демонстрирует, что в разных экологических условиях вид может реализовать разные стратегии – в одних случаях уменьшаться, в других случаях увеличиваться. Например, песцы тихоокеанских популяций Командорских о-вов и о-вов Прибылова, сформировавшиеся в условиях отсутствия грызунов и более мягкого климата (по сравнению с климатом основного ареала песца), увеличились в размере. Песцы о-ва Гренландия, напротив, уменьшились в размере, что может быть связано с крайней ограниченностью кормовых ресурсов здесь, т.к. основную площадь острова занимает ледник.
Список литературы
Гептнер В.Г., Наумов Н.П., 1967. Млекопитающие СССР. Т. 2. Ч. 1. М.: Высшая школа. 1003 с.
Загребельный С.В., 2000. Командорские подвиды песца (Alopex lagopus beringensis Merriam, 1902 и Alopex lagopus semenovi Ognev, 1931): особенности островных популяций. Дис. … канд. биол. наук. МГУ им. Ломоносова. 166 с.
Загребельный С.В., Пузаченко А.Ю., 2006. Изменчивость черепа песцов беринговского Alopex lagopus beringensis, медновского A. l. semenovi и материкового A. l. lagopus подвидов (Carnivora, Canidae) // Зоологический журнал. Т. 85. № 8. С. 1007–1023.
Лебедев В.С., Лисовский А.А., 2008. Географическая изменчивость метрических признаков черепа и таксономическая структура хомячков Cricetulus группы barabensis (Rodentia, Cricetidae) // Зоологический журнал. Т. 87. № 3. С. 361–374.
Нанова О.Г., 2008. Анализ изменчивости щечных зубов трех видов псовых (Mammalia: Canidae) методами стандартной и геометрической морфометрии // Сб. трудов Зоологического музея МГУ. Т. 49. С. 413–427.
Нанова О.Г., 2009. Структура морфологического разнообразия признаков черепа и зубов трех видов хищных млекопитающих (Mammalia: Carnivora). Дис. … канд. биол. наук. МГУ им. Ломоносова. 222 с.
Нанова О.Г., 2010. Возрастная изменчивость морфометрических признаков черепа материкового песца (Alopex lagopus lagopus) и песца командорских островов (A. l. beringensis, A. l. semenovi; Mammalia // Зоологический журнал. Т. 89. № 7. С. 87–881.
Нанова О.Г., 2021. Сопоставление морфологической дифференциации командорских песцов (Vulpes lagopus semenovi, V. l. beringensis) с межвидовым уровнем различий в родах Urocyon и Vulpes (Canidae) // Зоологический журнал. Т. 100. № 5. С. 573–589.
Огнев С.И., 1931. Звери Восточной Европы и Северной Азии. Т. 2. М.-Л.: Главнаука. 776 с.
Пузаченко А.Ю., Загребельный С.В., 2008. Изменчивость черепа песцов (Alopex lagopus, Carnivora, Canidae) Евразии // Зоологический журнал. Т. 87. № 9. С. 1106–1123.
Сдобников В.М., 1940. Опыт массового мечения песцов // Проблемы Арктики. № 12. С. 106–110.
Цалкин В.И., 1944. Географическая изменчивость в строении черепа песцов Евразии // Зоологический журнал. Т. 23. № 4. С. 156–169.
Adler G.H., Levins R., 1994. The island syndrome in rodent populations // The Quarterly Review of Biology. V. 69. P. 473–490.
Bailey E.P., 1993. Introduction of foxes to Alaskan Islands – history, effect on aviafauna, and eradication. Technical Report. US department of the interior fish and wildlife service. Washington D.C. Opler P.A. (Ed.). 55 p.
Banfield J.D., Raftery A.E., 1993. Model-based Gaussian and non-Gaussian clustering // Biometrics. V. 49. P. 803–821.
Cordeiro-Estrela P., Baylac M., Denys Ch., Polop J., 2008. Combining geometric morphometrics and pattern recognition to identify interspecific patterns of skull variation: case study in sympatric Argentinian species of the genus Calomys (Rodentia: Cricetidae: Sigmodontinae) // The Biological Journal of the Linnean Society. V. 94. № 2. P. 365–378.
Daitch D.J., Guralnick R.P., 2007. Geographic variation in tooth morphology of the arctic fox, Vulpes (Alopex) lagopus // Journal of Mammalogy. V. 88. P. 384–393.
Dalén L., Fuglei E., Hersteinsson P., Kapel C.M.O., Roth J.D., Samelius G., Tannerfeldt M., Angerbjörn A., 2005. Population history and genetic structure of a circumpolar species: the arctic fox // The Biological Journal of the Linnean Society. V. 84. P. 79–89.
Emerson S.B., Bramble D.M., 1993. Scaling, allometry and skull design. The Skull: Functional and Evolutionary Mechanisms. Hanken J., Hall B.K. (eds). Chicago: University of Chicago Press. P. 386–393.
Fitch W.T., 2000. Skull dimensions in relation to body size in nonhuman mammals: The causal bases for acoustic allometry // Zoology. V. 103. P. 40–58.
Foster B.J.,1964. Evolution of mammals on islands // Nature. V. 202. P. 234–235.
Frafjord K., 1993. Circumpolar size variation in the skull of the arctic fox Alopex lagopus // Polar Biology. V. 13. P. 235–238.
Fraley C., Raftery A.E., 1998. How many clusters? Which clustering method? Answers via Model-based cluster analysis // The Computer journal. V. 41. P. 578–588.
Fraley C., Raftery A.E., 1999. MCLUST: software for modelbased cluster analysis // Journal of classification. V. 16. P. 297–306.
Fraley C., Raftery A.E., 2002. Model-based clustering, discriminant analysis, and density estimation // Journal of the American Statistical Association. V. 97. P. 611–631.
Fraley C., Raftery A.E., 2002a. Mclust: software for model-based clustering, density estimation and discriminant analysis. Technical Report 415. Department of Statistics, University of Washington.
Fuglei E., Tarroux A., 2019. Arctic fox dispersal from Svalbard to Canada: one female’s long run across sea ice // Polar Research. V. 38.https://doi.org/10.33265/polar.v38.3512
Geffen E., Waidyaratne S., Dalen L., Angerbjorn A., Vila C., Hersteinsson P., Fuglei E., White P.A., Goltsman M., Kapel C.M., et al., 2007. Sea ice occurrence predicts genetic isolation in the Arctic fox // Molecular Ecology. V. 16. P. 4241–4255.
Gimranov D.O., 2021. The structure of the upper teeth of the red fox (Vulpes vulpes) and arctic fox (V. lagopus) with an analysis of dental variability in insular forms // Russian Journal of Theriology. V. 20. P. 96–110.
Goltsman M.E., Kruchenkova E.P., Macdonald D.W., 1996. The Mednyi Arctic foxes: treating a population imperilled by disease // Oryx. V. 30. № 4. P. 251–258.
Goltsman M.E., Kruchenkova E.P., Sergeev S.N., Volodin I.A., Macdonald D.W., 2005. “Island syndrom” in a population of Arctic fox (Alopex lagopus) from Mednyi Island // Journal of Zoology. V. 267. P. 405–418.
Gould S.J., 1966. Allometry and size in ontogeny and phylogeny // Biological Reviews. V. 41. P. 587–640.
Huxley J.S., 1932. Problems of Relative Growth. Methuen, London. 1st edition. 276 p.
Klingenberg C.P., 1996. Multivariate allometry // Advances in Morphometrics. P. 23–49.
Lomolino M.V., 1985. Body size of mammals on islands. The island rule reexamined // The American Naturalist. V. 125. P. 310–316.
Martín-Serra A., Nanova O., Varón-González C., Ortega G., Figueirido B., 2019. Phenotypic integration and modularity drives skull shape divergence in the Arctic fox (Vulpes lagopus) from the Commander Islands // Biology Letters. V. 15: 20190406.
Nanova O., 2014. Geographical variation in the cranial measurements of the midday jird Meriones meridianus (Rodentia: Muridae) and its taxonomic implications // Journal of Zoological Systematics and Evolutionary Research. V. 52. P. 75–85.
Nanova O., Prôa M., 2017. Cranial features of mainland and Commander Islands Arctic foxes Vulpes lagopus reflect their diverging foraging strategies // Polar Research. Special issue on the Arctic fox biology. V. 36 (S1). P. 68–72.
Nanova O., Prôa M., Fitton L., Evteev A., O’Higgins P., 2017. Comparison of cranial performance between mainland and two island subspecies of the Arctic fox Vulpes lagopus (Carnivora: Canidae) during simulated biting // The Biological Journal of the Linnean Society. V. 121. № 4. P. 923–935.
Pengilly D., 1981. Variation in skull measurements of north American arctic foxes, Alopex lagopus L., and the taxonomic status of A. l. hallensis Merriam and A. I. pribilofensis Merriam. MSc Thesis, University of Alaska, Fairbanks. 206 p.
Pimentel R.A., 1979. Morphometrics: The multivariate analysis of biological data. Kendall/Hunt Publishing Company, Dobuque, Iowa, USA. 276 p.
Reyment R.A., 1991. Multidimensional palaeobiology. Oxford: Pergamon Press. 377 p.
Shienok A., Mamaev E., Mikhnevich J., Ploshnitsa A., 2017. Population dynamics of the Bering arctic fox (Vulpes lagopus beringensis) // 5th International Conference in Arctic Fox Biology. Université du Québec à Rimouski, Québec, Canada. P. 56.
Szuma E., 2011. Ecological and evolutionary determinants of dental polymorphism in the arctic fox Vulpes (Alopex) lagopus // Annales Zoologici Fennici. V. 48. № 4. P. 191–213.
Vibe C., 1967. Arctic animals in relation to climatic fluctuations. Meddelelser om Grønland. Copenhagen: C. A. Reitzel. V. 170. № 5. P. 103–117.
Wayne R.K., Modi W.S., O’Brien S.J., 1986. Morphological variability and asymmetry in the cheetah (Acinonyx jubatus), a genetically uniform species // Evolution. V. 40. P. 78–85.
White P.A., 2017. Population status of the endemic Pribilof Fox Alopex lagopus pribilofensis: Continued decline amidst escalating threats is cause for concern // 5th International Conference in Arctic Fox Biology. Université du Québec à Rimouski, Québec, Canada. P. 24.
White P.A., Spraker T., 2009. Arctic Foxes on Shemya Island, Alaska: An Introduced Species with Unexpected Conservation Value // International conference on arctic fox biology. Vålådalen Mountain Station, Sweden. February 16-18th 2009. Program, abstract and lists of participants. NINA Special Report Angerbjörn A., Eide N.E. & Noren K. (eds), Trondheim. P. 34.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал