

Зоологический журнал, 2022, T. 101, № 4, стр. 439-450
Парадоксы ареалов некоторых видов уток в Северной Евразии
В. П. Белик *
Южный федеральный университет
344006 Ростов-на-Дону, Россия
* E-mail: vpbelik@mail.ru
Поступила в редакцию 15.01.2021
После доработки 28.05.2021
Принята к публикации 30.05.2021
- EDN: QKXGLG
- DOI: 10.31857/S0044513422020039
Аннотация
Рассмотрены особенности ареалов и экологии некоторых видов уток (Aythya fuligula, Anas acuta, Anas penelope), распространенных в степной зоне Казахстана и Западной Сибири. Эти виды исчезли из европейских степей в исторический период, что было вызвано распашкой лугово-степных угодий (обычных мест их гнездования), а также созданием искусственных лесонасаждений, чрезмерным промыслом водоплавающих и околоводных птиц и другими негативными факторами. Судьбу этих видов в степях Восточной Европы могут разделить савка (Oxyura leucocephala) и белоглазый нырок (Aythya nyroca), а также, несомненно, серая утка (Anas strepera) и красноголовый нырок (Aythya ferina), если не будут приняты надлежащие меры по их охране и восстановлению численности.
Гусеобразные птицы (отряд Anseriformes) всегда имели очень важное хозяйственное значение для человека. Ряд видов был успешно доместицирован, некоторые виды служат основными объектами спортивной и промысловой охоты во всех ландшафтных зонах Евразии. Так, объем добычи охотничьих видов гусей и уток в России составляет десятки и сотни тысяч особей, а всего у нас добывают многие миллионы водоплавающих птиц ежегодно (Формозов, 1934, 1940; Янушевич, Золотарева, 1947; Приклонский, 1965; Чельцов-Бебутов, 1967; Кривенко, 1977; Русанов, Бочарников, 1982; Zykov, Ulitin, 2000; Блохин, 2004, 2010; Кривенко, Виноградов, 2008, с. 436–437; и др.). Поэтому корректное управление их популяциями в ключевых регионах может значительно увеличить ресурсы этой ценной охотничьей дичи.
Величина популяций, их численность, общий промысловый запас уток определяются (помимо их гнездовой продуктивности, специфичной для каждого вида), прежде всего обилием птиц, т.е. плотностью населения отдельных таксонов, а также размерами их ареалов. Но при изучении распространения некоторых видов уток, имеющих обширные ареалы в Палеарктике (рис. 1), в частности, шилохвости (Anas acuta), свиязи (Anas penelope), хохлатой чернети (Aythya fuligula), встает ряд вопросов, требующих логического объяснения. Почему эти виды широко распространены в степной зоне Западной Сибири и Казахстана с их резко континентальным, засушливым климатом, но практически отсутствуют на гнездовании в степях Восточной Европы, имеющих более влажный и мягкий климат? Не обитали ли они в европейских степях в прошлом? Какие причины могли привести к сокращению их ареалов в Европе?
Рис. 1.
Ареалы свиязи (а) и хохлатой чернети (б) в середине ХХ в. (по: Исаков, 1952): 1 – граница гнездового ареала, 2 – не вполне выясненная граница, 3 – районы наибольшей гнездовой численности, 4 – места летних встреч и спорадичного гнездования, 5 – районы скопления птиц на линьку, 6 – области зимовок. Зеленая заливка – районы гнездования.

Аналогичные вопросы возникают и по отношению к некоторым другим видам водоплавающих и околоводных птиц, имеющим сходную структуру ареалов, в том числе к лебедю-кликуну (Cygnus cygnus), чернозобой гагаре (Gavia arctica), малой (Larus minutus) и сизой (Larus canus) чайкам и др. В середине ХХ в., по данным Птушенко (1952), в общем так же были распространены лебедь-шипун (Cygnus olor) и серый гусь (Anser anser), почти исчезнувшие в Восточной Европе (рис. 2), но затем благодаря строгой охране частично восстановившие прежние ареалы, расселившись из южных рефугиумов далеко к северу (Кривенко, Виноградов, 2008; Калякин, 2014; del Hoyo, Collar, 2014; European breeding bird atlas …, 2020; и др.). Лишь в 1990-е годы, после открытия весенней охоты на гусей, в России вновь прослежена быстрая деградация ареала и численности всех популяций серого гуся (Казаков и др., 2004; Fox, Leafloor, 2018; Атлас …, 2020; и др.).
Рис. 2.
Изображение ареала серого гуся в Евразии в середине ХХ в. (а) и в начале ХХI в. в Европе (б) (по: Птушенко, 1952; European breeding bird atlas …, 2020): 1 – границы гнездового ареала, 2 – области зимовок, 3 – отдельные места гнездования. Зеленая заливка – области гнездования в середине ХХ века. Точки на растровой карте – современные места гнездования в Европе по квадратам 50×50 км.

Отмеченные черты распространения перечисленных видов гусеобразных существенно контрастируют с ареалами некоторых других видов уток. Так, кряква (Anas platyrhynchos) и чирок-трескунок (Anas querquedula) широко заселяют степную и лесную зоны, не демонстрируя в европейских степях столь резкого сдвига южной границы ареала к северу, как у видов, упомянутых выше.
Особенности границ ареалов всех названных уток могут объясняться, очевидно, как генезисом отдельных видов, так и их аутэкологией, экологическими связями с различными ландшафтами. Существенное влияние на ареалы водоплавающих птиц оказывают также хозяйственная деятельность человека, изменения климата и колебания гидрорежима водоемов (Кривенко, Виноградов, 2008; Михантьев, Селиванова, 2009, 2016; Белик, 2015). Локальная численность уток в значительной степени зависит от успеха их размножения, который, например в Прибайкалье, составляет 24–59%, т.е. в период насиживания кладок от хищничества и других неблагоприятных воздействий погибает около 40–75% яиц, не считая последующей гибели птенцов (Мельников, 2013). Поэтому при выяснении роли различных факторов в динамике популяций уток целесообразно обращать внимание, прежде всего, на гнездовую экологию перечисленных видов.
МАТЕРИАЛЫ И МЕТОДЫ
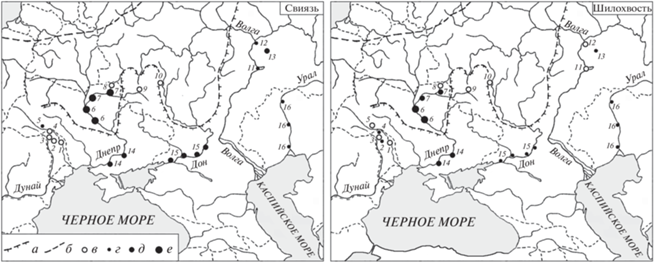
Непосредственные наблюдения над исторической динамикой ареалов уток в степях Северной Евразии, к сожалению, практически отсутствуют. Недостаточно здесь и палеонтологических данных, которые позволили бы установить границы распространения уток в плейстоцен-голоцене. Так, по имеющимся сведениям, в археологических раскопках и аллювиальных отложениях крупных рек юга Русской равнины свиязь и шилохвость найдены, соответственно, на 9 и 8 из 16 изученных местонахождений (рис. 3), а хохлатая чернеть – лишь в одном месте в низовьях р. Десна (Воинственский, 1967). При этом число костных остатков нырковых уток (Aythya sp.) в исследованных районах по каким-то причинам было на порядок меньше, чем речных (Anas sp.) (Белик, 2000, с. 218–221). Однако палеонтологические находки северных видов уток на юге Русской равнины нельзя однозначно интерпретировать как гнездовые, поскольку эти птицы могли появляться здесь также и в миграционные периоды.
Рис. 3.
Палеонтологические находки свиязи и шилохвости в плейстоцен-голоцене на юге Русской равнины (по: Таттар, 1958; Воинственский, 1960, 1967; Ганя, 1972; Белик, 2000; и др.): а – границы Днепровского (Рисского) ледника, б – границы Валдайского (Вюрмского) ледника, в – вид не найден, г – обнаружены костные остатки 1–2 особей, д – обнаружены 3–5 особей, е – собрана 8–31 особь.Места сборов: 1 – Выхватинцы: грот, средний плейстоцен; 2 – Старые Дуруиторы: грот, средний-поздний плейстоцен; 3 – Тринка-1: грот, средний плейстоцен; 4 – Брынзены-1: грот, поздний плейстоцен; 5 – Нижнее Крывче: пещеры, плейстоцен; 6 – Киев–Канев: аллювий Днепра, плейстоцен-ранний голоцен; 7 – Черниговский и Кролевецкий районы: аллювий р. Десна, плейстоцен-ранний голоцен; 8 – Новгород-Северский: стоянка, ранний голоцен; 9 – Авдеево: стоянка, поздний плейстоцен; 10 – Галичья Гора: пещера, древний голоцен; 11 – Липовая Поляна: пещера, древний-начало раннего голоцена; 12 – устье р. Кама: аллювий, плейстоцен-ранний голоцен; 13 – Кармалка: битумы, плейстоцен; 14 – Запорожье–Каховка: аллювий Днепра, плейстоцен-ранний голоцен; 15 – устье р. Чир–Новочеркасск: аллювий Дона, плейстоцен-ранний голоцен; 16 – Уральск–Гурьев (Атырау): аллювий р. Урал, плейстоцен-ранний голоцен.
Поэтому ответы на поставленные выше вопросы можно найти при изучении имеющихся в настоящее время немногочисленных сравнительных данных пока лишь методами каузального анализа. И в основу представленной работы положено именно сравнение материалов по биотопическому размещению гнезд уток и эффективности их размножения на различных водоемах Западно-Казахстанской обл. Казахстана, где в 70–80-е годы ХХ века были собраны данные по гнездованию 7 видов, в том числе кряквы (n = 71 гнездо), шилохвости (n = 21), серой утки (Anas strepera) (n = 62), широконоски (Anas clypeata) (n = 16), чирка-трескунка (n = 13), красноголового нырка (Aythya ferina) (n = 36) и хохлатой чернети (n = 71). Всего на степных озерах тогда было найдено 290 гнезд (Дебело, 1978; Шевченко и др., 1993, 2020). Дополнительные сравнительные данные по распространению, динамике численности и гнездованию уток получены также в результате собственных полевых исследований, проводившихся в течение 1967–2020 гг. в степном Придонье, в Предкавказье, Нижнем Поволжье и Западном Казахстане (Белик, Казаков, 1988; Белик, 2000а, 2003, 2004, 2008; и др.).
Материалы по размножению уток в степной зоне регулярно собирались также в других районах Казахстана (Формозов, 1937; Панченко, 1959; и др.), на юге Западной Сибири (Михантьев, Селиванова, 2009, 2016; и др.), а также в дельте Волги (Луговой, 1963; Русанов, 1979, 2003 и др.), в Предкавказье (Казаков, 1982; Линьков, 1985; Хохлов, 1989; Казаков и др., 2004; и др.) и на юге Украины (Ардамацкая, 1984; Лысенко, 1991; и др.). Однако эти данные получены в степных регионах, существенно различающихся условиями гнездования и видовым составом уток, а особенности биотопического размещения их гнезд в этих публикациях охарактеризованы недостаточно детально. Поэтому наиболее приемлемыми для анализа оказались сведения Шевченко и др. (1993, 2020), хотя они и не были столь значительными по объему, как в других районах (табл. 1).
Таблица 1.
Размещение гнезд уток по биотопам в Западно-Казахстанской области Казахстана (по: Шевченко и др., 1993, 2020)
| Биотопы | Виды уток | ||||||
|---|---|---|---|---|---|---|---|
| Кряква | Серая утка | Шилохвость | Широконоска | Чирок- трескунок |
Красноголовый нырок | Хохлатая чернеть |
|
| Куртины злаков | 18 | 30 | 8 | 5 | 14 | 30 | |
| Разнотравье у берега | 13 | 16 | 2 | 9 | 27 | ||
| Полынь на сусликовинах | 8 | 6 | 9 | 2 | 12 | 8 | |
| Макрофиты на мелководьях | 8 | 4 | 2 | 1 | 10 | 3 | |
| Острова/колонии чаек | 3 | 1 | |||||
| Копны и стога сена | 15 | 3 | 1 | 1 | 3 | ||
| Лесополосы, сады, кусты | 6 | 1 | 1 | ||||
| Гнезда вороны и сороки | 3 | ||||||
| Постройки человека | 3 | ||||||
| Полудупло | 1 | ||||||
| Ниша обрыва | 1 | ||||||
| Прочее | 1 | 1 | |||||
| Всего | 71 | 62 | 21 | 16 | 13 | 36 | 71 |
Биотопы, в которых размещались найденные гнездовья уток, указаны по сведениям авторов (Дебело, 1978; Шевченко и др., 1993, 2020). При неизбежной генерализации этих данных, те гнезда, что были обнаружены в зарослях тростника, рогоза, камыша или осоки в воде, среди так называемых макрофитов, объединены вместе. Вместе показаны и гнезда, располагавшиеся в лесополосах, садах и кустарниках.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ многолетних материалов по экологии уток, собранных в степном Заволжье у границы Европы и Азии (Дебело, 1978; Шевченко и др., 1993, 2020), дает сейчас наиболее показательные сравнительные данные. Там во второй половине ХХ в. у озер было найдено 290 гнезд 7 видов уток. Биотопическое размещение всех этих гнезд показано на рис. 4.
Рис. 4.
Биотопическое распределение гнезд массовых видов уток в степях Западного Казахстана (по: Шевченко и др., 1993, 2020): 1 – куртины пырея и других злаков на берегу; 2 – высокое разнотравье у берега; 3 – полынь и кермек на сусликовинах в степи; 4 – макрофиты (тростник, рогоз, камыш, осока) на мелководьях; 5 – острова с колониями чаек; 6 – копны и стога сена среди весенних разливов; 7 – лесополосы, сады, кусты; 8 – гнезда врановых птиц; 9 – постройки человека; 10 – полудупла; 11 – ниши в обрывах; 12 – прочее.

Как видно из этих данных, кряква в экологическом плане является самым пластичным видом. В степном Заволжье она может использовать до 10 местообитаний, гнездясь как в траве по берегам водоемов, так и среди воды в сравнительно труднодоступных зарослях макрофитов (тростник и др.), а также на островах, на копнах и стогах сена среди разливов, в лесополосах, в том числе в старых гнездах врановых птиц на деревьях, в дуплах, в постройках человека и др. Очевидно, эта пластичность, а также повышенная защищенность гнездовий от хищников и позволяют крякве, по сравнению с другими видами уток, иметь наиболее широкий ареал и наиболее высокую общую численность.
Значительно менее пластична шилохвость, использующая 7 местообитаний, причем более 75% ее гнезд размещалось на земле в траве среди пастбищ по берегам водоемов. Еще более тесно связана со степными биотопами серая утка, у которой 87% гнезд было найдено на земле в траве на расстоянии до 50–100 м от берега водоемов. Аналогичные биотопические связи серой утки с открытыми степными водоемами прослеживаются и в других частях ареала, причем иногда специально подчеркивается редкость находок ее гнезд, устроенных в зарослях тростника среди воды (Исаков, 1952; Долгушин, 1960; Линьков, 1985; Казаков и др., 2004; и др.).
Наконец, 92% гнезд хохлатой чернети было обнаружено на земле в траве не далее 10 м от берега водоемов. При этом в 1970 г. птенцы вылупились лишь в 4 ее гнездах из 33, а в 1971 г. – в 9 из 19 гнезд, находившихся под наблюдением, т.е. доля погибших гнезд достигала 87.9 и 52.6% соответственно. Смертность же птенцов в выводках в 1970, 1971 и 1983 гг. составляла 42.5, 34.6 и 9.6% (n = 7, 16 и 6 наблюдавшихся выводков соответственно) (Шевченко и др., 1993, с. 41).
Сходные результаты дали и наблюдения за гнездованием уток на степных озерах в Наурзумском заповеднике Казахстана, где из 11 найденных гнезд (6 – широконоски, 2 – хохлатой чернети, по 1 гнезду – кряквы, шилохвости и серой утки) хищники разорили 4 гнезда и еще 2 гнезда были брошены, т.е. погибло 55% кладок (Формозов, 1937).
Несомненно, специфичный стереотип гнездования некоторых видов уток на земле в траве по берегам водоемов существенно лимитирует успешность их размножения, их географическое распространение и общую численность. Особенно актуально это в условиях сокращения площади целинных лугово-степных угодий вокруг озер и у рек по мере распашки коренных степей. К тому же это вызвало значительное увеличение концентрации домашнего скота, выпасавшегося у водоемов и приходившего к ним на водопой.
В отличие от рассмотренных видов уток, размножение красноголового нырка теснее связано с водоемами, на которых даже в условиях сухостепного Заволжья свои гнезда в зарослях тростника и рогоза устраивают 28% птиц (рис. 4). В других же регионах этот вид гнездится в основном среди тростников на озерах (Исаков, 1952; Долгушин, 1960; Казаков, 1982; Лысенко, 1991; Казаков и др., 2004; и др.). Поэтому этот типичный представитель лиманного фаунистического комплекса Номадийского типа фауны, связанный ценогенезисом с пустынно-степным поясом Палеарктики (Белик, 2000, 2006), широко распространен в европейских и сибирских степях, а в Европе проникает довольно далеко и в глубь лесной зоны.
Учитывая отмеченные особенности биотопического распределения рассмотренных видов уток, можно попытаться ретроспективно проследить их историю в европейских и азиатских степях, хозяйственное освоение которых человеком в течение XVIII–XX вв. шло совершенно разными темпами и, несомненно, наложило существенный отпечаток на формирование региональных фаун. Так, если степи Восточной Европы начали осваиваться оседлыми славянскими народами еще в XVI–XVII вв., а массовой распашке подверглись в XVIII–XIX вв. (Формозов, 1962; Кириков, 1959, 1966, 1983; Белик, 2000), то широкое заселение степей Северного Казахстана и их интенсивное земледельческое освоение пришлись лишь на середину ХХ в.
Сплошная распашка степей, очевидно, резко ограничила возможности размножения для тех видов уток, которые тяготеют к гнездованию на земле в траве (хохлатая чернеть, серая утка, широконоска, шилохвость и др.). Поэтому вероятно, что “степные” утки и некоторые другие водоплавающие и околоводные птицы, гнездящиеся сейчас в Северном Казахстане, прежде заселяли и европейские степи, и здесь до середины ХХ в. местами еще сохранялись небольшие очаги их гнездования, позволившие части из них начать впоследствии восстановление своих былых ареалов (Птушенко, 1952; Исаков, 1952; Белик, 2000, с. 235). Но точные документальные свидетельства распространения и гнездования отдельных видов в степях на юге России в XVIII–XIX вв., а тем более в XVI–XVII вв., к сожалению, практически отсутствуют.
Частные адаптивные перестройки в экологии некоторых видов иногда могут дать им возможность выйти из-под давления критических лимитирующих факторов и начать заселение совершенно новых местообитаний и географических районов. Одним из наиболее ярких примеров такой успешной адаптации стало недавнее освоение клинтухом (Columba oenas) полых бетонных опор высоковольтных ЛЭП, используемых им сейчас для регулярного гнездования, что позволило этому уязвимому виду-дуплогнезднику начать быстрое восстановление численности и даже заселение новых безлесных территорий (Белик, 2000б; Белик и др., 2010; Белик, Гугуева, 2013; Соколов, Недосекин, 2015; Венгеров, 2016; Шуков, 2018; и мн. др.).
По-видимому, именно за счет подобной адаптации, выработанной когда-то в прошлом, хохлатая чернеть смогла сформировать район повышенной численности в Прибалтике и на северо-западе России (Исаков, 1952; рис. 1). В последнее время аналогичный процесс интенсивно идет в Западной Европе и Прибалтике у серой утки (Кривенко, Виноградов, 2008, с. 380; Иовченко и др., 2010; Коузов, Кравчук, 2012; и др.). Однако степные группировки этого вида сейчас оказались в весьма критическом положении, в связи с чем серая утка внесена в Красные книги Украины (Ардамацька, Горбань, 2009) и Ростовской обл. (Савицкий, 2014). Южнорусские популяции серой утки были рекомендованы для внесения и в Красную книгу России (Белик, 2003б, 2011, 2014).
Длительное же существование популяций серой утки в южных регионах России и Украины в течение ХIХ–ХХ вв., в условиях интенсивного хозяйственного освоения степных территорий, можно объяснить, очевидно, только весьма высокой изначальной численностью этого вида. Так, еще в 1950-е годы серая утка была самой многочисленной среди уток в дельте Дуная; много серых уток обитало в низовьях Днепра, а на островах Черноморского заповедника и на северном побережье Азовского моря до конца ХХ в. отмечались их очень крупные гнездовые группировки, формировавшие местами своеобразные “колонии” (Гавриленко, 1929; Орлов, 1941; Воинственский, 1953; Ардамацкая, 1958, 1984; Лысенко, 1991). Серая утка в ХХ в. по численности лишь немного уступала крякве также на юго-востоке Ростовской обл. и в Калмыкии (Казаков, 1982; Кукиш, 1982; Белик, Казаков, 1988; Белик, 2004; Казаков и др., 2004; Кривенко, Виноградов, 2008). Столь же обычна она была в ХХ в. и в степях Волго-Уральского междуречья в Казахстане (Дебело, 1978; Шевченко и др., 1993, 2020).
Помимо распашки степей, мощное элиминирующее воздействие на “степных” уток и других околоводных птиц до середины ХХ в. оказывали, кроме того, варварский сбор яиц из их гнезд, хищнический лов линных уток, гусей и лебедей, временно теряющих летом способность к полету, а также браконьерская весенняя охота в европейских степях. Детальные описания этих “промыслов” и их последствий для птиц сохранились в ряде известных публикаций (Карелин, 1875; Силантьев, 1898; Качиони, 1910; Житков, 1914; Долгушин, 1929; Кривоносов, 1981; Белик, 2000; Русанов, 2004; и др.).
В степях Западной Сибири и Казахстана тоже существовали подобные промыслы, но они имели меньшее распространение и развивались значительно позже, по мере заселения этих регионов оседлыми земледельцами (Силантьев, 1898; Формозов, 1934). Так, в Астраханской губернии уже в 1869 г. было до 50 человек, специально занимавшихся сбором яиц лебедей, гусей, уток и др., причем один человек брал за день в среднем по тысяче яиц, которые зачастую шли на мыловаренные заводы или в корм свиньям. Тогда же в Астрахани только на рынках ежегодно продавали до полумиллиона яиц диких птиц и примерно столько же сборщики сами использовали в пищу. А на озерах Царицынского уезда в Поволжье два промысловика в 1888 г. лишь за один раз взяли 79 яиц лебедей, 136 яиц гусей и более 700 яиц других птиц. На юге же Западной Сибири, в Тарском округе Омской обл., в конце ХIХ в. за весну собирали всего лишь до 5 тысяч яиц гусей и уток, а в Павлодарской обл. Казахстана на одного промысловика приходилось до 300 гусиных и утиных яиц, но всего за весну там собирали до 50 тысяч яиц, т.е. в 10–20 раз меньше, чем в Астрахани (Силантьев, 1898, с. 383–384).
В начале ХХ в. сбор яиц уток и гусей приобрел массовый характер и в казахстанских степях. Сушкин (1908, с. 790) тогда замечал по этому поводу: “Истребление гнезд практикуется по всей Киргизской степи в самых безобразных размерах. На берегах некоторых озер, например, около Биль-копы, Каналы-куля, я находил настоящие отложения битой яичной скорлупы”. А позже на Чанской системе озер в Новосибирской области крестьянские хозяйства собирали для еды в 1924 г. в среднем по 219 яиц водоплавающих птиц, в 1929 г. по 350 яиц и в 1930 г. по 145 яиц, т.е. в расчете на все хозяйства (47 тысяч) в 1930 г. там собрали около 9 млн. яиц, пригодных в пищу, и еще больше насиженных яиц было погублено (Жданович, 1931; цит. по: Формозов, 1934, с. 258).
Кочевники же в азиатских степях испокон практиковали традиционные загонные зверовые облавы, охоту с борзыми или с ловчими птицами на зверей (Слудский, 1939, 1965; Кириков, 1966; Флинт, Сорокин, 1999), хотя в критические голодные периоды эти скотоводы могли заниматься также промыслом птиц и их яиц (Сушкин, 1908). Тем не менее общее воздействие кочевников на фауну и население птиц в степях Казахстана в прошлом было значительно слабее, чем на юге России, что и позволило сохраниться на севере Казахстана многим лимнофилам до последнего времени.
Но в середине ХХ в. на степных озерах по югу Западной Сибири отмечено повсеместное снижение численности водоплавающих птиц, что было вызвано, как подчеркивают специалисты (Михантьев, Селиванова, 2016), той же массовой распашкой целины, что и на юге России, а также усилением охотничьего пресса, связанным с многократным увеличением числа охотников, развитием дорожной сети и ростом числа транспортных средств.
Еще один важный фактор, оказывающий существенное влияние на распространение “степных” уток, связан с облесённостью берегов водоемов, которая принципиально различается в европейских и казахстанских степях. В Восточной Европе естественные байрачные и аренные леса широко распространены к югу вплоть до побережий Черного и Азовского морей, до Нижнего Дона, а также до Волгограда в Поволжье. Пойменные же леса по Волге и Уралу проникают к югу почти до Каспийского моря (Кулешова, 1965; Зозулин, 1992; Белик, 1997, 2000; и др.). А в более сухих, континентальных степях Казахстана с солонцеватыми и засоленными почвами естественные леса встречаются очень спорадично, в основном на выщелоченных песчаных почвах по речным и морским террасам или на останцовых гранитных возвышенностях (Казахстан …, 1950; Динесман, 1960).
Кроме того, по государственному плану агролесомелиорации зерносеющих регионов СССР, принятому в 1948 г., на юге Украины и России в середине ХХ в. началось интенсивное искусственное лесоразведение, тогда как в скотоводческих районах Казахстана эти работы проводились тогда лишь местами (Доброхвалов, 1950; Успенский, 1966; Митрюшкин, Павловский, 1979; Белик, 2009). В результате в европейских степях по берегам многих рек, озер и прудов сформировалась широкая, густая сеть лесополос, ограничивших возможности гнездования на земле для всех “степных” уток.
На фоне общего сокращения площади, пригодной для размножения этих уток, которое было связано с распашкой степей и их агролесомелиорацией, во второй половине ХХ в. на юге Украины и России в результате появления искусственных лесонасаждений началось активное расселение и увеличение численности дендрофильных птиц и других животных (Белик, 2009). При этом часть из них оказалась весьма опасными хищниками, разорявшими гнезда уток и многих других видов (врановые птицы), а также уничтожавшими молодняк и взрослых птиц (орел-могильник (Aquila heliacal) и особенно ястреб-тетеревятник (Accipiter gentilis)). Воздействие хищничества врановых птиц на виды, гнездящиеся в лугово-степных ландшафтах, неоднократно обсуждалось в печати (Гражданкин, 1984; Кандауров, 1984; Русанов и др., 1984; Флинт, 1984; Бабко, 1989; Белик, 2015; и др.).
О значении уток в питании орла-могильника свидетельствуют наблюдения в Оренбургской обл., где утята в ХIХ в. составляли порой почти исключительный корм орлят (Зарудный, 1888), а в Башкирии этот орел добывал, помимо сусликов и зайцев, также различных птиц, преимущественно уток (Сушкин, 1897). В Татарстане в конце ХХ в. питание 2 из 7 пар орлов-могильников, гнездившихся на побережье водохранилища, на 44–45% состояло из околоводных птиц (Павлов, 1999). В Наурзумском заповеднике на севере Казахстана лимнофильные птицы (утки, чайки, лысухи, поганки, изредка гуси) занимали в питании орла-могильника третье место по встречаемости (15.5%) и второе – по массе (23.4%) (Bragin, 2000). А в Венгрии диких уток добывают около четверти гнездовых пар этого орла, доля же уток в его рационе достигает 2% (Horvath et al., 2010).
В добыче тетеревятника в степном Придонье и Предкавказье утки составляют 0.8–2.4% (Трофименко, 2002; Белик, 2003а; Ильюх, Хохлов, 2010), но в лесистых районах Кавказа их доля среди жертв ястреба увеличивается до 8.4–9.4% (Ткаченко, Витович, 1997; Комаров, 2007). Еще больше уток (36.4% в гнездовой период) тетеревятники добывают местами в лесном поясе (Теплова, 1957). Поэтому облесение степей и расселение по искусственным насаждениям тетеревятника должно, очевидно, значительно усиливать пресс его хищничества на уток.
Важно при этом отметить, что наибольшая угроза хищничества распространяется, прежде всего, на те виды уток, у которых отсутствует или слабо развит половой диморфизм в окраске (савка (Oxyura leucocephala), белоглазый нырок (Aythya nyroca), бэров нырок (Aythya baeri), серая утка), и их самки в равной мере с селезнями элиминируются хищниками, а также охотниками. У диморфных же видов хищники в первую очередь добывают ярко окрашенных самцов, тогда как их самки с покровительственной окраской имеют значительно больше шансов выжить и оставить потомство. В связи с этим общий репродуктивный успех в популяциях мономорфных видов в принципе должен снижаться, по сравнению с аналогичным показателем диморфных видов, и, очевидно, именно поэтому перечисленные выше виды среди прочих промысловых уток оказались первыми кандидатами на включение в Красные книги.
Следует сказать, наконец, о роли климатических изменений в динамике ареалов уток в степной зоне. Циклические колебания увлажненности и теплообеспеченности Северной Евразии и связанные с этим изменения гидрорежима многих степных водоемов всегда вызывали значительные пульсации ареалов и численности обитающих на них лимнофильных птиц, зачастую разнонаправленные у видов разного генезиса (Кривенко, 1991; Кривенко, Виноградов, 2008). В недавнем прошлом резко континентальный степной климат обеспечивал весной наполнение талыми водами многих замкнутых озер и лиманов, которые служили местом гнездования уток и других водоплавающих и околоводных птиц. Но с середины ХХ в. накопление снега в степях из-за потепления зимнего климата значительно уменьшилось, а при снеготаянии влага стала быстро впитываться в слабо промерзающую почву. Поэтому весеннее заполнение озер и различных водоемов талой водой, в частности в степном Заволжье, в последние десятилетия практически прекратилось (Линдеман и др., 2005).
Высыхание естественных водоемов в степи во второй половине ХХ в. частично компенсировалось широким строительством оросительных систем, прудов и водохранилищ. Но в 1990-е годы орошаемое земледелие в засушливых районах пришло в упадок, подача пресной воды сократилась, и часть каналов и прудов пересохли, что лишило уток также и многих искусственных водоемов (Белик, 2015).
Наряду с наблюдающимся сейчас общим потеплением и уменьшением континентальности степного климата, для нынешнего переходного климатического периода весьма характерно также усиление неустойчивости многих метеорологических процессов (Будыко, 1979). Поэтому в отдельные годы в степях развиваются очень сильные летние засухи, приводящие к периодическому пересыханию почти всех степных прудов и других водоемов и к исчезновению связанных с ними птиц.
В результате этих климатических перестроек в европейских степях в последние десятилетия катастрофически снизилась численность и у красноголового нырка (Атлас …, 2020), прежде благополучно пережившего критические периоды распашки целины, усиления пастбищной нагрузки и лесомелиорации степей. Можно полагать, что это есть ранняя стадия формирования еще одного примера “парадокса ареала”, наметившегося сейчас вследствие изменений степного климата.
ЗАКЛЮЧЕНИЕ
Проведенный анализ позволяет обоснованно полагать, что ряд видов уток и некоторых других водоплавающих и околоводных птиц, гнездящихся сейчас на озерах в степях Западной Сибири и Казахстана, в прошлом были распространены также и в европейских степях. Но в XVIII–XX веках они оказались вытеснены отсюда, по-видимому, в результате заселения степной зоны оседлыми земледельческими народами, последующей распашки целины, широкого искусственного лесоразведения, а также чрезмерного охотничьего промысла и воздействия некоторых других негативных факторов (Белик, 1997а, 2000, 2015).
О сравнительно недавнем исчезновении гнездовий этих видов свидетельствуют отдельные случаи их летних встреч и находок гнезд на юге России и Украины вплоть до середины ХХ в. (Исаков, 1952; Птушенко, 1952; Белик, 2000; и др.), а также их спорадичное гнездование до последнего времени в Калмыкии и степном Заволжье (Дебело, 1978; Кукиш, 1982; Шевченко и др., 1993, 2020; Кривенко, Виноградов, 2008; и др.). Единичные находки реликтовых гнездовий хохлатой чернети недавно отмечены на степных островах оз. Маныч-Гудило (Цапко, 2005, 2007; Цапко, Бобенко, 2012), а случайное гнездование свиязи было подтверждено в степях Саратовского Заволжья (Атлас …, 2020).
Таким образом, восстановленные гнездовые ареалы хохлатой чернети, шилохвости и, возможно, свиязи должны включать всю степную зону Восточной Европы, где птицы размножались еще в недавнем прошлом. Сейчас же драматичную судьбу этих исчезнувших в европейских степях видов могут разделить савка и белоглазый нырок, включенные в Красную книгу России (2001) и Червону книгу України (2009), а также, несомненно, серая утка и красноголовый нырок, если не будут приняты надлежащие меры по их охране и восстановлению численности.
Список литературы
Ардамацкая Т.Б., 1958. Некоторые особенности гнездования утиных в районе Черноморского заповедника // Труды Черноморского заповедника. Вып. 2. С. 35–64.
Ардамацкая Т.Б., 1984. Гнездование утиных и ржанкообразных на островах Тендровского залива Черноморского заповедника // Орнитология. Вып. 19. С. 41–49.
Ардамацька Т.Б., Горбань І.М., 2009. Нерозень // Червона книга України: Тваринний світ. Київ. С. 410.
Атлас гнездящихся птиц европейской части России, 2020. М.: Фитон ХХI. 908 с.
Бабко В.М., 1989. Хищническая деятельность серой вороны в пойме р. Десны // Врановые птицы в естеств. и антропоген. ландшафтах: Материалы 2 Всесоюз. совещ. Ч. 2. Липецк. С. 42–43.
Белик В.П., 1997. Проникновение северных дендрофильных видов птиц в глубь пустынь Казахстана // Беркут. Т. 6. Вып. 1–2. С. 19–22.
Белик В.П., 1997а. Некоторые последствия использования пестицидов для степных птиц Восточной Европы // Беркут. Т. 6. Вып. 1–2. С. 70–82.
Белик В.П., 2000. Птицы степного Придонья: Формирование фауны, ее антропогенная трансформация и вопросы охраны. Ростов-на-Дону: Изд-во РГПУ. 376 с.
Белик В.П., 2000а. К летней орнитофауне песков Аккумы и их окрестностей (Уральская область, Западный Казахстан) // Материалы к распространению птиц на Урале, в Приуралье и Зап. Сибири. Екатеринбург. С. 9–17.
Белик В.П., 2000б. О катастрофическом снижении численности восточноевропейской популяции клинтуха // Редкие, исчезающие и малоизученные птицы России: Сб. науч. статей. М.: СОПР. С. 85–90.
Белик В.П., 2003. К летней орнитофауне бассейна оз. Шалкар, Республика Казахстан (неворобьиные) // Стрепет. Вып. 2. С. 22–35.
Белик В.П., 2003а. Хищничество тетеревятника и его роль в биоценозах // Ястреб-тетеревятник: Место в экосистемах России: Материалы к 4 конф. по хищным птицам Северной Евразии. Пенза – Ростов. С. 146–168.
Белик В.П., 2003б. Имя из “Красной книги”: Наземные позвоночные животные степного Придонья, нуждающиеся в особой охране. Ростов-на-Дону: Донской издательский дом. 432 с.
Белик В.П., 2004. Птицы долины озера Маныч-Гудило: Non-Passeriformes // Труды гос. природного заповедника “Ростовский”. Вып. 3: Биоразнообразие заповедника “Ростовский” и его охрана. Ростов-на-Дону: Донской издательский дом. С. 111–177.
Белик В.П., 2006. Фауногенетическая структура авифауны Палеарктики // Зоологический журнал. Т. 85. № 3. С. 298–316.
Белик В.П., 2008. Материалы к мониторингу орнитофауны Камыш-Самарских озер (Западный Казахстан) // Стрепет. Т. 6. Вып. 2. С. 5–28.
Белик В.П., 2009. Птицы искусственных лесов степного Предкавказья: Состав и формирование орнитофауны в засушливых условиях. Кривой Рог. 216 с.
Белик В.П., 2011. Обоснование включения серой утки в Красную книгу Ростовской области // Проблемы Красных книг и преподавание охраны природы: Науч.-метод. сборник. Ростов-на-Дону. С. 109–112.
Белик В.П., 2014. Состояние популяций серой утки на юге России // Казарка. № 17. М. С. 118–124.
Белик В.П., 2015. Степные птицы в ХХI веке: проблемы и перспективы // Степные птицы Северного Кавказа и сопредельных регионов: Изучение, использование, охрана: Материалы Междунар. конф. Ростов-на-Дону. С. 11–37.
Белик В.П., Ветров В.В., Милобог Ю.В., 2010. Ренессанс клинтуха в Восточной Европе: демографический потенциал новой адаптации // Стрепет. Т. 8. Вып. 1. С. 70–74.
Белик В.П., Гугуева Е.В., 2013. Новая экологическая адаптация клинтуха и ее популяционное значение // Поволжский экологический журнал. № 2. С. 123–131.
Белик В.П., Казаков Б.А., 1988. Современное состояние фауны гусеобразных Нижнего Дона (в пределах Ростовской области) // Ресурсы животного мира Северного Кавказа: Тезисы докладов науч.-практ. конф. Ставрополь. С. 12–17.
Блохин Ю.Ю., 2004. Охотничья добыча уток и гусей по данным лицензий в Ямало-Ненецком автономном округе // Казарка. № 10. С. 119–141.
Блохин Ю.Ю., 2010. Добыча водоплавающей дичи в Московской области по данным именных разовых лицензий // Казарка. № 13. С. 104–122.
Будыко М.И., 1979. Проблема углекислого газа. Л.: Гидрометиздат. 59 с.
Венгеров П.Д., 2016. Гнездование клинтуха Columba oenas в опорах линий электропередачи в окрестностях Воронежского заповедника // Русский орнитологический журнал. Т. 25. № 1265. С. 1031–1036.
Воинственский М.А., 1953. Птицы плавней дельты Дуная // Наук. зап. Київського ун-ту. Т. 12. Вип. 3. С. 49–74.
Воинственский М.А., 1960. Птицы степной полосы Европейской части СССР. Киев: Изд-во АН УССР. 292 с.
Воинственский М.А., 1967. Ископаемая орнитофауна Украины // Природная обстановка и фауны прошлого. Вып. 3. Киев: Наукова думка. С. 3–76.
Гавриленко Н.И., 1929. Птицы Полтавщины. Полтава. 133 с.
Ганя И.М., 1972. История орнитофауны Молдавии с позднего миоцена до наших дней // Фауна наземн. позвоночных Молдавии и проблемы ее реконструкции. Кишинев: Штиинца. С. 20–43.
Гражданкин А.В., 1984. Влияние врановых птиц на успех гнездования птиц открытых ландшафтов // Экология, биоценотическое и хозяйственное значение врановых птиц. М.: Наука. С. 163–164.
Дебело П.В., 1978. Материалы по распространению и экологии гнездящихся уток Уральской области // Биология птиц в Казахстане. Алма-Ата: Наука КазССР. С. 84–93.
Динесман Л.Г., 1960. Изменение природы северо-запада Прикаспийской низменности. М.: Изд-во Академии наук СССР. 160 с.
Доброхвалов В.П.,1950. Очерк истории степного лесоразведения. М.: Изд-во МГУ. 208 с.
Долгушин И., 1929. Хищнический промысел водоплавающих птиц в Барабинской степи // Uragus. Кн. 10. № 2. С. 17–18.
Долгушин И.А., 1960. Птицы Казахстана. Т. 1. Алма-Ата: Изд-во Академии наук КазССР. 470 с.
Житков Б.М., 1914. О промысле и охране птиц в дельте Волги // Материалы к познанию русского охотничьего дела. Вып. 4. СПб. С. 1–59.
Зарудный Н.А., 1888. Орнитологическая фауна Оренбургского края // Зап. Импер. Академии Наук. Приложение к т. 57. № 1. СПб. С. 1–338.
Зозулин Г.М., 1992. Леса Нижнего Дона. Ростов-на-Дону: Изд-во Ростов. ун-та. 203 с.
Ильюх М.П., Хохлов А.Н., 2010. Хищные птицы и совы трансформированных экосистем Предкавказья. Ставрополь. 760 с.
Иовченко Н.П., Рычкова А.Л., Смирнова О.П., 2010. Стремительное освоение водоемов Санкт-Петербурга серой уткой (Anas strepera) в начале 21 века // Орнитология в Северной Евразии: Материалы XIII междунар. орнитол. конф. Северной Евразии: Тезисы докладов. Оренбург. С. 144.
Исаков Ю.А., 1952. Подсемейство утки Anatinae // Птицы Сов. Союза. Т. 4. М.: Советская наука. С. 344–635.
Казаков Б.А., 1982. Отряд Гусеобразные // Ресурсы живой фауны. Ч. 2: Позвоночные животные суши. Ростов-на-Дону: Изд-во Ростов. ун-та. С. 178–196.
Казаков Б.А., Ломадзе Н.Х., Белик В.П., Хохлов А.Н., Тильба П.А. и др., 2004. Птицы Северного Кавказа, том 1: Гагарообразные, Поганкообразные, Трубконосые, Веслоногие, Аистообразные, Фламингообразные, Гусеобразные. Ростов-на-Дону: Изд-во РГПУ. 398 с.
Казахстан: Общая физико-географическая характеристика, 1950. М.: Изд-во Академии наук СССР. 491 с.
Калякин М.В. (ред.), 2014. Полный определитель птиц европейской части России. Ч. 1. М.: Фитон-ХХI. 268 с.
Кандауров Е.К., 1984. Проблема врановых в антропогенных ландшафтах // Экология, биоценотическое и хоз. значение врановых птиц. М.: Наука. С. 22–24.
Карелин Г.С., 1875. Разбор статьи г. А. Рябинина “Естественные произведения земель Уральского казачьего войска”, извлеченной из книги его: Материалы для географии и статистики России. Уральское казачье войско. 2 части. СПб. 1866 г. Санкт-Петербург. 113 с.
Качиони С., 1910. Несколько слов об охотничьем промысле в Астраханской губернии // Природа и охота. № 11. С. 7–11.
Кириков С.В., 1959. Изменения животного мира в природных зонах СССР (ХIII–ХIХ вв.): Степная зона и лесостепь. М.: Изд-во Академии наук СССР. 175 с.
Кириков С.В., 1966. Промысловые животные, природная среда и человек. М.: Наука. 348 с.
Кириков С.В., 1983. Человек и природа степной зоны: Конец Х – середина ХIХ в.: Европейская часть СССР. М.: Наука. 125 с.
Комаров Ю.Е., 2007. К биологии птиц Республики Северная Осетия–Алания // Проблемы развития биологии и экологии на Сев. Кавказе. Ставрополь. С. 87–99.
Коузов С.А., Кравчук А.В., 2012. Серая утка в восточной части Финского залива: история заселения, биология и миграции // Казарка. Т. 15. № 2. С. 106–136.
Красная книга Российской Федерации (Животные), 2001. М. 862 с.
Кривенко В.Г., 1977. Добыча водоплавающих птиц в Астраханской области // Ресурсы пернатой дичи побережий Каспия и прилежащих районов (охрана, использование и изучение). Астрахань. С. 35–38.
Кривенко В.Г., 1991. Водоплавающие птицы и их охрана. М.: Агропромиздат. 271 с.
Кривенко В.Г., Виноградов В.Г., 2008. Птицы водной среды и ритмы климата Северной Евразии. М.: Наука. 588 с.
Кривоносов Г.А., 1981. Природная среда водоемов и водоплавающие птицы дельты Волги (XVIII–XX вв.) // Бюллетень Московского общества испытателей природы. Отд. биол. Т. 86. Вып. 5. С. 30–41.
Кукиш А.И., 1982. Животный мир Калмыкии. Птицы. Элиста. 128 с.
Кулешова Л.В., 1965. Типы птичьего населения долины Нижней Волги и прилежащих участков Северного Каспия // Орнитология. Вып. 7. С. 235–243.
Линдеман Г.В., Абатуров Б.Д., Быков А.В., Лопушков В.А., 2005. Динамика населения позвоночных животных Заволжской полупустыни. М.: Наука. 252 с.
Линьков А.Б., 1985. К экологии гнездования уток озера Маныч-Гудило // Птицы Сев.-Зап. Кавказа: Сборник научных трудов. М.: ЦНИЛ Главохоты РСФСР. С. 78–95.
Луговой А.Е., 1963. Птицы дельты реки Волги // Фауна и экология птиц дельты реки Волги и побережий Каспия // Труды Астраханского заповедника. Вып. 8. Астрахань. С. 9–185.
Лысенко В.И., 1991. Фауна Украины. Т. 5: Птицы. Вып. 3: Гусеобразные. Киев: Наук. думка. 205 с.
Мельников Ю.И., 2013. Популяционный гомеостаз – основной путь поддержания численности водоплавающих птиц Прибайкалья // Известия Самарского научного центра РАН. Т. 15. № 3 (3). С. 1133–1137.
Митрюшкин К.П., Павловский Е.С., 1979. Лес и поле. М.: Колос. 280 с.
Михантьев А.И., Селиванова М.А., 2009. Экологические основы прогнозирования продуктивности и численности уток // Казарка. Т. 12. № 1. С. 47–67.
Михантьев А.И., Селиванова М.А., 2016. Динамика численности некоторых видов уток на юге Западной Сибири (Северная Кулунда) в связи с гидроклиматическими и метеорологическими условиями // Казарка. Т. 19. № 2. С. 67–80.
Орлов П.П., 1941. Орнiтофауна Черкаського району // Наук. зап. Черкаського пед. iн-ту. Вип. 2. С. 1–75.
Павлов Ю.И., 1999. Новое в питании орла-могильника в Татарстане // Королевский орел: Распространение, состояние популяций и перспективы охраны орла-могильника (Aquila heliaca) в России: Сборник науч. трудов. Серия: Редкие виды птиц. Вып. 1. М.: Союз охраны птиц России. С. 95.
Панченко С.Г., 1959. Материалы по распространению и экологии пластинчатоклювых Центрального Казахстана // Труды Института зоологии Академии наук КазССР. Т. 10. С. 35–55.
Приклонский С.Г., 1965. Опыт оценки количества водоплавающих птиц, добываемых в Советском Союзе // География ресурсов водоплавающих птиц в СССР, состояние запасов, пути их воспроизводства и правильного использования: Тез. докладов совещания. Ч. 1. М. С. 21–24.
Птушенко Е.С., 1952. Подсемейство гусиные Anserinae // Птицы Советского Союза. Т. 4. М.: Советская наука. С. 255–344.
Русанов Г., Бочарников В., 1982. Водоплавающая дичь дельты Волги // Охота и охотничье хозяйство. № 9. С. 12–14.
Русанов Г.М., 1979. Современные условия гнездования кряквы в западной части дельты Волги и меры по их улучшению // Природная среда и птицы побережий Каспийского моря и прилежащих низменностей. Баку. С. 131–145.
Русанов Г.М., 2003. Размножение водоплавающих птиц в дельте Волги // Казарка. № 9. М. С. 323–343.
Русанов Г.М., 2004. Несколько слов об охоте в дельте Волги: свидетельства минувшего и комментарии современника // Стрепет. Т. 2. Вып. 1. С. 107–115.
Русанов Г.М., Кривоносов Г.А., Бондарев Д.В., Реуцкий Н.Д., Гаврилов Н.Н., 1984. Численность серой вороны в низовьях дельты Волги и ее роль в жизни птиц // Экология, биоценотическое и хоз. значение врановых птиц. М.: Наука. С. 143–147.
Савицкий Р.М., 2014. Серая утка // Красная книга Ростовской области. Т. 1: Животные. 2-е изд. Ростов-на-Дону. С. 180.
Силантьев А.А., 1898. Обзор промысловых охот в России. СПб. 619 с.
Слудский А.А., 1939. Азиатская борзая тазы и охота с ней. Алма-Ата. 28 с.
Слудский А.А., 1965. Казахская борзая тазы и охота с ней // По охотничьим просторам Казахстана. Алма-Ата: Кайнар. С. 105–121.
Соколов А.Ю., Недосекин В.Ю., 2015. Изменение состояния популяции клинтуха в Центральном Черноземье в связи с новой гнездовой адаптацией // 14-я Международная орнитол. конф. Северной Евразии. 1: Тезисы. Алматы. С. 455–456.
Сушкин П.П., 1987. Птицы Уфимской губернии // Материалы к познанию фауны и флоры Рос. империи. Отд. зоол. Вып. 4. С. I–XI, 1–331.
Сушкин П.П., 1908. Птицы Средней Киргизской степи (Тургайская область и восточная часть Уральской) // Материалы к познанию фауны и флоры Российской империи. Отд. зоол. Вып. 8. С. 1–803.
Таттар А.В., 1958. Фауна млекопитающих и птиц из верхнечетвертичных отложений пещер Верхнего Дона и Жигулей и условия ее существования // Учен. зап. Ленинград. пед. ин-та. Т. 179. С. 113–189.
Теплова Е.Н., 1957. Птицы района Печоро-Илычского заповедника // Труды Печоро-Илычского заповедника. Вып. 6. С. 5–115.
Ткаченко И.В., Витович О.А., 1997. Питание тетеревятника в Тебердинском заповеднике // Кавказ. орнитол. вестник. Вып. 9. Ставрополь. С. 122–130.
Трофименко В.В., 2002. Зимняя экология ястребов на юге Ростовской области // Птицы Южной России: Материалы Международной орнитол. конф. “Итоги и перспективы развития орнитологии на Сев. Кавказе в XX) веке”, посвященной 20-летию деятельности Северокавказской орнитол. группы. Ростов-на-Дону. С. 165–167.
Успенский С.Н., 1966. Полезащитное лесоразведение в Казахской ССР и перспективы его развития // Труды Казах. научно-исслед. института лесного хозяйства. Т. 5. Вып. 4. Полезащитное лесоразведение в Казахстане и Зап. Сибири. С. 62–73.
Флинт В.Е., 1984. Врановые птицы: изучение и регулирование численности // Экология, биоценотическое и хозяйственное значение врановых птиц. М.: Наука. С. 3–8.
Флинт В.Е., Сорокин А.Г., 1999. Сокол на перчатке. М.: Эгмонт Россия Лтд. 328 с.
Формозов А.Н., 1934. Озерная лесостепь и степь Западной Сибири как области массового обитания водяных птиц (Эколого-географический очерк) // Бюллетень Московского общества испытателей природы. Нов. сер. Отд. биол. Т. 43. Вып. 2. С. 256–286.
Формозов А.Н., 1937. Материалы к экологии водяных птиц по наблюдениям на озерах Государственного Наурзумского заповедника (Северный Казахстан) // Памяти академика М.А. Мензбира. М.–Л.: Изд-во Академии наук СССР. С. 551–595.
Формозов А.Н., 1940. Практическое значение птиц // Руководство по зоологии. Т. 6: Птицы. М.–Л.: Изд-во Академии наук СССР. С. 762–801.
Формозов А.Н., 1962. Изменения природных условий степного Юга Европейской части СССР за последние сто лет и некоторые черты современной фауны степей // Исследование географии природных ресурсов животного и растительного мира. М.: Изд-во Академии наук СССР. С. 14–161.
Хохлов А.Н., 1989. Современное состояние фауны гусеобразных Ставропольского края // Орнитологические ресурсы Северного Кавказа: Тезисы докладов науч.-практ. конф. Ставрополь. С. 106–136.
Цапко Н.В., 2005. Гнездование колониальных птиц в долине озера Маныч // Стрепет. Т. 3. Вып. 1–2. С. 88–93.
Цапко Н.В., 2007. Авифаунистические находки в Калмыкии // Птицы Кавказа: изучение, охрана и рац. использование: Материалы науч.-практ. конф., посвященной 25-летней деятельности Северокавказской орнитол. группы. Ставрополь. С. 136–139.
Цапко Н.В., Бобенко О.А., 2012. Первые гнездовые находки белощекой крачки и хохлатой чернети в Ставропольском крае // Кавказский орнитологический вестник. Вып. 24. Ставрополь. С. 119–123.
Чельцов-Бебутов А.М., 1967. О путях использования водоплавающей дичи (спорт или заготовки?) // Орнитология. Вып. 8. С. 47–66.
Червона книга України: Тваринний свiт, 2009. Київ: Глобалконсалтинг. 600 с.
Шевченко В.Л., Дебело П.В., Гаврилов Э.И., Наглов В.А., Федосенко А.К., 1993. Об орнитофауне Волжско-Уральского междуречья // Фауна и биология птиц Казахстана. Алматы: Наука КазССР. С. 7–103.
Шевченко В.Л., Дебело П.В., Гаврилов Э.И., Наглов В.А., Федосенко А.К., 2020. Об орнитофауне Волжско-Уральского междуречья (окончание) // Selevinia. Т. 28. С. 120–182.
Шуков П.М., 2018. О гнездовании клинтуха Columba oenas на опорах высоковольтных ЛЭП в Нижегородской области // Русский орнитологический журнал. Т. 27. № 1709. С. 6103–6105.
Янушевич А.И., Золотарева О.С., 1947. Водоплавающая дичь Барабы. Новосибирск. 79 с.
Bragin E., 2000. On the demography of the Imperial Eagle Aquila heliaca in Kazakhstan // Raptors at Risk. Berlin. P. 409–413.
del Hoyo J., Collar N.J. (eds), 2014. HBW and BirdLife International illustrated checklist of the Birds of the World. Vol. 1: Non-passerines. Barcelona: Lynx Ed. 903 p.
European breeding bird atlas 2: Distribution, abundance and change, 2020. Barcelona: European Bird Census Council & Lynx Edicions. 967 p.
Fox A.D., Leafloor J.O. (eds), 2018. A global audit of the status and trends of Arctic and Northern Hemisphere goose populations (Component 2: Population accounts). Akureyri, Iceland: CAFF International Secretariat. 173 p.
Horváth M., Szitta T., Firmánszky G., Solti B., Kovács A., Moskát C., 2010. Spatial variation in prey composition and its possible effect on reproductive success in an expanding eastern Imperial Eagle (Aquila heliaca) population // Acta Zoologica Academiae Scientiarum Hungaricae. V. 56. № 2. P. 187–200.
Zykov C.D., Ulitin A.A., 2000. Waterfowl resources in Russia: numbers estimate and harvesting on hunting grounds of the Rosokhotrybolovsoyuz // Kaзapкa. № 6. C. 19–38.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал