

Зоологический журнал, 2023, T. 102, № 2, стр. 172-180
К вопросу о географической изменчивости морфологических признаков итальянской саранчи Calliptamus italicus italicus (L.) (Orthoptera, Acrididae)
К. Е. Гаранин a, *, М. Е. Черняховский a, **
a Институт биологии и химии,
Московский педагогический государственный университет
129164 Москва, Россия
* E-mail: kirik952@rambler.ru
** E-mail: mech41@yandex.ru
Поступила в редакцию 22.04.2022
После доработки 20.09.2022
Принята к публикации 22.09.2022
- EDN: HSVMMJ
- DOI: 10.31857/S004451342302006X
Аннотация
Описана клинальная изменчивость ряда морфологических признаков Calliptamus italicus italicus. Увеличение размеров с запада на восток и с севера на юг может быть объяснено влиянием климатических факторов, в основном температурой, числом теплых дней и количеством осадков в теплый период. Приведены данные о находке вида в Восточном Забайкалье за пределами известного ареала.
Размер тела прямокрылых нередко подвержен географической изменчивости как в широтном, так и в долготном направлении. При этом у разных видов ряды изменчивости одного и того же признака могут иметь противоположную направленность. Подобные различия в действии природных факторов на морфологические признаки привлекают внимание многих исследователей (Ciplak et al., 2008; Parsons, Joern, 2014).
В настоящее время существует несколько гипотез, объясняющих это явление (Brown et al., 2004; Whitman, 2008; Parsons, Joern, 2014). Одна из них связывает его с правилом Бергмана, сформулированным изначально для теплокровных животных. Действительно, размеры частей тела у некоторых прямокрылых увеличиваются с увеличением широты (Bidau, Marti, 2008; Ciplak et al., 2008). Однако у других видов наблюдается обратная закономерность – уменьшение размера тела с запада на восток. Некоторые авторы предполагают, что направление изменений может зависеть от размера тела и продолжительности развития конкретного вида: у мелких видов изменчивость соответствует правилу Бергмана, а крупные виды с более длительным временем развития демонстрируют противоположную тенденцию (Blanckenhorn, Demont, 2004).
Однако, как показали Леманн и Леманн (Lehmann, Lehmann, 2008), многие факторы, такие как продолжительность теплого сезона, плотность популяции, наличие кормовых ресурсов, местные патогены и межвидовая конкуренция, могут корректировать направление и размах клинальной изменчивости. Помимо этого, на формирование морфологических признаков влияют генетический полиморфизм и фенотипическая пластичность.
Олигер (1971) показал, что у Chortippus (Glyptobothrus) brunneus (Thunberg 1815) и Ch. (Gl.) mollis (Charpentier 1825) (Orthoptera, Acrididae) с севера на юг наблюдаются увеличение размеров тела и изменение формы надкрылья. Аналогичная закономерность была описана Крицкой (1972) для Chorthippus macrocerus (Fischer-Waldheim 1846), у которого с севера на юг происходит увеличение общей длины тела, длины переднеспинки, надкрылий, задних бедер и ширины медиального и кубитального полей надкрылий. Литвиновой (1972) установлены закономерности географической изменчивости ряда морфологических признаков Euchorthippus pulvinatus (Fischer-Waldheim 1846).
Calliptamus italicus (Linnaeus 1758) распространен в центре и на юге Европы, на юге европейской части России, Кипре, в Малой Азии, на Северном Кавказе, в Закавказье, Средней Азии, Сирии, Ираке, Иране, Афганистане, на юге западной Сибири, по всему Казахстану и в Северо-Западном Китае (Синьцзян) (Лачининский и др., 2002; Сергеев и др., 2016). На большей части ареала он представлен номинативным подвидом C. italicus italicus, который достаточно хорошо изучен вследствие своего практического значения.
Вполне закономерно, что в пределах столь обширного ареала у номинативного подвида проявляется географическая изменчивость. Для некоторых признаков это было показано в работах Ваньковой (2002, 2006) и Сергеева и др. (2016); например, в направлении с севера на юг в популяциях увеличивается доля особей с полосами на затылке, переднеспинке и на надкрыльях (Ванькова, 2006).
Цель настоящей работы – изучение географической изменчивости C. italicus italicus в широтном и долготном направлениях.
МАТЕРИАЛЫ И МЕТОДЫ
Для морфометрических исследований был использован материал, хранящийся в коллекции кафедры зоологии и экологии Московского педагогического государственного университета и в коллекции Зоологического музея Московского государственного университета им. М.В. Ломоносова. Изучено 428 особей из 7 точек: Республика Крым, окрестности города Белогорск, сборы В.С. Гусевой (44 самки, 59 самцов); Ростовская обл., окрестности Цимлянска, сборы Д.П. Довнар-Запольского (16 самок, 18 самцов); Ставропольский край, окрестности поселка Арзгир, сборы М.Е. Черняховского (46 самок, 43 самцов); Грузия, поселок Кулеви, сборы Е.Г. Банковской (26 самок, 56 самцов); Казахстан, Туркестанская обл., Ордабасынский р-н, окрестности поселка имени Исаханова, сборы М.Е. Черняховского (18 самок, 12 самцов); Таджикистан, заповедник Рамит, сборы М.Е. Черняховского (26 самок, 26 самцов); Забайкальский край, окрестности поселка Абагайтуй, сборы М.Е. Черняховского (24 самки, 12 самцов). Во всех случаях особи относились к одиночной форме.
Были использованы следующие показатели (рис. 1), обычно применяемые в систематике саранчовых (Uvarov, 1966): ELL – длина, ELW – ширина надкрылья; sEL – площадь надкрылья (ELL × × ELW), FEL – длина, FEW – ширина бедра; PRL – длина, PRW – ширина переднеспинки; iGR – индекс стадности (ELL/FEL), iFE – индекс бедра (FEL/FEW). Измерения проводились с помощью стереомикроскопа МБС-9, оснащенного окуляр-микрометром с ценой деления 0.1 мм.
Рис. 1.
Основные морфологические признаки особей из Восточного Забайкалья: A – самец; B – вариации формы церок самцов; C – самка, показаны промеры надкрылья; D – заднее бедро, изнутри, показаны промеры бедра; E – задняя нога, снаружи; ELL – длина надкрылья; ELW – ширина надкрылья; FEL – длина бедра; FEW – ширина бедра.

Статистическая обработка включала проверку нормальности распределения, оценку связи морфологических признаков с климатическими факторами (коэффициенты корреляции Пирсона и Спирмена) и проверку достоверности различий средних (критерии Стьюдента и Манна–Уитни). Совместное использование параметрических и непараметрических методов связано с тем, что распределение значений в ряде случаев (ширина переднеспинки у самцов из Крыма, Грузии, Ставропольского края, Таджикистана и Казахстана и индекс бедра у самцов из Крыма, Грузии, Таджикистана и Казахстана) значимо отличается от нормального (табл. 1).
Таблица 1.
Результаты проверки признаков тестом Колмогорова–Смирнова
| Точки сборов | Пол | Признаки | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| sEL | ELL | ELW | FEL | FEW | iFE | iGR | PRL | PRW | ||
| Крым | Самки | 0.08 | 0.09 | 0.18 | 0.09 | 0.18 | 0.07 | 0.11 | 0.11 | 0.17 |
| Самцы | 0.08 | 0.06 | 0.11 | 0.10 | 0.16 | 0.07 | 0.12 | 0.24 | 0.20 | |
| Ростов | Самки | 0.15 | 0.19 | 0.13 | 0.16 | 0.12 | 0.13 | 0.16 | 0.13 | 0.18 |
| Самцы | 0.17 | 0.14 | 0.15 | 0.15 | 0.18 | 0.21 | 0.15 | 0.20 | 0.14 | |
| Грузия | Самки | 0.19 | 0.14 | 0.09 | 0.14 | 0.10 | 0.14 | 0.11 | 0.21 | 0.13 |
| Самцы | 0.20 | 0.11 | 0.21 | 0.08 | 0.11 | 0.10 | 0.10 | 0.21 | 0.18 | |
| Ставропольский край | Самки | 0.06 | 0.07 | 0.09 | 0.09 | 0.13 | 0.13 | 0.13 | 0.09 | 0.19 |
| Самцы | 0.18 | 0.14 | 0.15 | 0.19 | 0.11 | 0.12 | 0.14 | 0.18 | 0.21 | |
| Таджикистан | Самки | 0.10 | 0.13 | 0.21 | 0.11 | 0.17 | 0.09 | 0.15 | 0.12 | 0.20 |
| Самцы | 0.16 | 0.15 | 0.10 | 0.12 | 0.12 | 0.11 | 0.17 | 0.13 | 0.21 | |
| Казахстан | Самки | 0.24 | 0.17 | 0.20 | 0.10 | 0.19 | 0.17 | 0.11 | 0.19 | 0.32 |
| Самцы | 0.25 | 0.25 | 0.23 | 0.14 | 0.16 | 0.14 | 0.18 | 0.17 | 0.23 | |
| Забайкальский край | Самки | 0.13 | 0.12 | 0.21 | 0.11 | 0.20 | 0.12 | 0.12 | 0.16 | 0.13 |
| Самцы | 0.18 | 0.16 | 0.26 | 0.19 | 0.34 | 0.2 | 0.23 | 0.15 | 0.24 | |
Для корреляционного анализа использовали следующие климатические показатели: средняя температура января, средняя температура июля, число дней с максимальной температурой воздуха выше 20°C, число дней с максимальной температурой воздуха выше 10°C, число дней без отрицательной температуры воздуха, число дней со снежным покровом за год, количество осадков в теплый период (мм), количество осадков за год (мм) (Атлас СССР, 1983).
Расчеты проведены в программе Statistica 10.0. В табл. 1–3, 5 серая заливка ячейки обозначает наличие статистической значимой связи (p < 0.05); белая заливка ячейки обозначает отсутствие статистически значимой связи (p > 0.05).
Таблица 2.
Клинальная изменчивость признаков Calliptamus italicus, оцененная как корреляция соответствующего признака и долготы/широты местообитания
| Географические координаты | sEL | ELL | ELW | FEL | FEW | iGR | iFE | PRL | PRW |
|---|---|---|---|---|---|---|---|---|---|
| Значение коэффициента корреляции Пирсона у самок | |||||||||
| Долгота | 0.28 | 0.11 | 0.23 | 0.18 | 0.28 | –0.05 | –0.15 | 0.26 | 0.09** |
| Широта | –0.47 | –0.45 | –0.39 | –0.48 | –0.47 | –0.17 | –0.05 | –0.37 | –0.10 |
| Географические координаты | Значение коэффициента корреляции Пирсона и Спирмена у самцов | ||||||||
| sEL | ELL | ELW | FEL | FEW | iGR | iFE* | PRL | PRW* | |
| Долгота | 0.20 | 0.23 | 0.36 | 0.41 | 0.36 | –0.08 | –0.01 | 0.40 | 0.16 |
| Широта | –0.31 | –0.37 | –0.39 | –0.47 | –0.43 | –0.14 | 0.06 | –0.54 | –0.18 |
Таблица 3.
Зависимость морфологических признаков Calliptamus italicus от климатических параметров
| Параметр | Значение коэффициента корреляции Пирсона и Спирмена у самцов | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| sEL | ELL | ELW | FEL | FEW | iGR | iFE* | PRL | PRW* | |
| A | –0.091 | –0.095 | –0.082 | –0.076 | –0.013 | –0.059 | –0.03 | –0.071 | 0.10 |
| B | 0.249 | 0.228 | 0.272 | 0.358 | 0.333 | 0.045 | 0.13 | 0.448 | –0.10 |
| C | 0.426 | 0.48 | 0.422 | 0.503 | 0.437 | 0.272 | –0.00 | 0.486 | 0.05 |
| D | 0.405 | 0.486 | 0.273 | 0.285 | 0.259 | 0.475 | –0.02 | 0.267 | 0.19 |
| E | 0.035 | –0.065 | 0.005 | 0.016 | 0.038 | –0.095 | –0.04 | 0.119 | 0.04 |
| F | –0.139 | –0.184 | –0.101 | –0.017 | 0.015 | –0.224 | –0.56 | 0.128 | –0.02 |
| G | –0.117 | –0.168 | –0.271 | –0.353 | –0.347 | 0.059 | –0.39 | –0.417 | 0.02 |
| H | 0.156 | 0.28 | 0.115 | 0.048 | 0.037 | 0.323 | –0.00 | –0.135 | –0.06 |
| Параметр | Значение коэффициента корреляции Пирсона у самок | ||||||||
| sEL | ELL | ELW | FEL | FEW | iGR | iFE* | PRL | PRW* | |
| A | –0.091 | –0.095 | –0.082 | –0.076 | –0.013 | –0.059 | –0.069 | –0.071 | 0.132 |
| B | 0.296 | 0.346 | 0.234 | 0.321 | 0.254 | 0.168 | 0.107 | 0.106 | –0.073 |
| C | 0.529 | 0.578 | 0.357 | 0.358 | 0.246 | 0.407 | 0.175 | 0.139 | –0.244 |
| D | 0.243 | 0.247 | 0.132 | 0.096 | 0.05 | 0.151 | 0.045 | –0.002 | –0.168 |
| E | –0.09 | –0.092 | –0.063 | –0.052 | –0.149 | –0.033 | 0.124 | –0.13 | 0.085 |
| F | –0.299 | –0.393 | –0.214 | –0.287 | –0.131 | –0.269 | –0.238 | –0.061 | 0.048 |
| G | –0.139 | –0.092 | –0.197 | –0.216 | –0.219 | 0.047 | 0.006 | –0.179 | –0.133 |
| H | 0.265 | 0.266 | 0.129 | –0.053 | –0.017 | 0.34 | –0.042 | –0.015 | –0.356 |
Примечания. * – признаки, для которых использовалась корреляция Спирмена. Параметры: A – среднемесячная температура января, B – среднемесячная температура июля, C – число дней в году с температурой больше 20°C, D – число дней в году с температурой больше 10°C, E – число дней в году с положительной температурой, F – число дней в году со снегом, G – количество осадков в теплый период, I – количество осадков за год. Остальные обозначение см. в тексте.
Таблица 4.
Среднее значение морфологических показателей Calliptamus italicus
| Точки сборов | Пол | sEL | ELL | ELW | FEL | FEW | PRL | PRW |
|---|---|---|---|---|---|---|---|---|
| Крым | Самцы | 91.9 ± 15.9 | 15.9 ± 1.8 | 2.9 ± 0.2 | 11.8 ± 0.9 | 3.5 ± 0.3 | 3.6 ± 0.3 | 3.1 ± 0.5 |
| Самки | 37.7 ± 7.4 | 24.0 ± 0.2 | 5.0 ± 0.6 | 18.7 ± 1.8 | 5.4 ± 0.5 | 6.1 ± 0.5 | 5.4 ± 1.2 | |
| Ростов | Самцы | 98.8 ± 11.7 | 16.4 ± 1.5 | 3.1 ± 0.3 | 11.5 ± 0.8 | 4.0 ± 0.3 | 2.8 ± 0.3 | 2.8 ± 0.3 |
| Самки | 51.1 ± 26.3 | 24.2 ± 2.7 | 4.3 ± 0.7 | 17.1 ± 2.0 | 4.7 ± 0.5 | 5.9 ± 0.6 | 4.3 ± 0.4 | |
| Грузия | Самцы | 151.4 ± 19.3 | 17.7 ± 2.9 | 3.2 ± 0.5 | 12.6 ± 0.7 | 3.9 ± 0.2 | 4.2 ± 0.2 | 3.1 ± 0.2 |
| Самки | 35.7 ± 2.9 | 26.9 ± 3.0 | 5.0 ± 0.4 | 19.3 ± 1.3 | 5.6 ± 0.4 | 6.7 ± 0.6 | 4.9 ± 0.4 | |
| Ставрополь | Самцы | 114.9 ± 23.0 | 17.6 ± 2.9 | 3.2 ± 0.5 | 12.6 ± 1.1 | 3.7 ± 0.4 | 3.8 ± 0.4 | 2.8 ± 0.4 |
| Самки | 47.9 ± 14.1 | 26.9 ± 3.0 | 5.3 ± 0.6 | 19.1 ± 1.6 | 5.4 ± 0.4 | 6.2 ± 0.4 | 3.9 ± 1.0 | |
| Таджикистан | Самцы | 115.7 ± 27.4 | 17.3 ± 3.4 | 3.1 ± 0.4 | 13.5 ± 1.3 | 4.2 ± 0.6 | 4.6 ± 0.8 | 3.9 ± 1.0 |
| Самки | 48.3 ± 16.3 | 26.9 ± 3.0 | 5.1 ± 0.7 | 19.5 ± 2.2 | 5.7 ± 0.6 | 6.8 ± 1.0 | 6.0 ± 1.7 | |
| Казахстан | Самцы | 117.1 ± 29.6 | 17.8 ± 3.5 | 3.3 ± 0.5 | 12.8 ± 1.2 | 3.7 ± 0.2 | 3.7 ± 0.2 | 3.7 ± 0.3 |
| Самки | 39.2 ± 15.4 | 25.8 ± 3.0 | 5.6 ± 1.0 | 18.8 ± 2.2 | 6.7 ± 0.8 | 5.2 ± 1.4 | 5.2 ± 1.5 | |
| Забайкалье | Самцы | 60.8 ± 11.1 | 10.8 ± 1.4 | 2.6 ± 0.3 | 10.9 ± 1.0 | 3.3 ± 0.3 | 3.3 ± 0.4 | 3.4 ± 0.2 |
| Самки | 21.7 ± 2.8 | 18.3 ± 1.8 | 4.4 ± 0.3 | 16.5 ± 0.9 | 5.0 ± 0.1 | 5.8 ± 0.9 | 5.9 ± 0.7 |
Таблица 5.
Результаты оценки критерием Стьюдента и Манна–Уитни особей Calliptamus italicus из разных популяций
| Признак | Самцы из сравниваемых популяций | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Казахстан/ Забайкалье | Забайкалье/ Ставрополь | Крым/ Забайкалье | Забайкалье/ Таджикистан | Забайкалье/ Ростов | Забайкалье/ Грузия | |||||||
| значение критерия | p | значение критерия | p | значение критерия | p | значение критерия | p | значение критерия | p | значение критерия | p | |
| sEL | 3.87 | 0.00 | –6.32 | 0.00 | 7.27 | 0.00 | –5.57 | 0.00 | –4.1 | 0.00 | –8.39 | 0.00 |
| ELL | 6.26 | 0.00 | –7.62 | 0.00 | 8.87 | 0.00 | –6.14 | 0.00 | –9.75 | 0.00 | –12.28 | 0.00 |
| ELW | 3.65 | 0.00 | –3.4 | 0.00 | 3.75 | 0.00 | –4.73 | 0.00 | –3.45 | 0.00 | –7.1 | 0.00 |
| FEL | 3.67 | 0.00 | –3.98 | 0.00 | 2.87 | 0.01 | –5.51 | 0.00 | –1.73 | 0.09 | –6.08 | 0.00 |
| FEW | 2.88 | 0.00 | –3.17 | 0.00 | 2.34 | 0.02 | –4.69 | 0.00 | –2.02 | 0.05 | –6.14 | 0.00 |
| PRL | 2.13 | 0.05 | –3.19 | 0.00 | 2.43 | 0.02 | –4.88 | 0.00 | –4.59 | 0.00 | –9.25 | 0.00 |
| PRW | 4* | 0.00 | 55* | 0.00 | 230* | 0.16 | 127* | 0.37 | 5.16 | 0.00 | 21* | 0.00 |
| Признак | Самки из сравниваемых популяций | |||||||||||
| Казахстан/ Забайкалье | Забайкалье/ Ставрополь | Крым/ Забайкалье | Забайкалье/ Таджикистан | Забайкалье/ Ростов | Забайкалье/ Грузия | |||||||
| значение критерия | p | значение критерия | p | значение критерия | p | значение критерия | p | значение критерия | p | значение критерия | p | |
| sEL | –10.85 | 0.00 | –8.52 | 0.00 | –10.63 | 0.00 | –6.89 | 0.00 | 8.47 | 0.00 | 9.09 | 0.00 |
| ELL | –12.81 | 0.00 | –10.03 | 0.00 | –15.94 | 0.00 | –8.14 | 0.00 | 11.75 | 0.00 | 9.12 | 0.00 |
| ELW | –5.92 | 0.00 | –4.95 | 0.00 | –5.63 | 0.00 | –0.83 | 0.41 | 3.68 | 0.00 | 7.54 | 0.00 |
| FEL | –7.23 | 0.00 | –4.59 | 0.00 | –8.48 | 0.00 | –1.16 | 0.25 | 5.52 | 0.00 | 6.93 | 0.00 |
| FEW | –3.98 | 0.00 | –1.56 | 0.13 | –6.28 | 0.00 | 1.83 | 0.08 | 3.33 | 0.00 | 4.9 | 0.00 |
| PRL | –1.85 | 0.07 | –2.64 | 0.01 | –4.07 | 0.00 | –0.16 | 0.87 | 0.79 | 0.43 | 3.25 | 0.00 |
| PRW | 9.87 | 0.00 | 2.06 | 0.05 | 6.21 | 0.00 | 7.76 | 0.00 | –1.81 | 0.07 | 0.27 | 0.79 |
РЕЗУЛЬТАТЫ
Географическую изменчивость оценивали как корреляцию широты или долготы местности с величиной признака. Этот анализ показал разную степень клинальной изменчивости изученных признаков (табл. 2).
Долготная изменчивость не обнаружена для ширины переднеспинки, индекса стадности и индекса бедра. Для одного из признаков (площадь надкрылья) обнаружена ложная корреляция. Анализ полной выборки показал, что у самцов и самок есть слабая, но значимая связь между площадью надкрылья и долготой (самцы r = 0.20 и самки r = 0.28, p < 0.05). Однако на графике (рис. 2A, 2C) видно, что это вызвано влиянием выборки из Забайкальского края. После процедуры цензурирование выборки, т.е. изъятия из нее забайкальских особей, получена иная картина: площадь надкрылья увеличивается с запада на восток, как показано на рис. 3В, 3D.
Рис. 2.
Долготная изменчивость (r – коэффициент корреляции Пирсона) площади надкрылья у итальянской саранчи: A, C – полная выборка; B, D – цензурированная (удалены особи Забайкальской точки); A, B – самцы; C, D – самки; p – уровень значимости; n – объем выборки; сплошная линия – линия регрессии, пунктирная линия – доверительный интервал.

Рис. 3.
Широтная изменчивость (r-коэффициент корреляции Пирсона) длины бедра у итальянской саранчи: A, B – самцы и самки полная выборка; p – уровень значимости; n – объем выборки; сплошная линия – линия регрессии; пунктирная линия – доверительный интервал.

Популяция C. italicus italicus из Восточного Забайкалья (окрестности поселка Абагайтуй) находится за пределами известного ареала этого вида. Данная находка значительно дополняет представления о его распространении, смещая границу ареала на восток более чем на 1500 км. По основным морфологическим признакам, а именно, окраске задних крыльев, форме и размеру надкрылий, форме церок самцов, изученные особи относятся именно к подвиду C. italicus italicus (рис. 1). Насекомые встречались на каменистых участках мелкосопочника, а также на выходах коренной породы с ксероморфной растительностью (эфедра, полынь, колючетравие); покрытие составило 60–70%, высота травостоя – порядка 50 см.
Широтная изменчивость прослеживается для всех показателей, кроме индекса бедра, а у самок еще и ширины переднеспинки; кроме того, у самцов не изменяется индекс стадности. При этом и у самцов и у самок с юга на север уменьшаются длина и ширина надкрылья, длина и ширина бедра, и длина переднеспинки (рис. 4).
Рис. 4.
Зависимость между климатическими факторами и длиной бедра у итальянской саранчи: r – коэффициент корреляции Пирсона; A, C – самцы; B, D – самки; A, B – среднемесячная температура июля; C, D – число дней с температурой выше 20°C; p – уровень значимости; n – объем выборки; сплошная линия – линия регрессии, пунктирная линия – доверительный интервал.
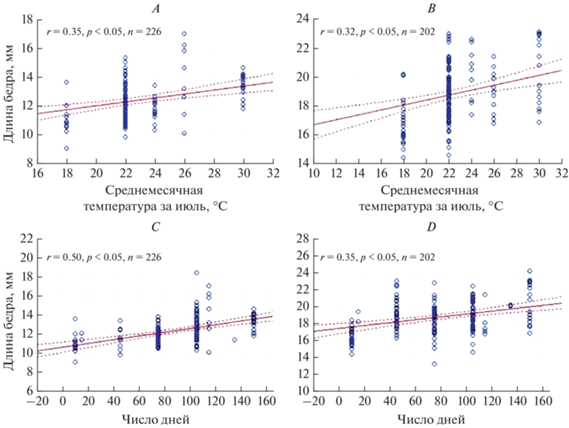
Очевидно, что выявленная географическая изменчивость имеет много причин, часть из которых – свойства местообитания, в какой-то мере определяемые широтой и долготой местности. Поэтому на следующем этапе анализа мы оценили зависимость признаков C. italicus italicus от параметров климата в районах сборов (табл. 3).
Ряд показателей (длина и ширина надкрылья, длина и ширина бедра у обоих полов; у самцов – еще и длина переднеспинки) положительно коррелирует со среднемесячной температурой в июле (рис. 4). Почти все морфологические показатели, кроме индекса бедра и ширины переднеспинки у самцов и длины переднеспинки у самок, положительно коррелируют с числом дней с температурой воздуха выше 20°C (табл. 3, рис. 4).
Обилие осадков в теплый период, напротив, отрицательно влияет на длину и ширину бедра и длину переднеспинки. Количество осадков за год положительно влияет только на индекс стадности.
Для многих показателей (средняя температура января, число дней с температурой воздуха больше 10°C, число дней с положительной температурой воздуха и число дней со снегопадом за год) не выявлено статистической значимого влияния на морфологические признаки (табл. 3).
Степень различий между выборками из разных популяций варьировала. Наиболее обособленной оказалась Забайкальская популяция, особи из которой практически по всем морфологическим признакам (площадь надкрылья, длина и ширина надкрылья и бедра) достоверно отличаются от особей из других точек (p < 0.05) (табл. 4, 5). Только при сравнении с выборкой из Ростовской обл. не выявлено статистически значимых различий между средними значениями длины и ширины бедра у самцов, а также длины и ширины надкрылья и бедра у самок (табл. 4, 5).
ОБСУЖДЕНИЕ
Таким образом, для номинативного подвида C. italicus italicus доказана клинальная изменчивость площади надкрылий, длины и ширины надкрылий и бедра, длины переднеспинки в широтном направлении. Аналогичную закономерность выявила Литвинова (1972) у Euchortippus pulvinatus, в популяциях которого с севера на юг увеличивается длина тела и переднеспинки. Увеличение размерных признаков в южных популяциях по сравнению с северными было описано и у ряда других саранчовых (Laiolo et al., 2013), а некоторые авторы считают это явление характерным для насекомых в целом (Арнольди, 1939).
Клинальная изменчивость в долготном направлении у C. italicus italicus менее выражена и выявлена только для длины и ширины заднего бедра и длины переднеспинки.
В большинстве случаев (длина и ширина надкрылья, длина и ширина бедра, а у самцов еще и длина переднеспинки) клинальная изменчивость согласована с изменением климатических параметров (среднемесячная температура в июле и число дней с температурой воздуха больше 20°C). Положительная корреляция температуры и длины тела была также выявлена у южноамериканских саранчовых Dichroplus vittatus Bruner 1900 и D. pratensis Bruner 1900 (Bidau, Marti, 2007, 2008). Аналогичную зависимость описывали и другие авторы (Крицкая, 1972; Laiolo et al., 2013).
Роль увлажнения (количества осадков) неоднозначна. Положительная связь обнаружена только с индексом стадности, а ряд признаков (длина и ширина бедра, длина переднеспинки) демонстрирует отрицательную корреляцию с обилием осадков в теплый период. Ряд авторов (Harris et al., 2012; Bai et al., 2016) также указывает на отрицательное влияние осадков на размерные признаки (длину тела и надкрылий): чем больше количество осадков в том или ином регионе, тем меньше морфологические показатели в обитающей там популяции.
Особи, из обнаруженной за пределами известного ареала изолированной популяции в Восточном Забайкалье, хотя и соответствуют диагнозу номинативного подвида, по ряду морфометрических признаков (площадь надкрылья, длина и ширина надкрылья и бедра) значительно отличаются от C. italicus из других регионов. Однако об их принадлежности к этому виду свидетельствуют следующие признаки. Задние крылья у основания окрашены, надкрылья с закругленной вершиной, слабо сужены, заходят за вершину задних бедер. Церки у самца с резким заостренным нижним зубцом нижней вершинной лопасти (рис. 1). Согласно имеющимся определительным ключам и описаниям (Мищенко, 1952 и др.), по окраске и форме крыльев, по строению церок изученные особи из Забайкальского края относятся к номинативному подвиду. Что касается размеров тела, то морфометрические отличия особей из забайкальской популяции можно объяснить экстремальными условиями обитания, такими, как сильная инсоляция и высокое количество осадков. Сходная закономерность была описана у Ch. macrocerus (Крицкая, 1972) и Phaulacridium vittatum Brunner (Parsons, Joern, 2014). Не исключено, что выявленные отличия C. italicus italicus из Забайкальской популяции обусловлены географической изоляцией и представляют собой проявление первой стадии формообразовательного процесса (см. Правдин, 1964).
Список литературы
Арнольди К.В., 1939. К вопросу о непрерывной географической изменчивости в ее общем и таксономическом значении // Зоологический журнал. Т. 18. № 4. С. 685–709.
Атлас СССР, 1983. Ред.: Точенов В.В., Марков В.Ф. М.: Главное управление геодезии и картографии при Совете министров СССР. 194 с.
Ванькова И.А., 2002. Географическая изменчивость фенотипических признаков саранчовых рода Calliptamus Serv. (Orthoptera. Acrididae) // 12-й съезд Русского энтомологического общества. Тезисы докладов. СПБ. С. 53–54.
Ванькова И.А., 2006. Особенности фенотипической изменчивости итальянского пруса Calliptamus italicus L. (Orthoptera. Acrididae) в период вспышки в Кулундинской степи в 1999–2002 гг. // Энтомологические исследования в Северной Азии. Материалы 7-го Межрегионального совещания энтомологов Сибири и Дальнего Востока. Новосибирск. С. 321–322.
Крицкая И.Г., 1972. Географическая изменчивость и внутривидовая дифференциация усатого конька Chorthippus macrocerus (F.-W.) (Orthoptera, Acrididae) // Энтомологическое обозрение. Т. 51. № 2. С. 254–266.
Лачининский А.В., Сергеев М.Г., Чильдебаев М.К., Черняховский М.Е., Локвуд Дж.А., Камбулин В.Е., Гаппаров Ф.А., 2002. Саранчовые Казахстана, Средней Азии и сопредельных территорий. Ларами: Международная Ассоциация Прикладной Акридологии и Университет Вайоминга. 387 с.
Литвинова Н.Ф., 1972. Закономерности географической изменчивости морфологических признаков в роде Euchorthippus (Orthoptera, Acrididae) // Зоологический журнал. Т. 51. № 6. С. 821–828.
Мищенко Л.Л., 1952. Насекомые прямокрылые. Фауна СССР. Т. 4. Вып. 2. М.–Л.: изд. АН СССР. 610 с.
Олигер И.М., 1971. О географической и популяционной изменчивости у некоторых видов саранчовых рода Chorthippus (Orthoptera, Acrididae) // Зоологический журнал. Т. 50. № 11. С. 1658–1664.
Правдин Ф.Н., 1964. Эндемизм и формообразовательный процесс у прямокрылых насекомых (Orthoptera) в горах Средней Азии // Зоологический журнал. Т. 43. № 12. С. 1784–1794.
Сергеев М.Г., Чильдебаев М.К., Ванькова И.А., Гапипаров Ф.А., Камбулин В.Е., Коканова Э., Лачининский А.В., Пшеницына Л.Б., Темрешев И.И., Черняховский М.Е., Соболев Н.Н., Молодцов В.В., 2016. Итальянская саранча [Calliptamus italicus (Linnaeus. 1758)]: морфология, распространение, экология, управление популяциями. Рим: Продовольственная и сельскохозяйственная организация ООН. 330 с.
Bai Y., Dong J.J., Guan D.L., Xie J.Y., Xu J.Y., 2016. Geographic variation in wing size and shape of the grasshopper Trilophidia annulata (Orthoptera: Oedipodidae): morphological trait variations follow an ecogeographical rule // Scientific reports. V. 6. № 1. P. 1–15.
Bidau C.J., Marti D.A., 2007. Dichroplus vittatus (Orthoptera: Acrididae) follows the converse to Bergmann’s rule although male morphological variability increases with latitude // Bulletin of Entomological Research. V. 97. P. 69–79.
Bidau C.J., Marti D.A., 2008. Geographic and climatic factors related to a body-size cline in Dichroplus pratensis Bruner, 1900 (Acrididae, Melanoplinae) // Journal of Orthoptera Research. V. 17. № 2. P. 149–156.
Blanckenhorn W.U., Demont M., 2004. Bergmann and converse Bergmann latitudinal clines in arthropods: two ends of a continuum? // Integrative and Comparative Biology. V. 44. № 6. P. 413–424.
Brown J.H., Gillooly J.F., Allen A.P., Savage V.M., West G.B., 2004. Toward a metabolic theory of ecology // Ecology. V. 85. № 7. P. 1771–1789.
Ciplak B., Sirin D., Taylan M.S., Kaya S., 2008. Altitudinal size clines, species richness and population density: case studies in Orthoptera // Journal of Orthoptera Research. V. 17. № 2. P. 157–163.
Harris R., McQuillan P., Hughes L., 2012. Patterns in body size and melanism along a latitudinal cline in the wingless grasshopper, Phaulacridium vittatum // Journal of Biogeography. V. 39. № 8. P. 1450–1461.
Laiolo P., Illera J.C., Obeso J.R., 2013. Local climate determines intra-and interspecific variation in sexual size dimorphism in mountain grasshopper communities // Journal of Evolutionary Biology. V. 26. № 10. P. 2171–2183.
Lehmann G.U.C., Lehmann A.W., 2008. Variation in body size among populations of the bushcricket Poecilimon thessalicus (Orthoptera: Phaneropteridae): an ecological adaptation // Journal of Orthoptera Research. V. 17. № 2. P. 165–169.
Parsons S.M.A., Joern A., 2014. Life history traits associated with body size covary along a latitudinal gradient in a generalist grasshopper // Oecologia. V. 174. № 2. P. 379–391.
Uvarov B.P., 1966. Grasshoppers and Locusts: Anatomy, physiology, development, phase polymorphism, introduction to taxonomy. Cambridge University Press. Published for the Anti-Locust Research Centre. V. 1. 481 p.
Whitman D.W., 2008. The significance of body size in the Orthoptera: a review // Journal of Orthoptera research. V. 17. № 2. P. 117–134.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал