

Биология внутренних вод, 2023, № 1, стр. 28-34
Особенности метаболизма планктона: результаты автоматизированных измерений на Можайском водохранилище
А. В. Гончаров a, *, В. В. Пуклаков a, М. Г. Гречушникова a, Н. М. Юмина a
a Московский государственный университет им. М.В. Ломоносова
Москва, Россия
* E-mail: mama15333@mail.ru
Поступила в редакцию 31.08.2021
После доработки 23.06.2022
Принята к публикации 27.06.2022
- EDN: KSGSFO
- DOI: 10.31857/S0320965223010060
Аннотация
Метаболизм планктона определяет кислородный режим водоема, его экологическое состояние, обмен СО2 c атмосферой. Нами использовано автоматизированное устройство, которое позволило провести массовые измерения внутрисуточных значений валовой первичной продукции (ВП) и дыхания (Д) планктона. В работе на Можайском водохранилище летом 2017 г. выявлен важный факт: максимумы ВП существенно опережают таковые Д в дневное время. Это объясняется тем, что ВП связана с солнечной радиацией (максимум в середине дня), а Д – с температурой воды (максимум во второй половине дня). Соответственно, наибольшее содержание растворенного кислорода наблюдается во второй половине дня, когда ВП и Д уравниваются. Рассматриваемый метод может использоваться для непрерывного мониторинга метаболизма планктона и его реакции на различные гидрометеорологические, гидрохимические и антропогенные факторы.
ВВЕДЕНИЕ
Первичная продукция и дыхание, составляющие метаболизм сообщества, определяют особенности функционирования, а также состав и структуру различных экосистем (Odum, 1983; Wetzel, 2001). Существует множество методов изучения метаболизма планктона; каждый из них имеет свои достоинства и недостатки (Винберг, 1960; Wetzel, Likens, 2000; Staehr et al., 2011; Vinçon-Leite, Casenave, 2019). Например, в широко известном скляночном радиоуглеродном методе определения первичной продукции фитопланктона часть вносимого изотопа С-14 может использоваться фитопланктоном повторно, занижая результат (Williams et al., 2002). Исследование метаболизма планктона по суточному ходу кислорода в водоеме осложняется вертикальным перемешиванием водной массы и обменом кислорода с атмосферой (McNair et al., 2015).
Имеющиеся подходы и методы нацелены, как правило, на определение ВП и Д за достаточно длительные промежутки времени – сутки, месяцы, сезоны. При рассмотрении внутрисуточных изменений возникают методические проблемы: обычный кислородный скляночный метод слишком трудоемок для проведения постоянных круглосуточных наблюдений (Depew et al., 2006, Минеева и др., 2016). Исследование суточного хода кислорода, измеряемого непосредственно в водоеме с помощью логгеров О2, дает возможность оценивать короткопериодичные (например, почасовые) изменения чистой первичной продукции днем и дыхания сообщества ночью (Staehr et al., 2010; LaBuhn, Klump, 2016). Однако получить почасовые изменения таких основных параметров, как ВП и Д, при этом не удается (Hanson et al., 2008).
Суточные или месячные изменения метаболизма планктона состоят из короткопериодичных внутрисуточных изменений, которые постоянно происходят в водоемах при изменении гидрометеорологических условий – солнечной радиации, температуры, ветра, атмосферных осадков. Чтобы восполнить имеющийся пробел в методах изучения метаболизма планктона, нами была разработана автоматизированная установка, позволяющая круглосуточно измерять ЧП, Д и определять ВП в водоеме (Гончаров и др., 2018). С помощью такой установки нами проведены исследования на Можайском водохранилище – с целью изучить метаболизм планктона при краткосрочных изменениях гидрометеорологических условий.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Работу проводили в августе 2017 г. на Можайском водохранилище, расположенном в 100 км к северо-западу от г. Москвы и служащим одним из источников водоснабжения столицы. Длина водоема 28 км, площадь 30.7 км2, средняя глубина – 7 м, у плотины – до 22 м, водообмен ~2 раза за год. Водоем хорошо изучен (Goncharov, 2007; Belova, Kremenetskaya, 2010; Пуклаков и др., 2015), характеризуется высокой биологической продуктивностью с концентрацией хлорофилла а 13–67 мкг/л, биомассой фитопланктона в период массового развития водорослей ~5–10 мг/л, ВП ~1 г С/(м2 · сут). Концентрация фосфора фосфатов – 0.01–0.54 Р мг/л. В летний период водохранилище стратифицировано, при этом в гиполимнионе ежегодно наблюдается аноксия, однако в поверхностном слое содержание кислорода может значительно превышать 100% насыщения. Прозрачность воды по диску Секки летом обычно 1–2 м.
Для регистрации внутрисуточных значений ВП и Д нами разработана автоматизированная продукционная установка, описанная ранее (Гончаров и др., 2018). В основе ее работы лежит кислородный скляночный метод, в котором автоматизирована смена воды и измерение растворенного кислорода. Вода в сосудах менялась каждые 3 ч, кислород измеряли каждые 15 мин. Разность последовательно измеренных значений кислорода в темном сосуде представляет собой Д, аналогичная разница в светлом сосуде – ЧП. Сумма Д и ЧП равна ВП. Следует учитывать, что дыхание в темноте и на свету неодинаковы, и это вносит некоторую неопределенность в ВП и Д. При анализе рассматривали часовые и трехчасовые значения показателей, полученные суммированием данных за 15-минутные интервалы.
Кроме того, каждый час регистрировали температуру воды на глубинах 0.1, 0.5, 1, 2, 3, 4, 6, 7, 10, 14 м и ФАР – с помощью приборов фирмы Оnset, США (http://www.onsetcomp.com). Продукциометр и основные приборы были установлены на плавучей платформе в середине Красновидовского плеса Можайского водохранилища (координаты 55°34.958′ c.ш., 035°51.536′ в.д.), сосуды располагались на глубине 0.5 м. Обслуживание продукциометра заключалось в еженедельной промывке сосудов.
При анализе изменений погоды использовали данные метеостанции г. Можайска, расположенной в 12 км от пункта наблюдений; проводили единичные измерения концентрации фосфатов в воде методом Морфи-Райли11. Результаты наблюдений обрабатывали с помощью стандартных статических программ.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Гидрометеорологические условия. По погодным условиям время наблюдений четко делится на два периода: антициклональный (с 12 по 21 августа при средних значениях атмосферного давления Ратм 1000 гПа, температуры воздуха Тв 20.4°С, скорости ветра W 1.5 м/с, общей облачности N 3 балла) и циклональный (с 22 по 31 августа при Ратм 990 гПа, Тв 14.4°С, W 2.4 м/с, N 8 баллов).
При антициклональном типе погоды суточная амплитуда изменений температуры воды в поверхностном слое достигала в среднем 1.4°С, при циклональном – почти не заметна на фоне общего плавного снижения температуры воды поверхностного слоя к концу периода до 18.2°С (рис. 1а).
Рис. 1.
Гидрометеорологические (a) и продукционно-биологические (б) показатели в поверхностном слое Можайского водохранилища в августе 2017 г. Т ‒ температура, ВП – валовая первичная продукция, Д – дыхание планктонного сообщества, ФАР – фотосинтетическая активная солнечная радиация; О2 ‒ концентрация кислорода.

Режим поступления ФАР резко изменялся. В первый период среднесуточное поступление ФАР было достаточно стабильным (в среднем 438 μE/(м2 · с)), максимум ФАР приходился на 12–13 ч и достигал 1541 μE/(м2 · с). Во второй период при прохождении циклона с 22 по 31 августа среднее значения ФАР снизилось до 186 μE/(м2 · с), максимум ФАР в большинстве случаев по-прежнему приходился на 12–13 ч, а его значения не превышали 1000 μE/(м2 · с).
Такие различия погодных условий сказались не только на характеристиках поверхностного слоя воды, но и на вертикальной структуре водных масс, которая имеет существенное значение для протекающих в водоеме процессов продукции и дыхания. Во время антициклональной погоды наблюдали усиление вертикальной стратификации вод (рис. 2). Разность среднесуточных значений поверхностной и придонной температуры воды увеличилась с 7.1°С 12 августа до 7.6°С 21 августа. Сезонный слой скачка температуры располагался на глубине 7 м.
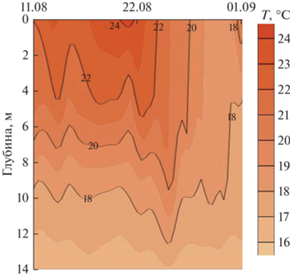
Приход циклона сопровождался усилением облачности до 10 баллов, скорости ветра до 4–5 м/с, выпадением атмосферных осадков. Это привело к быстрому охлаждению верхних слоев воды. К 30 августа в Красновидовском плесе сформировался почти однородный эпилимнион до глубины 10 м: разность поверхностной и придонной температуры воды уменьшилась до 1.8°С.
Результаты измерений 11 и 20 августа показали, что содержание фосфора фосфатов достигало 4–5 мкг Р/л у поверхности и 240–390 мкг Р/л у дна. В конце второго периода (28 августа) концентрация у поверхности увеличилась до 20 мкг Р/л за счет перемешивания водной массы.
Валовая первичная продукция и дыхание планктона. Внутрисуточные изменения ВП и Д в водоеме представлены на рис. 1а. Видно, что в циклональный период продукция снижается в 1.5 раза по сравнению с таковой в антициклональный (табл. 1), дыхание тоже снижается в ~1.6 раза. Вместе с тем, во время похолодания в циклональный период (26, 27, 30 августа) наблюдали довольно высокие значения ВП, как и в начале антициклонального периода (рис. 1а). По-видимому, температура воды, в данном случае, не была решающим фактором продукционного процесса. Об этом свидетельствует и то, что максимумы ВП в антициклональный период не совпадают с таковыми температуры воды (рис. 1а). Они опережают температуру воды на несколько часов.
Таблица 1.
Средние значения показателей за антициклональный (I) и циклональный (II) периоды исследования и соотношение между ними в августе 2017 г. на Можайском водохранилище
| Показатель | I (12.08–21.08) | II (22.08–31.08) | I/II | ||
|---|---|---|---|---|---|
| среднее | Cv, % | среднее | Cv, % | ||
| Т, °С | 23.5 | 2 | 20.6 | 8 | 1.14 |
| ВП, мгО2/(л · ч) | 0.200 | 36 | 0.134 | 26 | 1.49 |
| Д, мгО2/(л · ч) | 0.158 | 19 | 0.099 | 23 | 1.60 |
| ФАР, μE/(м2 · с) | 438 | 11 | 186 | 44 | 2.35 |
| Содержание О2, мг О2/л | 12.01 | 8 | 8.03 | 21 | 1.50 |
| ВП/Д | 1.32 | 15 | 1.46 | 27 | 0.90 |
Примечание. Здесь и в табл. 2, ВП – валовая первичная продукция, Д – дыхание планктонного сообщества, ФАР – фотосинтетическая активная солнечная радиация, T – температура воды, О2 – растворенный кислород, Cv – коэффициент вариации.
ВП тесно связана с ФАР (рис. 1а, табл. 2); коэффициент корреляции r = 0.85 (табл. 2), однако связь ВП с температурой воды очень слабая (r = = 0.22). Максимумы температуры существенно отстают от таковых ФАР (рис. 1а). Причина этого в том, что максимум ФАР наблюдался в 12–13 ч, а температура воды достигала суточного максимума значительно позже, поскольку вода из-за высокой теплоемкости прогревается медленно (кроме того, температура воды зависит от факторов, влияющих на теплообмен с атмосферой и нижележащими слоями воды). Поэтому, когда во второй половине дня (~17 ч) температура воды достигает максимума, солнце уже садится и освещенность становится лимитирующим фактором продукционного процесса.
Таблица 2.
Коэффициенты корреляции между рассматриваемыми показателями по данным за каждые 3 ч в период с 12 по 30 августа 2017 г.
| Показатель | ФАР | T | ВП | Д | ВП/Д | O2 |
|---|---|---|---|---|---|---|
| ФАР | 1.00 | – | – | – | – | – |
| T | 0.23 | 1.00 | – | – | – | – |
| ВП | 0.85 | 0.22 | 1.00 | – | – | – |
| Д | 0.10* | 0.59 | 0.30 | 1.00 | – | – |
| ВП/Д | 0.77 | –0.03* | 0.84 | –0.15* | 1.00 | – |
| O2 | 0.17 | 0.92 | 0.18 | 0.69 | –0.12* | 1.00 |
Выявлена умеренная корреляция (r = 0.59) Д с температурой (табл. 2). Максимумы Д обычно наблюдаются позже таковых ВП и чаще совпадают с температурой (рис. 1а). Это даeт основание утверждать, что не только температура и ФАР разнесены во времени, но и биологические процессы, которые от них зависят. Такое несовпадение во времени процессов выделения и поглощения кислорода может иметь очень существенное значение для кислородного и вообще гидрохимического режима водоема. Дополнительной причиной временного несовпадения ВП и Д в течение суток может быть большая чувствительность гетеротрофных процессов (ответственных за Д) к изменению температуры, чем фотосинтетических (Минеева, 2009). Это обусловлено различием ферментов в клеточных структурах, ответственных за протекание рассматриваемых процессов.
Растворенный кислород (O2) и отношение ВП/Д. Отношение ВП/Д считается фундаментальным экосистемным параметром (Odum, 1983). При ВП/Д = 1 основные процессы в системе сбалансированы; ВП/Д >1 свидетельствует о накоплении органического вещества, ВП/Д < 1 – о его расходовании, сопровождаемом, в частности, потреблением О2 и выделением СО2 в атмосферу (Hanson et al., 2004). Рассмотрением этого параметра в водоемах занимались разные исследователи (Depew et al., 2006; Hoellein et al., 2013). При этом речь шла о данных за сутки, месяцы, сезоны. Возможность проведения массовых внутрисуточных измерений появилась лишь благодаря разработанному нами автоматизированному методу (Гончаров др., 2018).
Внутрисуточные изменения отношения ВП/Д в поверхностном слое водоема представлены рис. 1б. Максимальные значения ВП/Д приходятся на середину дня. Этот показатель коррелирует с ВП и ФАР (табл. 2), поскольку в полдень продукция максимальна, а наибольшее дыхание наблюдается во второй половине дня. Средние суточные значения ВП/Д мало различаются в теплый и прохладный периоды (табл. 1). Это связано с тем, что первичная продукция во второй период активизируется, по нашему мнению, с увеличением содержания фосфатов у поверхности (с 4–5 до 20 мкг Р/л из-за вертикального перемешивания воды), кроме того, при похолодании уменьшается Д.
Несколько неожиданны максимумы кислорода, приуроченные ко второй половине дня (рис. 1б) – предполагалось, они тоже должны быть в полдень. Это особенно заметно в первый период (антициклональный) с очень слабым перемешиванием воды, когда выделяющийся в течение дня кислород накапливается (до 130–170% насыщения) в воде и лишь в малой степени уходит в атмосферу и нижележащие слои водоема. Действительно, как показывает расчет, разность между измеренной чистой первичной продукцией днем и наблюдаемым ежедневным увеличением О2, в среднем за первый период достигает лишь 0.021 мг О2/(л · ч). Это тот кислород, который уходит в атмосферу или в нижележащие слои, и он не может оказать существенного влияния на концентрацию О2 в изучаемом поверхностном горизонте водоема.
Сходный характер суточного изменения О2 с максимумом во второй половине дня наблюдался в датском оз. Фредериксборг в конце июня 2004 г. (Staehr, Sand-Jensen, 2007). Аналогичные изменения кислорода отмечены летом в Форелевом и Кристальном озерах в штате Винконсин (Hanson et al., 2008). По нашим данным, представленным на рис. 1б, такая ситуация объясняется возрастанием концентрации кислорода днем до тех пор, пока ВП > Д. Как только ВП/Д станет <1, потребление кислорода превысит его выделение, и концентрация газа начнет снижаться. Рассматриваемая особенность также видна в табл. 2, где содержание кислорода коррелирует с температурой воды и Д и почти не связано с ВП и ФАР.
Однако во второй (циклональный) период процессы ВП и Д оказывают меньшее влияние на газовый состав воды, поскольку происходит интенсивное перемешивание водной массы, и богатые кислородом поверхностные воды уходят вниз. Расчет, аналогичный приведенному выше, показывает, что поток кислорода, который уходит в нижележащие слои водоема или в атмосферу, в этот период днем составляет в среднем 0.087 мг О2/(л · ч), т.е. в ~4 раза больше, чем в первый период. Особенно сильное снижение концентрации кислорода было 22–23 августа (рис. 1б), когда поток кислорода достигал 0.186 мг О2/(л · ч).
Метаболизм планктона и изменение гидрометеорологических условий. Изученная связь между метаболизмом планктона и гидрометеорологическими условиями может быть показана в виде схемы (рис. 3). Отставание максимума температуры воды от максимума солнечной радиации, в течение суток, приводит к соответствующему смещению пиков ВП и Д, которые зависят от них. Превышение ВП над Д в течение дня влечет за собой возрастание концентрации растворенного кислорода, содержание которого начинает снижаться только к вечеру, когда ВП и Д уравниваются.
Рис. 3.
Схема связи метаболизма планктона с гидрометеорологическими показателями: 1 — ФАР, μE/(м2 · с); 2 — температура воды, °С; 3 — ВП; 4 — Д, мг О2/(л · ч); 5 — ВП/Д; 6 — содержание О2, мг/л. Сокращения см. рис. 1.

По-видимому, в других условиях, например при сильном охлаждении и перемешивании водной массы водохранилища осенью, может наблюдаться несколько иной отклик метаболизма планктона.
Представляет интерес также сопоставление внутрисуточных и сезонных изменений метаболизма планктона. В.Г. Девяткин с соавт. (Девяткин и др., 2000, 2012) при изучении Рыбинского водохранилища отмечали, что максимум солнечной радиации в течение года наступает значительно раньше, чем максимум температуры воды. При этом существенного расхождения во времени фотосинтеза и дыхания не происходит – оба параметра коррелируют с температурой воды, и их максимумы наблюдаются в середине лета.
По-видимому, причиной такого отличия от полученных нами результатов является то, что в сезонном аспекте большую роль играют дополнительные факторы – биогенные элементы и биотические взаимодействия, которые в течение суток изменяются мало. Кроме того, влияние солнечной радиации на фотосинтез в рассматриваемые временные интервалы совершенно различно. Это связано с тем, что в течение суток освещенность меняется очень резко (от 0 до 1500 μE/(м2 · с) в нашем исследовании), однако в течение вегетационного сезона изменение среднесуточных значений освещенности происходит плавно (от 50 до 600 μE/(м2 · с)), а для большей части сезона – еще меньше (Девяткин и др., 2012).
Выводы. Выявлено существенное отставание максимума дыхания сообщества днем по сравнению с первичной продукцией, вызванное соответствующим расхождением суточного хода солнечной радиации и температуры воды. Это приводит к смещению максимальных концентраций растворенного кислорода на конец дня. Ухудшение погоды, при котором ФАР уменьшилось в 2.4 раза, а температура воды – почти на 3°С, привело к уменьшению ВП, Д и O2 в ~1.5 раза. Концентрация кислорода снизилась из-за подъема глубинных вод, содержащих малое количество этого газа. Использованное автоматизированное устройство может применяться для непрерывного мониторинга состояния водных объектов, при изучении газообмена в системе водоем–атмосфера.
Список литературы
Винберг Г.Г. 1960. Первичная продукция водоемов. Минск: Изд-во АН БССР.
Девяткин В.Г., Метелева Н.Ю., Митропольская И.В. 2000. Гидрофизические факторы продуктивности литорального фитопланктона: корреляционные связи между гидрофизическими факторами и продуктивностью фито планктона // Биология внутр. вод. № 3. С. 42.
Гончаров А.В., Гречушникова М.Г., Пуклаков В.В. 2018. Новые возможности классического метода: автоматизированное определение первичной продукции и деструкции органического вещества в водоеме // Биология внутр. вод. № 4. С. 107. https://doi.org/10.1134/S0320965218040083
Девяткин В.Г., Метелева Н.Ю., Вайновский П.А. 2012. О роли фитопланктона в формировании кислородного режима водоема в связи с климатическими вариациями. Вода: Химия и экология. № 12. С. 68.
Минеева Н.М. 2009. Первичная продукция планктона в водохранилищах Волги. Ярославль: Принтхаус.
Минеева Н.М., Корнева Л.Г., Соловьева В.В. 2016. Фотосинтетическая активность фитопланктона водохранилищ р. Волги // Биология внутр. вод. № 2. С. 11. https://doi.org/10.7868/S0320965216020121
Пуклаков В.В., Даценко Ю.С., Гончаров А.В. и др. 2015. Гидроэкологический режим водохранилищ Подмосковья (наблюдения, диагноз, прогноз). Москва: Изд-во “Перо”.
Belova S.L., Kremenetskaya E.R. 2010. The effect of water level variations on production destruction processes in the Mozhaisk reservoir // Water Res. V. 34. № 1. P. 70. https://doi.org/10.1134/S0097807810060060
Depew D., Smith R., Guildford S., 2006. Production and Respiration in Lake Erie Plankton Communities // J. Great Lakes Res. V. 32. Issue 4. P. 817.
Goncharov A.V. 2007. Comparison of reservoirs in the Moskva-Vazuza water system in terms of phytoplankton abundance and eutrophication degree // Water Res. V. 34. № 1. P. 78. https://doi.org/10.1134/S0097807807010071
Hanson P.C., Pollard D.L., Bade K. et al. 2004. A model of carbon evasion and sedimentation in temperate lakes // Global Change Biol. V. 10. P. 1285. https://doi.org/10.1111/J.1529-8817.2003.00805.X
Hanson P.C., Carpenter S.R., Kimura N. et al. 2008. Evaluation of metabolism models for free-water dissolved oxygen methods in lakes // Limnol., Oceanogr. Methods 6: P. 454. https://doi.org/10.4319/lom.2008.6.454
Hoellein T.J., Bruesewitz D.A., Richardson D.C. 2013. Revisiting Odum (1956): A synthesis of aquatic ecosystem metabolism // Limnol., Oceanogr. V. 58. № 6. P. 2089. https://doi.org/10.4319/lo.2013.58.6.2089
LaBuhn S., Klump J.V. 2016. Estimating summertime epilimnetic primary production via in situ monitoring in a eutrophic freshwater embayment, Green Bay, Lake Michigan // Journal of Great Lakes Research. V. 42. Issue 5. P. 1026. https://doi.org/10.1016/J.JGLR.2016.07.028
McNair J., Sesselmann M., Kendall S. et al. 2015. Alternative approaches for estimating components of lake metabolism using the free-water dissolved-oxygen (FWDO) method // Fundamental and Applied Limnology. V. 186 (1–2). P. 21. https://doi.org/10.1127/fal/2015/0626
Odum E.P. 1983. Basic ecology. USA: Philadelphia Saunders College Publ.
Staehr P.A., Sand-Jensen K. 2007. Temporal dynamics and regulation of lake metabolism // Limnol., Oceanogr. V. 52 (1). P. 108. https://doi.org/10.4319/lo.2007.52.1.0108
Staehr P.A., Bade D., Van de Bogert M.C. et al. 2010. Lake metabolism and the diel oxygen technique: State of the science // Limnol., Oceanogr. Methods 8. P. 628. https://doi.org/10.4319/lom.2010.8.0628
Staehr P.A., Testa J.M., Kemp W.M. et al. 2011. The metabolism of aquatic ecosystems: history, applications, and future challenges // Aquat. Sci. V. 74. P. 15. https://doi.org/10.1007/s00027-011-0199-2
Vinçon-Leite B., Casenave C. 2019. Modelling eutrophication in lake ecosystems: A review // Science of The Total Environment. V. 651. Part 2. P. 2985. https://doi.org/10.1016/j.scitotenv.2018.09.320
Wetzel R.G. 2001. Limnology. Lake and River Ecosystems. San Diego: Acad. Press.
Wetzel R.G., Likens G.E. 2000. Limnological analyses. New York: Springer.
Williams P., Jle B., Thomas D.N., Reynolds C.S. 2002. Phytoplankton Productivity. Carbon assimilation in marine and freshwater ecosystems. Oxford: Blackwell Science.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод