

Биология внутренних вод, 2021, № 2, стр. 133-141
Морфофункциональный статус монгольского хариуса Thymallus brevirostris в горном озере (юг Восточной Сибири)
И. С. Некрасов a, *, Л. А. Шуман a, А. Г. Селюков a
a Тюменский государственный университет
Тюмень, Россия
* E-mail: i.s.nekrasov@utmn.ru
Поступила в редакцию 13.12.2019
После доработки 10.06.2020
Принята к публикации 29.10.2020
Аннотация
Проведен гистологический анализ внутренних органов озерной (бентосоядной) формы монгольского хариуса Thymallus brevirostris (Kessler, 1879) в горном холодноводном оз. Хиндиктиг-Холь (Тува) в летний период. Установлено, что самки созревают в возрасте 4+, а пик нерестовой активности наблюдается в 5+. Самцы созревают на год раньше самок. В яичниках половозрелых рыб в летний период старшая генерация половых клеток представлена ооцитами фазы вакуолизации цитоплазмы, очередные генерации – многочисленными превителлогенными ооцитами. В семенниках проходит волна сперматогенеза. В эпителии жабр выявлены значительные патоморфологические изменения, что угнетает респираторную функцию. Зарегистрированные слабая гиперемия печени и умеренная базофилия гепатоцитов свидетельствуют о ее нормальном функционировании у хариуса в этот период.
ВВЕДЕНИЕ
Исследования сем. хариусовых Thymallidae проводятся уже более двух столетий (Kessler, 1879; Boulenger, 1898; Cветовидов, 1936; Берг, 1948; Тугарина, Дашидоржи, 1972; Pivnicka, Hensel, 1978; Cеверин, Зиновьев, 1982; Михеев, Мазникова, 2016). В ряде водоемов ареала хариусовые имеют важное значение в структуре рыбного населения литорали, являясь доминирующими видами среди остальных компонентов ихтиоценоза. Происхождение этой группы лососевидных рыб приурочено к водоемам Алтая и Монголии, где отмечено их наибольшее видовое разнообразие (Pallas, 1776; Cветовидов, 1936; Берг, 1948; Бaaсанжав и др., 1988; Travers, 1989; Коskinen et al., 2002; Weiss, 2007; Книжин и др., 2008). Однако, если европейский и сибирский хариусы широко распространены, и их биология достаточно хорошо исследована, то наименее изученным представителем семейства является монгольский хариус Thymallus brevirostris (Kessler, 1879).
Монгольский хариус распространен во внутренних водоемах Северо-Западной Монголии, в бассейнах рек Кобдо и Дзабхан и в ряде озер (Гундризер, 1966; Тугаринa, Дашидоржи, 1972), некоторые из которых находятся в горных регионах и характеризуются экстремальными гидрологическими условиями. В настоящее время считается (Книжин и др., 2008; Cлынько и др., 2010), что монгольский хариус бассейна р. Кобдо представлен крупной (хищной) и мелкой (бентосоядной) формами. Генетическое единство крупной и мелкой форм, однородность по меристическим признакам, сходство в характере варьирования окраски чешуйного покрова и рисунка на спинном плавнике свидетельствуют об их принадлежности к одному виду. Водоемы Центрально-Азиатского бассейна также населяют популяции монгольского хариуса со смешанным типом питания и признаками внешнего строения, в разной степени свойственными обеим формам.
Цель работы – исследовать размерно-возрастной состав и гистофизиологическое состояние гонад, жаберного аппарата и печени монгольского хариуса в оз. Хиндиктиг-Холь (Тува) для оценки состояния этого вида при низких среднегодовых температурах воды в период их максимальных значений.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Озеро Хиндиктиг-Холь моренно-подпрудного происхождения, расположено в пределах высокогорной тундры на высоте 2 306 м над уровнем моря. Площадь озера 6 700 га, питание смешанное, в основном ледниковое. В западной части из него вытекает р. Моген-Бурен, приток р. Кобдо (бассейн Больших Бессточных озер Монголии). Преобладающие глубины в северной части озера 20–25 м, в восточной – >35 м. Сход льда наблюдается в конце июня. По температурному режиму считается самым холодноводным среди основных промысловых водоемов Тувы. Температура воды в конце июня у поверхности – до 5.7°С, в конце июля – 11–12°С. Вода слабоминерализованная (до 30 мг/л), относится к гидрокарбонатному классу, группе натрия. Прозрачность воды по стандартному диску Секки 16 м. По биомассе (0.2 г/м3) и доминирующему комплексу зоопланктона (Calanoida) озеро характеризуется как олиготрофный водоем. В среднем по водоему численность зоопланктона 8.0 тыс. экз./м3, биомасса – 0.2 г/м3. Преобладание мелких форм зоопланктона связано с тем, что младшие возрастные группы хариуса питаются преимущественно зоопланктоном. Биомасса зообентоса в литорали в среднем 2.9 г/м2, в профундали – 1.9 г/м2. По биомассе в литорали доминируют моллюски, хирономиды и гаммарусы, в профундали – хирономиды. Из рыб в оз. Хиндиктиг-Холь обитает только монгольский хариус.
Монгольского хариуса отлавливали c 12 по 20 июля 2010 г. В ходе исследования рыб измеряли и взвешивали, их гонады, жабры и печень фиксировали в смеси Буэна. Гистологический анализ проводили по стандартным методикам (Микодина и др., 2009). Серийные парафиновые срезы толщиной 5 мкм готовили на автоматизированном ротационном микротоме HM 335S (“MICROM”), препараты окрашивали железным гематоксилином по Гейденгайну, заключали в среду Bio Mount (“Bio Optica”) и анализировали на микроскопе AxioImager A1 (“Zeiss”).
С использованием программного обеспечения AxioVision 4.7.1 (“Zeiss”) на срезах яичников измеряли диаметр ооцитов и их ядер, подсчитывали число ооцитов разных фаз развития. На препаратах семенников у каждой особи произвольно выбирали по 3 среза, на которых выделяли по семь областей площадью 80 мкм2 каждая, на этих участках подсчитывали число сперматогониев А- и Б-типов, сперматоцитов I и II порядков.
В жабрах, в пределах исследованной зоны, измеряли ширину респираторных ламелл (проксимальный, медиальный, дистальный участки), число слоев клеток вставочного эпителия, количество слизистых клеток в 1 мм2. Учитывали патоморфологические изменения (цитолиз, утолщение, десквамация, слияние и деструкция респираторных ламелл и др.) и измеряли их площадь. Вычисляли индекс патологии органа, который рассчитывали из суммы долей каждой патологии, помноженной на так называемый коэффициент значимости (Bernet et al., 1999). Коэффициент значимости присваивали каждой патологии в зависимости от ее опасности для здоровья рыбы и респираторной функции жабр (Шуман, 2015).
При оценке состояния печени на гистологических срезах органа учитывали наличие ряда отклонений адаптационного характера (гиперемия, васкуляризация, базофилия цитоплазмы гепатоцитов) и патологических изменений – жировая дистрофия, разрушение печеночной ткани и появление каверн. У каждой особи анализировали по пять срезов печени, на которых в произвольном порядке измеряли площади гепатоцитов, их ядер и липидных включений. Препараты фотографировали камерой AxioCam MRc5 (“Zeiss”).
Всего исследовано 52 экз. монгольского хариуса, 50 из них изучено с применением гистологических методов, приготовлено 50 препаратов гонад, 47 – жабр и 44 – печени. Все расчеты проводили с помощью программ MS Excel 2007.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В уловах преобладали особи в возрасте 4+…5+, по размерным показателям соответствующие литературным данным (Книжин и др., 2008), что свидетельствует о нормальном развитии этого вида в холодноводном оз. Хиндиктиг-Холь (табл. 1). Тем не менее, вследствие устойчиво низких температур воды в течение года, метаболизм у рыб должен быть понижен и сопровождаться замедлением полового созревания, низкой плодовитостью и неежегодным нерестом.
Таблица 1.
Размерно-массовые показатели монгольского хариуса в оз. Хиндиктиг-Холь
| Возраст, лет | Длина по Смитту, мм | Масса, г | ||
|---|---|---|---|---|
| самки | самцы | самки | самцы | |
| 2+…3+ | $\frac{{218.0 \pm 3.4}}{{208 - 230}}$ (n = 7) |
$\frac{{218.4 \pm 9.9}}{{190 - 247}}$ (n = 5) |
$\frac{{86.0 \pm 5.0}}{{70 - 110}}$ (n = 7) |
$\frac{{88.4 \pm 11.0}}{{54 - 106}}$ (n = 5) |
| 4+ | $\frac{{250.4 \pm 6.5}}{{215 - 270}}$ (n = 9) |
$\frac{{248.8 \pm 4.4}}{{235 - 260}}$ (n = 5) |
$\frac{{126.2 \pm 8.6}}{{82 - 160}}$ (n = 9) |
$\frac{{128.8 \pm 5.8}}{{110 - 142}}$ (n = 5) |
| 5+ | $\frac{{273.6 \pm 3.3}}{{256 - 300}}$ (n = 11) |
$\frac{{274.1 \pm 4.9}}{{255 - 292}}$ (n = 9) |
$\frac{{159.5 \pm 5.8}}{{124 - 196}}$ (n = 11) |
$\frac{{156.7 \pm 7.8}}{{122 - 192}}$ (n = 9) |
| 6+ | $\frac{{285.0 \pm 10.2}}{{265 - 323}}$ (n = 5) |
286 (n = 1) |
$\frac{{190.8 \pm 20.5}}{{142 - 266}}$ (n = 5) |
202 (n = 1) |
Примечание. Здесь и в табл. 2–4 над чертой – среднее значение и его ошибка, под чертой – min–max, n – число исследованных рыб.
Гонады. Яичники у большинства самок монгольского хариуса в оз. Хиндиктиг-Холь находились на III стадии зрелости, в которой старшей генерацией половых клеток были вителлогенные ооциты фазы вакуолизации цитоплазмы (рис. 1а). Очередное поколение половых клеток представлено превителлогенными ооцитами (рис. 2). Они же в большом количестве (до 77%) присутствовали у отнерестившихся особей (рис. 1б). У части рыб в возрастных группах 4+…7+, по сравнению с трех- и четырехлетними особями, число вителлогенных ооцитов фазы вакуолизации цитоплазмы значительно возрастало (рис. 1в). Существенное различие соотношения ооцитов разных фаз в старших возрастных группах может свидетельствовать, что в оз. Хиндиктиг-Холь хариус созревает в возрасте 4+ и старше. У самок в возрасте 5+…7+ некоторые вителлогенные ооциты начинали резорбироваться, тогда как другие вступали в период накопления желтка при одновременно проходящей дегенерации постовуляторных фолликулов (рис. 1г). Следовательно, такие самки отнерестились в текущем году и готовятся к нересту в следующем.
Рис. 1.
Яичники монгольского хариуса на разных стадиях зрелости: а – в ооцитах старшей генерации начинается фаза вакуолизации цитоплазмы (×100), б – среди ооцитов фазы вакуолизации цитоплазмы видны дегенерирующие постовуляторные фолликулы (×40), в – участок яичника IIIб стадии зрелости с большим количеством ооцитов фазы вакуолизации цитоплазмы (×40), г – начало накопления желтка в ооците, можно видеть дегенерацию постовуляторного фолликула (×400).

Рис. 2.
Соотношение (%) фаз ооцитов у разных возрастных групп монгольского хариуса: а – превителлогенные ооциты, б, в – вителлогенные ооциты фазы вакуолизации цитоплазмы и фазы накопления желтка соответственно.

Анализ половых клеток на разных стадиях оогенеза показал (табл. 2), что у разновозрастных самок размеры ооцитов сходных фаз достоверно не различались (p > 0.05) или варьировали незначительно.
Таблица 2.
Генеративные показатели у разновозрастных групп монгольского хариуса
| Половые клетки | Диаметр ооцитов, мкм | Диаметр ядер, мкм | Ядерно-цитоплазматическое соотношение, % |
|---|---|---|---|
| 2+…3+, лет | |||
| Превителлогенные ооциты | $\frac{{258.2 \pm 7.6}}{{240--273}}$ | $\frac{{105.3 \pm 1.7}}{{101--109}}$ | 68.93 |
| Вителлогенные ооциты | |||
| фаза вакуолизации цитоплазмы | $\frac{{494.9 \pm 10.3}}{{485--505}}$ | $\frac{{179.7 \pm 13.5}}{{166--193}}$ | 57.03 |
| 6+…7+, лет | |||
| Превителлогенные ооциты | $\frac{{271.4 \pm 2.1}}{{266--275}}$ | $\frac{{107.3 \pm 2.5}}{{103--112}}$ | 65.40 |
| Вителлогенные ооциты | |||
| фаза вакуолизации цитоплазмы | $\frac{{454.2 \pm 31.6}}{{423--486}}$ | $\frac{{151.0 \pm 6.8}}{{144--158}}$ | 49.80 |
| фаза накопления желтка | $\frac{{638.8 \pm 47.1}}{{533--745}}$ | $\frac{{200.4 \pm 9.4}}{{184--226}}$ | 45.71 |
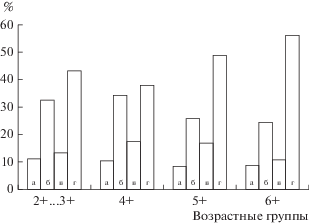
Семенники у основной массы самцов находились на II стадии зрелости, половые клетки были представлены сперматогониями (рис. 3а). У части особей появлялись цисты со сперматоцитами (рис. 3в). В гонадах большинства самцов присутствовали все типы клеток, но количество сперматоцитов II порядка с возрастом увеличивалось. У рыб в возрасте 2+…4+ в семенниках преобладали сперматоциты II порядка и сперматогонии Б-типа. (рис. 3б). В меньшем количестве присутствовали сперматоциты I порядка и сперматогонии А‑типа. В возрасте 5+ и старше наибольшую долю половых клеток составляли сперматоциты II порядка (рис. 4), в семенных канальцах присутствовали группы остаточных спермиев от прошедшего нереста (рис. 3г).
Рис. 3.
Состояние семенников монгольского хариуса: а – II стадия зрелости, половые клетки представлены сперматогониями (×1000), б – массы сперматоцитов I и II порядков в семеннике III стадии зрелости (×400), в – преобладание сперматогониев Б-типа в семеннике II стадии зрелости (×1000), г – сперматоциты в окружении цист сперматогониев Б-типа, в просвете семенного канальца – резорбирующиеся остаточные спермии (×1000).

Рис. 4.
Соотношение (%) половых клеток в семенниках монгольского хариуса разных возрастных групп: а – сперматогонии А-типа, б – сперматогонии B-типа, в – сперматоциты I порядка, г – сперматоциты II порядка.
Жабры. Воды оз. Хиндиктиг-Холь, находящегося в природной зоне, исключающей химическое загрязнение, могут считаться вполне приемлемыми для монгольского хариуса, поэтому мы не предполагали выявление каких-либо серьезных отклонений в состоянии его жаберного аппарата. Для части особей данное предположение оказалось справедливым (рис. 5а), но у большинства изученных рыб нами установлены различные по характеру и глубине патологии. Аномалии в строении жаберного аппарата монгольского хариуса проявлялись в плазмолизе и частичном (рис. 5б) или полном слиянии респираторных ламелл, охватывающем отдельные участки жабр (рис. 5в) с последующей деструкцией респираторного и вставочного эпителиев (рис. 5г). Разнообразие и частота встречаемости нарушений увеличивались с возрастом, что нашло свое отражение в индексе патологии (табл. 3). При этом число слоев клеток вставочного эпителия у половозрелых особей несколько снижалось. Наблюдаемый высокий индекс патологий в жабрах рыб старшевозрастной группы при достоверно меньшем количестве слизистых клеток и сходном числе клеток вставочного эпителия расходятся с проведенными на форели исследованиями В.Е. Матей (1987). В соответствии с ними, барьер, создаваемый жаберным эпителием между внешней средой и кровью, возрастает при стрессе или в неблагоприятных условиях (закисление, различные виды загрязнений).
Рис. 5.
Жаберный аппарат монгольского хариуса из оз. Хиндиктиг-Холь: а – нормальное состояние жаберного эпителия (×200), б – утолщение (стрелка) и плазмолиз респираторных ламелл (×200), в – слияние респираторных ламелл (×100), г – слияние филаментов и разрушение респираторных ламелл (×40).
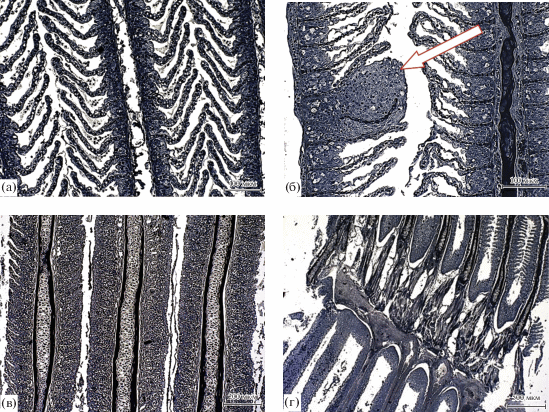
Таблица 3.
Характеристики жаберного эпителия у монгольского хариуса в оз. Хиндиктиг-Холь (июль 2010 г.)
| Показатель | Возраст | ||
|---|---|---|---|
| 3+ (n = 4) |
4+ (n = 5) |
5+ (n = 9) |
|
| Число слоев клеток вставочного эпителия | $\frac{{4.0 \pm 0.4}}{{3.3 - 5.0}}$ | $\frac{{3.5 \pm 0.6}}{{2.5 - 5.6}}$ | $\frac{{3.5 \pm 0.3}}{{2.7 - 5.3}}$ |
| Ширина респираторных ламелл, мкм | $\frac{{17.6 \pm 0.7}}{{16.2 - 19.2}}$ | $\frac{{19.6 \pm 2.4}}{{16.5 - 29.1}}$ | $\frac{{19.6 \pm 1.5}}{{14.5 - 30.1}}$ |
| Количество слизистых клеток в 1 мм2 | $\frac{{168.6 \pm 51.8}}{{104.4 - 322.0}}$ | $\frac{{174.4 \pm 25.1}}{{132.6 - 219.5}}$ | $\frac{{122.4 \pm 15.3}}{{65.5 - 199.8}}$ |
| Индекс патологии, % | $\frac{{1.0 \pm 0.4}}{{0.2 - 1.9}}$ | $\frac{{1.2 \pm 0.4}}{{0.6 - 1.9}}$ | $\frac{{1.3 \pm 0.2}}{{0.2 - 2.3}}$ |
Можно утверждать, что значимые отклонения в состоянии жаберного аппарата у монгольского хариуса в оз. Хиндиктиг-Холь вызваны влиянием некоего неблагоприятного фактора(ов).
Печень. При изучении печени установлено, что у самок и самцов хариуса этот орган гиперемирован, но каких-либо существенных отклонений в его состоянии ни на макро-, ни на микроскопическом уровне не отмечено. Размеры гепатоцитов у рыб разных возраста и пола (табл. 4) почти не различались (p > 0.05). Цитоплазма гепатоцитов была умеренно базофильна. Сами клетки печени имели немногочисленные липидные капли, количество которых варьировало незначительно (рис. 6а). Наибольшее количество и вариабельность липидных капель были свойственны гепатоцитам неполовозрелых особей обоего пола в возрасте 2+…3+ (рис. 6б). Повышенная степень базофилии клеток печени отмечена у созревающих особей с гонадами III стадии зрелости: вителлогенными ооцитами у самок и сперматоцитами II порядка у самцов.
Таблица 4.
Размерные плоказатели гепатоцитов монгольского хариуса в оз. Хиндиктиг-Холь
| Показатель | Возраст, лет | ||
|---|---|---|---|
| 3+ (n = 5) |
4+ (n = 4) |
5+ (n = 7) |
|
| Диаметр гепатоцитов, мкм | $\frac{{6.0 \pm 0.18}}{{5.4 - 6.4}}$ | $\frac{{5.7 \pm 0.07}}{{5.5 - 5.8}}$ | $\frac{{5.8 \pm 0.07}}{{5.5 - 5.9}}$ |
| Диаметр ядер гепатоцитов, мкм | $\frac{{2.1 \pm 0.06}}{{1.9 - 2.3}}$ | $\frac{{2.0 \pm 0.07}}{{1.8 - 2.1}}$ | $\frac{{2.0 \pm 0.06}}{{1.9 - 2.3}}$ |
| Ядерно-цитоплазматическое соотношение, % | 54.3 | 53.9 | 52.6 |
Рис. 6.
Печень монгольского хариуса разных возрастных групп: а – в цитоплазме гепатоцитов половозрелых рыб липидные капли присутствуют в незначительном количестве (×1000), б – многочисленные липидные капли в гепатоцитах неполовозрелых особей (×1000). Стрелками указаны липидные капли.

Таким образом, у подавляющего большинства особей монгольского хариуса в горном холодноводном оз. Хиндиктиг-Холь отмечен типичный для этого вида темп роста с некоторой задержкой полового созревания, возможно, вызванной снижением морфофункционального состояния жаберного аппарата; у большинства самок при достижении половой зрелости – ежегодный нерест.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Согласно ранее проведенным исследованиям (Книжин и др., 2008; Cлынько и др., 2010), размерные показатели “бентосоядной” формы монгольского хариуса из оз. Хиндиктиг-Холь вполне сопоставимы с показателями этого вида из других водоемов (Тугарина, Дашидоржи, 1972; Шатуновский, 1983).
Нами показано, что у большинства самок старшая генерация половых клеток в период активного посленерестового нагула находилась в фазе вакуолизации цитоплазмы. Очередной пул клеток у рыб всех возрастных групп был представлен превителлогенными ооцитами, и они же в своем большинстве присутствовали у отнерестившихся особей. У части самок в возрастных группах 4+…7+, по сравнению с трех- и четырехлетними особями, число вителлогенных ооцитов фазы вакуолизации цитоплазмы значительно возрастало. Существенное различие соотношения ооцитов разных фаз в разных возрастных группах может свидетельствовать о том, что в этом озере хариус созревает в возрасте 4+ и старше. У самок в возрасте 5+…7+ некоторые вителлогенные ооциты начинали резорбироваться, но основная масса половых клеток приступала к накоплению желтка. В размерных характеристиках ооцитов монгольского хариуса из оз. Хиндиктиг-Холь явных различий с ооцитами байкальского хариуса (Зайцева и др., 2010) не выявлено. Отсутствие пропускающих предстоящий нерестовой сезон самок и наличие в яичниках хариуса следов прошедшего нереста в виде опустевших фолликулов свидетельствуют об экологической пластичности его репродуктивной системы, что можно соотнести с состоянием репродуктивной системы сиговых рыб Обь-Иртышского бассейна (Селюков, 2002а, 2002б, 2012; Cелюков и др., 2012). Таким образом, самки “бентосоядной” формы монгольского хариуса в олиготрофном оз. Хиндиктиг-Холь созревают в возрасте 4+, а пик нерестовой активности наблюдается в возрасте 5+.
У большей части самцов половые клетки представлены сперматогониями. В семенниках остальных рыб присутствовали все типы клеток. В возрастной группе 2+…3+ наблюдалось преобладание сперматоцитов II-го порядка и сперматогониев B-типа, что свидетельствовало о высокой вероятности участия этих особей в следующем нерестовом сезоне, т.е. самцы созревали на год раньше самок. В семенниках некоторых мужских особей старших возрастных групп, находящихся в посленерестовом состоянии и готовящихся к очередному нересту, в просветах семенных канальцев наблюдались остаточные спермии. В целом, в развитии гонад какие-либо существенные отклонения отсутствовали.
Анализ состояния жаберного аппарата рыб часто используют для оценки их морфофункционального статуса (Матей, 1990), у монгольского хариуса в оз. Хиндиктиг-Холь оно находится в крайне угнетенном состоянии. Причины подобных аномалий, очевидно, вызваны гидрохимическим режимом, обусловленным аэрогенными переносами и/или рудогенными факторами, слагающими горный массив.
Нарушений в состоянии печени исследуемого вида, в отличии от сиговых рыб Северной Сосьвы (Некрасов и др., 2014), не было установлено, в гепатоцитах неполовозрелых особей обоего пола (2+…3+) отмечен значительный объем липидных включений в сравнении с печеночными клетками рыб старших возрастных групп. В печени созревающих самок липидные включения исчезают, что свидетельствует об энергетических затратах организма на синтез вителлогенина.
Состояние внутренних органов у монгольского хариуса данной популяции несет отчетливые признаки токсического стресса, касающиеся преимущественно респираторной системы. В отношении печени и половых желез аналогичные утверждения пока преждевременны, однако нарушение ключевой системы жизнедеятельности неизбежно ведет к угнетению остальных функций организма.
Выводы. Размерно-массовые показатели у “бентосоядной” формы монгольского хариуса из оз. Хиндиктиг-Холь соответствуют приведенным в литературе из других водоемов, что свидетельствует о достаточной кормовой базе, обеспечивающей рост хариуса даже в условиях низких среднегодовых температур, в уловах преобладали пяти–шестилетние особи обоего пола. Самки хариуса созревают на пятом году жизни, самцы – на год раньше. В яичниках половозрелых особей присутствовали дегенерирующие опустевшие фолликулы от прошлогоднего нереста, отмечен ежегодный нерест самок и самцов. Существенные отклонения в жаберном эпителии как важнейшем биомаркере качества окружающей среды свидетельствуют о наличии некоего фактора (ов), угнетающего респираторную функцию этого вида в оз. Хиндиктиг-Холь. Печень у всех исследованных рыб не имела аномалий, выявленные различия в размерах клеток и наличии липидных включений отражают полоспецифические особенности физиологического состояния разновозрастных особей.
Список литературы
Баасанжав Г., Дгебуадзе Ю.Ю., Лапин В.И. 1988. К изучению хариусов водоемов реки Кобдо // Природные условия, растительный покров и животный мир Монголии. Пущино: Научный центр биологических исследований РАН. С. 319.
Берг Л.С. 1948. Рыбы пресных вод СССР и сопредельных стран. Москва: Изд-во АН СССР. Ч. 1.
Гундризер А.Н. 1966. О нахождении монгольского хариуса Thymallus brevirostris Kessler в водоемах СССР // Вопр. ихтиологии. Т. 6. Вып. 4. С. 638.
Зайцева А.Н., Смирнова-Залуми Н.С., Захарова Н.И. 2010. Сравнительный анализ роста ооцитов у двух форм байкальского хариуса Thymallus baicalensis (Thymallidae) // Вопр. ихтиол. Т. 50. № 4. С. 541.
Книжин И.Б., Вайс С.Д., Богданов Б.Э. и др. 2008. Хариусы (Thymallidae) водоемов западной Монголии: морфологическое и генетическое разнообразие // Вопр. ихтиол. Т. 48. № 6. С. 745.
Матей В.Е. 1987. Ультраструктура жаберного эпителия ручьевой форели в норме и при закислении воды // Цитология. Т. 29. № 10. С. 1120.
Матей В.Е. 1990. Функциональная морфология жаберного эпителия пресноводных костистых рыб // Физиология, биохимия и токсикология пресноводных животных. Ленинград: Наука. С. 104.
Микодина Е.В., Седова М.А., Чмилевский Д.А. и др. 2009. Гистология для ихтиологов: Опыт и советы. Москва: Всерос. научно-исследовательский ин-т рыб. хоз-ва и океанографии.
Михеев П.Б., Мазникова О.А. 2016. Сравнительный анализ двух видов амурских хариусов Thymallus (Salmoniformes; Thymallidae) в зоне их симпатрии по ряду остеологических и морфологических признаков // Биология внутренних вод. № 1, С. 62.https://doi.org/10.7868/S0320965216010137
Некрасов И.С., Пашина Л.С., Селюков А.Г. 2014. Морфофункциональные изменения печени сиговых рыб в условиях реки Северная Сосьва в период летнего нагула // Вестник Тюменского государственного университета. Экология и природопользование. № 12. С. 114.
Световидов А.Н. 1936. Европейско-азиатские хариусы (genus Thymallus Cuvier) // Тр. Зоол. ин-та АН СССР. Т. 3. С. 183.
Северин С.О., Зиновьев Е.А. 1982. Кариотипы изолированных популяций Thymallus arcticus (Pallas) бассейна реки Оби // Вопр. ихтиол. Т. 22. Вып. 1. С. 27.
Селюков А.Г. 2002а. Репродуктивная система сиговых рыб (Coregonidae, Salmoniformes) как индикатор состояния экосистемы Оби. I. Половые циклы пеляди Coregonus peled // Вопр. ихтиол. Т. 42. № 1. С. 85.
Селюков А.Г. 2002б. Репродуктивная система сиговых рыб (Coregonidae, Salmoniformes) как индикатор состояния экосистемы Оби. II. Половые циклы муксуна Coregonus muksun // Вопр. ихтиол. Т. 42. № 2. С. 225.
Селюков А.Г. 2012. Морфофункциональные изменения рыб бассейна Средней и Нижней Оби в условиях возрастающего антропогенного влияния // Вопр. ихтиол. Т. 52. № 5. С. 581.
Селюков А.Г., Шуман Л.А., Некрасов И.С. 2012. Состояние гонад у лососевидных рыб в субарктических озерах Ямала и Гыдана // Вестник Тюменского государственного университета. Экология и природопользование. № 6. С. 31.
Слынько Ю.В., Мендсайхан Б., Касьянов А.Н. 2010. К вопросу о внутривидовых формах монгольского хариуса (Thymallus brevirostris Kessl.) озера Хотон Нур (Западная Монголия) // Вопр. ихтиол. Т. 50. № 1. С. 32.
Тугарина П.Я., Дашидоржи А. 1972. Монгольский хариус Thymallus brevirostris Kessler бассейна р. Дзабхан // Вопр. ихтиол. Т. 12. Вып. 5. С. 843.
Шатуновский М.И. 1983. Рыбы Монгольской Народной Республики. Москва: Наука.
Шуман Л.А. 2015. Гистопатологические изменения и репродукционный потенциал у рыб в водоемах Обь-Иртышского бассейна с различной антропогенной нагрузкой: Дис. … канд. биол. наук. Москва: Всерос. научно-исследовательский ин-т рыб. хоз-ва и океанографии.
Bernet D., Schmidt H., Meier W. et al. 1999. Histopathology in fish: Proposal for a protocol to assess aquatic pollution // J. Fish Diseases. № 22. P. 25. https://doi.org/10.1046/j.1365-2761.1999.00134.x
Boulenger G.A. 1898. On a new genus of salmonoid fishes from the Altai Mountains // Ann. Mag. Natur. Hist. Ser. 7. V. I. № 4. P. 329.
Kessler K. 1879. Beitrage zur Ichthyologie von Central-Asien // Melanges diol. Tires Bull. Acad. St.-Petersb. V. 10. S. 233.
Koskinen M.T., Knizhin I., Primmer C.R. et al. 2002. Mitochondrial and nuclear DNA phylogeography of Thymallus spp. (grayling) provides evidence of ice-age mediated environmental perturbations in the world’s oldest body of freshwater, Lake Baikal // Mol. Ecol. V. 11. P. 2599. https://doi.org/10.1046/j.1365-294X.2002.01642.x
Pallas P.S. 1776. Reise durch verschiedene Provincen des Russischen Reiches. Th. 3. St.- Petersburg: Kaiserl. Acad. Der Wiss.
Pivnicka K., Hensel K. 1978. Morphological variation in the genus Thymallus Cuvier, 1829 and recognition of the species and subspecies // Acta Univ. Carolinae-Biologica 1975–1976. V. 4. P. 37.
Travers R.A. 1989. Systematic account of a collection of fishes from the Mongolia People"s Republic: with a review of the hydrobiology of the major Mongolian drainage basins // Bull. Brit. Mus. Natur. Hist. (Zool.). V. 55. № 2. P. 173.
Weiss S., Knizhin I., Romanov V., Kopun T. 2007. Secondary contact between two divergent lineages of grayling Thymallus in the lower Enisey basin and its taxonomic implications // J. Fish Biol. V. 71. Suppl. C. P. 371. https://doi.org/10.1111/j.1095-8649.2007.01662.x
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод