

Зоологический журнал, 2022, T. 101, № 4, стр. 451-460
Сезонные миграции и флуктуации численности ушастой совы (Asio Otus, Strigiformes, Strigidae) по данным 60-летнего отлова и кольцевания в Восточной Прибалтике
В. А. Паевский a, *, А. П. Шаповал b, **
a Зоологический институт РАН
199034 Санкт-Петербург, Россия
b Биостанция “Рыбачий” Зоологического института РАН
238535 Рыбачий, Россия
* E-mail: payevsky@yandex.ru
** E-mail: apshap@mail.ru
Поступила в редакцию 17.01.2021
После доработки 27.04.2021
Принята к публикации 28.04.2021
- EDN: UKVVGB
- DOI: 10.31857/S0044513422020076
Аннотация
По результатам отлова и кольцевания на Куршской косе в Калининградской обл. ушастой совы в количестве 3391 особи в течение 1957‒2016 гг. изучены динамика численности, пути миграции и места зимовки, возрастная структура и уровень гибели птиц. Общая эффективность кольцевания составила 3.01%, но с течением времени количество возвратов колец достоверно снижалось. Ежегодное количество отловленных сов в течение 60 лет резко колебалось, от одной до 365 особей. Колебания имели волнообразный характер с многочисленными волнами разной амплитуды. В связи с этим обсуждены известные положения о том, что численность ушастых сов и степень их территориальной мобильности зависят от обилия основного объекта питания – полевок. Среди мигрирующих сов преобладали молодые неполовозрелые особи (83%) и самки (65%), среди найденных по возвратам колец – тоже молодые, до 66%, а рекорд долгожительства – 16 лет и 3 месяца. Из всех 102 возвратов колец только 16 птиц были обнаружены живыми. Основное направление перелетов – юго-западное. Наибольшее количество возвратов колец, свидетельствующих о местах зимовок и пролетных путях, пришло из Германии и соседних стран на широтах 50°–54° N. Ряд возвратов колец свидетельствует о популяционном происхождении ушастых сов, мигрирующих через Куршскую косу, поскольку впоследствии они были найдены в Литве, Латвии, Белоруссии, Финляндии, а также на разных территориях России – от Ленинградской области до Татарстана.
Ушастая сова – обычный вид сов, распространенный в умеренных зонах Евразии и Северной Америки. Будучи наиболее многочисленной из сов Палеарктики, ушастая сова в своем питании зависит на всем пространстве ареала от плотности популяций мышевидных грызунов, в первую очередь полевок рода Microtus. Известно, что в годы вспышек численности полевок ушастые совы приступают к гнездованию в гораздо большем числе, чем обычно (Korpimäki, Norrdahl, 1991; Korpimäki, 1992; Приклонский, Иванчев, 1993; Houston, 2005; Волков и др., 2009, 2009а; Saurola, 2014). В ряде сводок о жизни сов указывается, что ушастая сова во многих частях ареала оседлая птица, а в осенне-зимнее время чаще всего ведет кочевой образ жизни. Северные популяции являются настоящими перелетными птицами. Однако в годы очень большой численности полевок на местах размножения они не совершают далеких передвижений (Дементьев, 1951; Eck, Busse, 1973; Пукинский, 1977; Сramp, 1985).
Ушастую сову, наряду с другими видами птиц, регулярно отлавливают и кольцуют на Куршской косе (Восточная Прибалтика) сотрудники Биологической станции “Рыбачий” Зоологического института РАН. Полученные сведения о миграциях птиц, в том числе и ушастой совы, с картами их передвижений по результатам первого десятилетия работы были опубликованы (Паевский, 1971). Впоследствии были опубликованы также материалы по численности этого вида на пролете и основным направлениям его миграций до 1985 г., в сравнении с данными мечения в других регионах (Белопольский, 1975; Сапетина, 1991; Dobrynina, 1994). За последующие 35 лет получены новые сведения по колебаниям ежегодного количества мигрирующих сов, результатам их прижизненного обследования и картине территориального распределения мигрантов в Европе. Эти новые материалы послужили побудительной причиной написания данной статьи.
МАТЕРИАЛ И МЕТОДИКА
Куршская коса – узкая полоса суши, отделяющая Куршский залив от Балтийского моря. Коса вытянута в направлении c северо-востока на юго-запад, что совпадает с основным направлением перелетов птиц в Восточной Прибалтике. Отлов и кольцевание птиц коллективом Биологической станции “Рыбачий” ЗИН РАН проводится на косе с 1957 г. до настоящего времени в двух местах: на полевом стационаре “Фрингилла” (55°05′ N, 20°44′ E) и на Росситенском мысу (55°09′ N, 20°51′ E). В первом из них ловят птиц в Рыбачинские ловушки, во втором – паутинными сетями. Рыбачинские ловушки для отлова мигрирующих птиц, их устройство и механизм действия подробно описаны (Дольник, Паевский, 1976). Ловушки действуют на протяжении 7 месяцев года, с конца марта по начало ноября. Работая круглосуточно, ловушки эффективно ловят сов в темное время суток. Сроки действия каждой ловушки (неизменной конструкции) варьировали по годам крайне незначительно, за счет нескольких дней начала весеннего и окончания осеннего отлова. Отлов и кольцевание птиц сопровождается их прижизненным обследованием, определением пола и возраста, а также стандартными измерениями длины крыла и массы тела. Возраст и пол ушастых сов стали определять, начиная с 1991 г., по характеристикам окраски оперения (Holt, 2016; Demongin, 2016).
Списки всех количественных данных отлова, кольцевания и возвратов колец опубликованы (Паевский, 1971; Bolshakov et al., 1999, 2001, 2005, 2008; Шаповал и др., 2017). В этих публикациях при сведениях о возвратах приведены координаты точек нахождения, время, прошедшее со дня кольцевания, расстояние и азимут. Эти данные были нами использованы для анализа территориального распределения мигрантов.
Тренды численности были рассчитаны регрессионным анализом по рядам динамики, где один ряд переменных – показатели времени (месяцы или годы), а другой – количество пойманных птиц. Расчет эффективности метода кольцевания проведен с использованием рангового коэффициента Спирмена, а анализ территориального распределения разных групп мигрирующих сов по возвратам колец – критерием χ2 и критерием Вилкоксона (Холлендер, Вулф, 1983; Sokal, Rohlf, 1998).
РЕЗУЛЬТАТЫ
Эффективность кольцевания. Всего за 60 лет, с 1957 по 2016 гг., на Куршской косе было поймана и окольцована 3391особь ушастой совы, и получены 102 “возврата” наших колец, т.е. сообщений об обнаружении окольцованных сов, позволяющих судить о дальности и направлении миграционных передвижений, а также о продолжительности их жизни. Полученные 102 сообщения о находках ушастых сов с нашими кольцами составляют 3.01 ± 0.01% от 3391 окольцованной особи. Расчет зависимости количества возвратов колец от количества окольцованных птиц был проведен по пятилетним периодам, поскольку были годы с нулевым количеством возвратов колец. Результат по ранговому коэффициенту Спирмена показал умеренную связь (rs = – 0.641, n = 12, p < 0.01), т.е. с ходом времени количество возвратов колец достоверно снижалось. В графическом виде эта зависимость более очевидна (рис. 1).

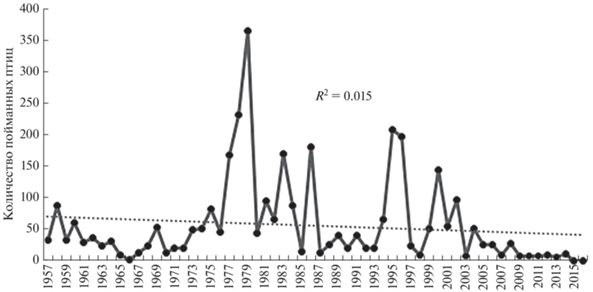
Динамика численности мигрантов. Ежегодное количество отловленных на Куршской косе ушастых сов в течение 60 лет резко колебалось, от одной до 365 особей. Эти изменения были волнообразными с разной амплитудой (рис. 2). Несмотря на огромные межгодовые различия количества пойманных сов, коэффициент детерминации R2 = 0.02 свидетельствует об отсутствии достоверного долговременного изменения численности.
Рис. 2.
Количество пойманных и окольцованных ушастых сов на Куршской косе в разные годы за 60 лет, 1957–2016 гг.
Возрастной и половой состав мигрантов. Размеры длины крыла самцов и самок ушастой совы в значительной степени перекрываются, что не дало возможности определять половой состав по длине крыла у всех особей. Длина крыла варьировала от 259 до 317 мм, но при этом птицы с длиной крыла от 287 до 299 мм могли быть любого пола. Поскольку возраст и пол ушастых сов на Куршской косе по признакам окраски оперения начали определять с 1991 г., мы располагаем материалами по возрастно-половому составу только для 1115 птиц. Из них во время миграций в 1991‒2016 гг. преобладали молодые неполовозрелые особи (83.1 ± 1.2%) и самки (65.1 ± 1.5%). В начале осенней миграции, во второй половине сентября, летели преимущественно молодые птицы, а взрослые появлялись только с октября.
По возвратам колец можно рассчитать приблизительное соотношение возраста сов в годовом выражении, используя даже особей, возраст которых в момент кольцевания был неизвестен, поскольку в среднем более 80% среди кольцуемых были молодые особи. Рассчитав долю обнаруженных с кольцами сов в возрасте одного года и более (1+), двух лет и более (2+), трех лет и более (3+) и т.д., мы таким образом получили возрастную структуру (рис. 3). Выяснилось, что в руки человека попадают тоже преимущественно молодые совы, до 65.7% от числа обнаруженных. Максимальная продолжительность жизни составила минимум 16 лет и 3 месяца.

Миграционные передвижения, районы зимовки и обстоятельства находок окольцованных сов. Помесячное распределение количества пойманных на Куршской косе ушастых сов показывает (рис. 4), что пик их миграции осенью приходится на октябрь и начало ноября. Весенняя миграция выражена слабо, небольшой подъем количества сов приходится на апрель и май. Количество полученных возвратов колец более или менее равномерно распределено в течение года, за исключением снижения в летние месяцы. Периоды кольцевания сов оказались значительно более узкие, нежели периоды возвратов колец.
Рис. 4.
Количество особей ушастой совы, пойманных и окольцованных в разные месяцы года (кривая), и количество особей, обнаруженных в разные месяцы на территориях пролета и зимовки (гистограмма).

Из всех 102 обнаруженных сов с нашими кольцами только 16 птиц (15.7%) были обнаружены живыми (из них 9 пойманы и выпущены, а 7 пойманы, но об их дальнейшей судьбе не сообщено). Среди остальных сов 62 особи (60.8%) найдены мертвыми или умирающими, включая убитых хищником, а также останки птиц с кольцами, 12 (11.7%) были убиты человеком и 12 (11.7%) стали жертвами столкновений с транспортом.
Из общего количества возвратов колец часть (n = 45) относилась к тому же сезону, что и сезон кольцевания (это так называемые «прямые» возвраты), с пределами разброса расстояния перелета от 54 до 1774 км, другая часть («непрямые» возвраты, n = 57) – к последующим сезонам, с пределами от 47 до 1790 км. Средние расстояния миграций по прямым возвратам колец составили 820.0 ± 64.2 км, по непрямым 833.9 ± 57.7 км, достоверных различий между ними не было (χ2 = 58.9, df = 44, n.s.).
Азимут разлёта варьировал от 22° до 355° (в среднем 239.5° ± 12.4°) у прямых возвратов и от 18° до 322° (в среднем 211.7° ± 10.1°) у непрямых, без достоверных различий (χ2 =1.12, df = 44, n.s.). Следовательно, средний пеленг точек обнаружения большинства сов находился к юго-западу от места нашего кольцевания. Тем не менее некоторые особи двигались и к югу, и к северо-западу, перелетая даже над океаном.
Расположение всех точек находок окольцованных ушастых сов приведено на рис. 5. Распределение их по странам Западной и Восточной Европы отражает разную степень концентрации на миграционных путях и территориях зимовок. Наибольшее количество возвратов колец – из Германии (33), России (13), Польши (9), Бельгии (8), Нидерландов (8), Франции (7) и Финляндии (6). Наибольшее расстояние перелета – до 1774 км. Возвраты колец с территорий, находящихся севернее и восточнее Куршской косы (15 птиц), предположительно говорят о широком популяционном происхождении птиц, мигрирующих через Куршскую косу, а именно – из Литвы, Латвии, Белоруссии, Финляндии, а также из разных областей России – Ленинградской, Тверской, Рязанской, Московской и из Татарстана. При предполагаемом возвращении сов на места размножения этим же путем, угол их разлёта на места размножения может достигать от 10° до 96°, а расстояние – до 1790 км.
Рис. 5.
Распределение ушастых сов, окольцованных на Куршской косе, на миграционных путях, местах зимовок и территориях размножения: 1– место кольцевания, 2 – точки обнаружения на путях миграций и местах зимовок, 3 – точки обнаружения на местах возможного гнездования.

Свидетельством кочевого образа жизни ушастых сов в областях зимовок может быть разная протяженность миграций по данным возвратов колец в разные месяцы года. Сроки нахождения ушастых сов в Европе по нашим данным приходятся на все месяцы года, но чаще их обнаруживают с октября по конец мая. На рис. 6 приведены расстояния от места кольцевания до точек обнаружения ушастых сов в разные месяцы. С октября по март включительно эти дистанции постепенно увеличиваются, в среднем от 671 до 1027 км. Попарное сравнение дистанций (октябрь ‒ ноябрь, ноябрь ‒ декабрь и т.д.) по их центральной тенденции непараметрическим критерием Вилкоксона во всех случаях показало отсутствие достоверных различий, и лишь сравнение дистанций в октябре с дистанциями в марте находилось на границе достоверности различия (медианы 642 и 1087 км, d.f. = 9, nx = 78, p < 0.05). Величины азимутов точек прямых возвратов в октябре-ноябре (в среднем 245°) и январе-феврале (в среднем 240°) достоверно не различались (χ2 = 3.2, d.f. = 7, n.s.). Таким образом, в целом небольшое увеличение протяженности миграций, начиная с октября, может говорить и о перемещениях сов в течение зимы, но достоверного заключения об этом сделать невозможно. В апреле и мае некоторые совы находились еще в странах Западной Европы, а не на местах размножения. Связана ли такая задержка на миграционных путях с возрастом птиц и сроками прихода их в половозрелое состояние, или же это объясняется другими причинами, пока остаётся неизвестным.
Рис. 6.
Расстояние от места кольцевания на Куршской косе до мест обнаружения окольцованных ушастых сов в разные месяцы, октябрь–март. Вертикальные линии – размах значений в километрах, внутри прямоугольника содержится половина значений, крестиком обозначена средняя арифметическая, горизонтальной линией – медиана.

Суточная скорость перелёта. Достоверную скорость передвижения ушастых сов по миграционным путям мы смогли определить лишь у восьми птиц, которые были обнаружены после кольцевания спустя 7–40 суток. Две из них окольцованы во время весенней миграции, остальные – во время осенней. Скорость передвижения варьировала от 21.6 до 58.2 км, составив в среднем 36.4 км в сутки.
ОБСУЖДЕНИЕ
Сравнительная результативность кольцевания ушастых сов. Результаты кольцевания на Куршской косе пролетных особей сов в виде полученных возвратов колец составили 3% от количества окольцованных. Такие результаты оказались более успешными по сравнению с результативностью кольцевания мелких певчих птиц, часто не превышающими даже 1%. Ранее мы уже проверяли эту зависимость на разных видах птиц и выяснили, что несмотря на фактическое снижение доли возвратов, во многих случаях достоверная корреляция между количеством окольцованных и количеством возвратов отсутствовала. По-видимому, погодные и иные условия года могут существенно влиять на этот показатель. Эффективность кольцевания ушастой совы до середины 1990-х гг. в разных европейских странах варьировала от 0.5 до 10.1%, в среднем 5.1% (Payevsky, Shapoval, 1998). В наше время в Финляндии результативность кольцевания ушастой совы тоже достигает 5.5%, хотя, в отличие от наших данных по кольцеванию мигрирующих, там более половины особей были окольцованы в возрасте птенцов на гнездах (Saurola, 2014). Известно, что начиная с восьмидесятых годов прошлого века в орнитологической литературе появилась информация о значительном снижении доли возвратов колец всех видов птиц по сравнению с предыдущими десятилетиями. По-видимому, изменение отношения людей к необходимости сообщить об окольцованной птице– это важная причина сокращения доли возвратов колец (Паевский, Шаповал, 2013).
Вопросы цикличности численности мигрантов. Резкие ежегодные флуктуации количества пойманных ушастых сов неизбежно обусловили появление вопросов относительно причин этого явления и связи его с особенностями жизненного цикла сов. Существует ли цикличность популяционных размеров этого вида? Материалы отлова ушастых сов на территориях других частей Прибалтики, откуда птицы могли прибывать на Куршскую косу, тоже свидетельствуют об огромных межгодовых колебаниях количества пойманных ушастых сов. В латвийском пункте кольцевания Папе размах колебаний был от одной до 960 особей (Граубиц, 1976; Руте, Бауманис, 1986), а в польском пункте Буково-Копань – от 8 до 736 (Busse, Busse, 2003; Michalonek et al., 2005). Сравнение данных из разных пунктов кольцевания показывает отсутствие синхронности в динамике отловов, однако на нашем и латвийском стационарах период с 1975 по 1979 годы отличался наиболее высокой численностью ушастой совы, тогда как в Польше таким периодом были 2000‒2002 гг. (Michalonek et al., 2005). В Северной Америке пики массовых миграционных передвижений ушастой совы могли указывать как на 10-летние, так и на 3-летние циклы численности. При этом разнообразие и направлений, и расстояний передвижения сов в Канаде и США могло свидетельствовать о кочевом образе жизни в связи с неустойчивой пищевой базой (Houston, 2005).
Известно, что численность ушастых сов и степень их территориальной мобильности целиком зависят от обилия основной пищи – полевок, резко меняющегося в разные годы, и при этом асинхронно в разных областях. Количество размножающихся сов и их перераспределение на гнездовании достоверно зависит от численности полевок, причем как непосредственно перед началом размножения, так и предшествующей осенью, а в годы их отсутствия совы вовсе не приступают к размножению (Korpimäki, 1992, 1994; Волков и др., 2009). Некоторая часть популяций ушастых сов в пики численности полевок остаются зимовать на более северных территориях (Korpimäki, Norrdahl, 1991; Korpimäki, 1994). Более того, в последнее время зарегистрировано множество случаев зимнего и осенне-зимнего гнездования во многих местах Европы, от России до Италии и Великобритании (Морозов, Конторщиков, 2008; Noga, 2009; Храбрый, Байбекова, 2015). Вообще для этого вида характерны очень растянутые сроки начала кладки, летом даже до середины июня (Шаповал, 2013). Всё это еще раз подтверждает, что размножение ушастой совы происходит только при наличии достаточной пищевой базы. А численность осенью и зимой в разных регионах, по результатам многих исследований, колеблется от нескольких десятков до нескольких сотен птиц (Константинов и др., 1982; Шариков и др., 2002). Следовательно, и численность мигрирующих сов может претерпевать разнообразные изменения большого размаха. Именно это и отражают данные нашего многолетнего отлова на Куршской косе, тем не менее строгой цикличности в этих колебаниях нет.
Некоторые исследователи как в Европе, так и в Северной Америке, полагают, что численность популяций ушастых сов в последние десятилетия имеет тенденцию к сокращению (Houston, 2005; Носков, Лапшин, 2016). Однако данные по успешности размножения этого вида в разных регионах этого не подтверждают. Межгодовые колебания численности сов в Финляндии в 1986–2015 гг. были очень большого размаха, но тем не менее успешность размножения оставалась близкой к среднему многолетнему показателю. Среднее количество птенцов на гнездо с успешным размножением составляло 2.94 (Björklund et al., 2015). Практически такое же (3.0) количество птенцов на успешное гнездо отмечено и в другом регионе Европы – в Белоруссии (Ивановский, 2015).
Соотношение возрастно-половых групп среди мигрирующих ушастых сов. Достоверное преобладание молодых птиц (83%) и самок (65%) среди пойманных нами мигрантов предположительно может указывать на пониженный миграционный стимул у старых самцов, хотя у нас нет подтверждающих данных. На балтийском побережье Польши в 1996‒2003 гг. среди 2044 пойманных мигрирующих ушастых сов тоже преобладали молодые птицы, 89.4 ± 0.7% (Michalonek et al., 2005). Несмотря на вариации показателей в разные годы, эти весьма сходные величины средних долей у нас и в Польше (83 и 89%) свидетельствуют, во-первых, о надежности полученных орнитологами сведений о возрастном составе перелетных сов и, во-вторых, о возможных для Восточной и Южной Прибалтики общих территориальных источниках мигрирующих популяций.
Известно, что последовательность осеннего перелета разных возрастно-половых групп птиц имеет свои особенности. По данным разных исследований, у ряда видов птиц осенью первыми улетают молодые особи обоих полов, а также самки разного возраста, а доля самцов и взрослых птиц постепенно увеличивается к концу пролёта (Паевский, 1985). У ушастой совы, по нашим данным, в самом начале перелета, во второй половине сентября, летели только молодые птицы, а взрослые появлялись с первых чисел октября. Однако по результатам отлова в Польше отмечалась обратная последовательность – взрослые совы мигрировали немного раньше молодых – в разные годы на 1–16 сут ранее (Michalonek et al., 2005).
По данным возвратов наших колец оказалось, что в руки человека попадают тоже преимущественно молодые совы, до 65.7% от числа всех обнаруженных. Весьма похожие данные соотношения разных возрастов по возвратам колец приведены и по результатам кольцевания в Финляндии. Сов на первом году жизни там было 57.8% (Saurola, 2014). Что же касается максимальной продолжительности жизни этого вида птиц, то и здесь показатели из разных стран и континентов оказываются очень сходными: у нас 16 лет и 3 месяца, в Финляндии 17 лет и 8 месяцев (Saurola, 2014), в Северной Америке 15 лет и 8 месяцев (Houston, 2005). По данным обзора о максимальной продолжительности жизни птиц, среднее значение для всех видов птиц из отряда совообразных составило 15 лет (Wasser, Sherman, 2010).
Причины смертности ушастых сов, их миграционные передвижения и основные места зимовки. Полученные возвраты наших колец оказались более или менее равномерно распределенными по месяцам в течение года, за исключением почти полного снижения в летние месяцы. Примерно такое же распределение по месяцам года этих показателей существует и по опубликованным данным из Финляндии (Saurola, 2014). Отличие от показателей из Финляндии оказалось в том, что по нашим данным живыми были обнаружены только около 16% сов от числа возвратов колец, тогда как по материалам из Финляндии – 29% от 756 возвратов колец (Saurola, 2014). Из всех разнообразных причин гибели окольцованных сов (прямое убийство человеком, столкновение с транспортом, жертва других хищных птиц и т.п.) наибольшая доля, 56%, остается неизвестной, поскольку эти птицы с нашими кольцами были найдены мертвыми или умирающими.
Сообщения о находках окольцованных птиц широко используются для составления карт миграционных путей и мест зимовок. Однако в ряде случаев существуют некоторые сомнения в правильности полученной таким образом картины распределения мигрантов. Сомнения проистекают из того очевидного обстоятельства, что количество возвратов колец зависит от плотности поселений человека и его культурного уровня. Один из основных факторов, искажающих полученные сведения, – концентрация охотников и птицеловов в некоторых местах Европы (Паевский, 1973; Busse, 2001). Тем не менее для сов, проявляющих, в отличие от многих других птиц, ночную активность, это обстоятельство может иметь не столь важное значение.
Распределение мигрирующих сов по Европе, приведенное на карте (рис. 5), с несомненностью говорит о том, что часть территории Европы, ограниченная широтой 50°–54° N и долготой 1°–16° E, представляет собой основное место, куда устремляются ушастые совы, мигрирующие через Куршскую косу, т.е. птицы из Прибалтики, Ленинградской, Тверской, Московской и Рязанской областей. Зимуют они в основном в северной Германии, Нидерландах и Бельгии. Однако отдельные птицы обнаруживаются на огромной территории, от Италии до Норвегии. Места зимовок ушастых сов из Финляндии тоже занимают весьма далекие от мест размножения регионы, от Норвегии и Британских о‒вов до Белоруссии и Украины, хотя медиана мест их зимнего нахождения находится в Дании (Saurola, 2014).
Скорость перелетов по данным возвратов колец. Изучение скорости миграционных передвижений птиц – один из необходимых аспектов детальных исследований их сезонных перелетов. Более или менее протяженные миграции в типичном случае включают несколько циклов полета, остановок и стартов для следующих полетов. Поведение мигрантов во время полета и на остановках определяется их видоспецифичным кормовым поведением, погодными и биотопическими условиями. Полученная нами средняя скорость передвижения сов, равная 36.4 км в сутки, вполне реальна по сравнению с другими видами птиц. У большинства видов птиц значения средней суточной скорости передвижения находятся в пределах от 20 до 100 км (Паевский, 2012). Разумеется, данные, полученные с помощью телеметрии, в первую очередь спутниковой, по сравнению с кольцеванием в ряде случаев принципиально меняют традиционные представления и о путях миграции, и о ее скорости. При этом достоверность таких данных все же зависит от принятого условия, что прикрепление к птице передатчика, как и кольца, не изменяет ее миграционное поведение.
ЗАКЛЮЧЕНИЕ
Обобщение всех данных кольцевания ушастых сов в течение шести десятилетий позволило выявить ряд особенностей их миграционных передвижений. Во-первых, существует весьма резко выраженная годовая нестабильность массового пролета, которая наблюдается и в других местах обитания вида и связана с цикличностью численности основного объекта питания – мышевидных грызунов, прежде всего полёвок. Во-вторых, несмотря на основное юго-западное направление осенних перемещений после пролета по Восточной Прибалтике и преимущественную зимовку в Германии, Бельгии, Нидерландах, на широтах 50°–54° N, наблюдается очень широкое распределение отдельных мигрирующих птиц по Европе, от Италии и Хорватии до Швеции, Норвегии и Оркнейских о-вов в Шотландии. В-третьих, огромные расстояния, по расчетам более 2–3 тысяч километров, преодолеваемые некоторыми птицами от мест размножения до мест зимнего пребывания. В-четвертых, возрастной состав мигрирующих сов характеризуется преобладанием молодых особей и по нашим данным, и по данным из разных мест кольцевания в Европе и в Северной Америке, а также сходной максимальной продолжительностью жизни отдельных птиц, от 15 до почти 18 лет.
Список литературы
Белопольский Л.О., 1975. Характеристика миграций некоторых сов (Strigiformes)по данным их отлова и кольцевания на Куршской косе в 1957–1968 гг. // Сообщения Прибалтийской комиссии по изучению миграций птиц. № 8. С. 51–71.
Волков С.В., Шариков А.В., Басова В.Б., Гринченко О.С., 2009. Влияние обилия мелких млекопитающих на выбор местообитаний и динамику численности ушастой (Asio otus) и болотной (Asio flammeus) сов // Зоологический журнал. Т. 88. № 10. С. 1248–1257.
Волков С.В., Шариков А.В., Морозов В.В. (Ред.), 2009а. Совы Северной Евразии: экология, пространственное и биотопическое распределение. М. 304 с.
Граубиц Г., 1976. Новое в отлове сов во время осенней миграции в Папе // Материалы IX Прибалтийской орнитологической конференции. Вильнюс. С. 76–80.
Дементьев Г.П., 1951. Отряд совы // Птицы Советского Союза. Т. 1. М.: Наука. С. 342–429.
Дольник В.Р., Паевский В.А., 1976. Рыбачинская ловушка // Кольцевание в изучении миграций птиц фауны СССР. М.: Наука. С. 73–81.
Ивановский В.В., 2015. Размножение ушастой совы Asio otus в Витебской области в период высокой численности вида // Русский орнитологический журнал. Т. 24. Вып. 1217. С. 4201–4206.
Константинов В.М., Марголин В.А., Бабенко В.Г., 1982. Особенности экологии ушастой совы в антропогенных ландшафтах Центрального района Европейской части СССР // Гнездовая жизнь птиц. Пермь. С. 121–132.
Морозов Н.С., Конторщиков В.В., 2008. Зимние кладки у ушастых сов в Москве в 2008 г. // Орнитология. Т. 35. С. 137–142.
Носков Г.А., Лапшин Н.В., 2016. Ушастая сова Asio otus // Миграции птиц Северо-Запада России. Неворобьиные. Под ред. Носкова Г.А., Рымкевич Т.А., Гагинской А.Р. СПб.: Изд-во АНОЛА “Профессионал”. С. 530–532.
Паевский В.А., 1971. Атлас миграций птиц по данным кольцевания на Куршской косе // Экологические и физиологические аспекты перелетов птиц (Труды ЗИН АН СССР, т. 50). Л. С. 3–110.
Паевский В.А., 1973. Достоверность информации о путях миграций птиц по данным кольцевания // Экология. № 2. С. 98–100.
Паевский В.А., 1985. Демография птиц. Л.: Наука. 285 с.
Паевский В.А., 2012. Скорость миграционных передвижений птиц как адаптивное поведение // Журнал общей биологии. Т. 73. № 5. С. 360–375.
Паевский В.А., Шаповал А.П., 2013. Падение доли возвратов колец от окольцованных птиц на протяжении последних 50 лет: каковы причины этого явления? // Орнитология. Т. 38. С. 24–31.
Приклонский С.Г., Иванчев В.П., 1993. Ушастая сова // Птицы России и сопредельных регионов: Рябкообразные, Голубеобразные. Кукушкообразные, Совообразные. М.: Наука. С. 302–313.
Пукинский Ю.Б., 1977. Жизнь сов // Жизнь наших птиц и зверей. Вып. 1. Л.: Изд-во Ленинградского университета. 240 с.
Руте Ю.Я., Бауманис Я.А., 1986. Многолетняя осенняя динамика численности нерегулярных мигрантов в Папе (Латвия) // Кольцевание и мечение птиц в СССР. 1979–1982 годы. М.: Наука. С. 23–29.
Сапетина И.М., 1991. Итоги кольцевания ушастой совы (Asio otus) в европейской части СССР // Результаты кольцевания и мечения птиц: 1985 г. М.: Наука. С. 20–28.
Холлендер М., Вульф Д., 1983. Непараметрические методы статистики. М.: Финансы и статистика. 518 с.
Храбрый В.М., Байбекова С.А., 2015. Зимнее размножение ушастой совы Asio otus в Санкт-Петербурге // Русский орнитологический журнал. Т. 24. Вып. 1123. С. 1057–1062.
Шаповал А.П., 2013. Материалы по гнездованию ушастой совы Asio otus в окрестностях села Лазорки (запад Полтавской области) // Русский орнитологический журнал. Т. 22. Вып. 843. С. 288–290.
Шаповал А.П., Леоке Д.Ю., Зеленова Н.П., 2017. Результаты отлова и кольцевания птиц Биологической станцией “Рыбачий” на Куршской косе в 2014 г. // Русский орнитологический журнал. Т. 26. Вып. 1549. С. 5605‒5627.
Шариков А.В., Константинов В.М., Климов С.М., Лысенков Е.В. Маловичко Л.В., 2002. Распределение и численность ушастой совы Asio otus в антропогенных ландшафтах Европейской России // Русский орнитологический журнал. Т. 11. Вып. 176. С. 135–142.
Bolshakov C.V., Shapoval A.P., Zelenova N.P., 1999. Results of bird ringing by the Biological Station “Rybachy” on the Courish Spit in 1998 // Avian Ecology and Behaviour. V. 2. P. 105‒150.
Bolshakov C.V., Shapoval A.P., Zelenova N.P., 2001. Results of bird ringing by the Biological Station “Rybachy” on the Courish Spit: long-distance recoveries of birds ringed in 1956–1997. Part 1 // Avian Ecology and Behaviour, Suppl. 1. P. 1–126.
Bolshakov C.V., Shapoval A.P., Zelenova N.P., 2005. Results of bird ringing by the Biological Station “Rybachy” on the Courish Spit in 2004 // Avian Ecology and Behaviour. V. 13. P. 47–95.
Bolshakov C.V., Shapoval A.P., Zelenova N.P., 2008. Results of bird ringing by the Biological Station “Rybachy"on the Courish Spit in 2005 // Avian Ecology and Behaviour. V. 14. P. 49‒100.
Björklund H., Saurola P., Valkama J., 2015. Breeding and population trends of common raptors and owls in Finland in 2015 // Linnut-Vuosikirja. Vuosi 2015. P. 41–53.
Busse P., 2001. European passerine migration system – what is known and what is lacking // Ring. V. 23. № 1‒2. P. 3–6.
Busse P., Busse W., 2003. Autumn migration of owls along the Polish Baltic Sea coast 1961–1999 // Vogelwelt. B. 124. S. 281–284.
Cramp S., 1985. The Birds of the Western Palearctic. V. IV. Oxford University Press. 960 p.
Demongin L., 2016. Identification guide to birds in the hand. Published by L. Demongin. 392 p.
Dobrynina I.N., 1994. Seasonal movements of owls (Strigiformes) in the Baltic region based on ringing data // Ring. V. 16. № 1–2. C. 77–83.
Eck S., Busse H., 1973. Eulen. Die Neue Brehm-Bücherei. A. Ziemsen Verlag. Wittenberg Lutherstadt. 196 S.
Holt D.W., 2016. Sex Differences in Long-eared Owl Plumage Coloration // Journal of Raptor Research. V. 50. № 1. P. 60–69.
Houston C.S., 2005. Long-eared owls, Asio otus: A review of North American banding // Canadian Field-Naturalist. V. 119. № 3. P. 395‒402.
Korpimäki E., Norrdahl K., 1991. Numerical and functional responses of kestrels, short-eared owls, and long-eared owls to vole densities // Ecology. V. 72. P. 814–826.
Korpimäki E., 1992. Diet composition, prey choice, and breeding success of Long-eared Owl: effect of multiannual fluctuations in food abundance // Canadian Journal of Zoology. V. 70. P. 2373–2381.
Korpimäki E., 1994. Rapid or delayed tracking of multiannual vole cycles by avian predators? // Journal of Animal Ecology. V. 63. P. 619–628.
Michalonek D., Busse W., Lasecki R., 2005. Age structure of the Long-eared Owl (Asio otus) migration at Bukowo-Kopañ station (southern Baltic coast) in autumns 1996–2003 //Ring. V. 27. № 2. P. 145–157.
Noga M., 2009. Winter breeding of the Long-eared Owl (Asio otus) in South-Western Slovakia // Slovak Raptor Journal. V. 3. C. 61–62.
Payevsky V.A., Shapoval A.P., 1998. Ringing efficiency of birds depending on their species, sex, age, season, and place of ringing // Ornithologia. V. 28. P. 212–218.
Saurola P., 2014. Sarvipöllö Asio otus // The Finnish Bird Ringing Atlas. Vol. II. Finnish Museum of Natural History, Helsinki. P. 255–264.
Sokal R.R., Rohlf F.J., 1998. Biometry: the principles and practice of statistics in biological research. 3nd ed. New York: Freeman and Co. 887 p.
Wasser D., Sherman P., 2010. Avian longevities and their interpretation under evolutionary theories of senescence // Journal of Zoology. V. 280. P. 103–155.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал