

Экология, 2020, № 5, стр. 386-391
Анализ факторов, влияющих на скорость поисковых перемещений зарянок Erithacus rubecula во время весенней миграционной остановки
В. Н. Булюк a, *, А. Ю. Синельщикова a, b
a Биологическая станция “Рыбачий”, Зоологический институт РАН
199034 Санкт-Петербург, Университетская наб., 1, Россия
b Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7-9, Россия
* E-mail: victor.bulyuk@mail.ru
Поступила в редакцию 27.11.2019
После доработки 14.02.2020
Принята к публикации 18.02.2020
Аннотация
Зарянки после завершения ночного миграционного полета утром начинают совершать исследовательские перемещения в поисках места остановки для отдыха и пополнения энергетических ресурсов. Данные по поимкам 111 зарянок во время весенней миграции на Куршской косе Балтийского моря показали, что скорость перемещений птиц во время поиска не зависит от температуры воздуха, скорости ветра, осадков и числа конспецифичных особей на остановке. В небольшой степени на нее влияют возраст и энергетическое состояние птиц, а также календарная дата. Предполагается, что наиболее сильное влияние на скорость поисковых перемещений зарянок весной оказывают эндогенные программы поиска места остановок, которые у отдельных птиц сильно различаются.
Большинство воробьиных – ночных мигрантов – во время сезонных миграций чередуют ночной полет с дневными остановками для пополнения энергетических запасов. В период миграции ночные мигранты обычно проводят на остановках около 90% времени, потраченного на миграцию, а непосредственно в миграционном полете около 10%. В энергетическом выражении расходы собственно на полет составляют лишь около трети расхода энергии во время миграции, а две трети энергии расходуется в ходе миграционных остановок [1, 2]. Чтобы найти место для остановки, птицы после ночного броска совершают поисковые перемещения [3–7]. Исследования факторов, влияющих на поведение и пространственное распределение птиц во время такого поиска на дневных остановках, имеют важное значение для понимания принципов организации миграции на уровне отдельных видов [6].
Наблюдения на миграционных остановках на Куршской косе Балтийского моря за европейскими зарянками Erithacus rubecula, помеченными кольцами и радиопередатчиками, показали, что весной в первый день после прибытия на остановку птицы перемещаются в разных направлениях. Масштаб этих перемещений редко превышает несколько сотен метров [7, 8].
Мы попытались выяснить, как влияют на скорость поисковых перемещений зарянок такие погодные факторы, как температура воздуха, скорость ветра и осадки. Эти погодные параметры могут влиять не только на активность и количество насекомых, которых потребляют зарянки, но и на успешность добывания птицами корма во время поиска подходящего биотопа для постоянного места остановки. Возможно, что при низких температурах, спокойной погоде и отсутствии осадков скорость передвижения птиц во время поиска места остановки будет выше, чем при высоких температурах, сильном ветре и дожде. Мы проверили, зависит ли скорость перемещений зарянок после прилета на остановку от их возраста и энергетического состояния, количества особей этого вида на остановке, разной продолжительности светлой части дня в течение сезона. Ожидалось, что взрослые птицы, имеющие больший опыт выбора мест остановки и более высокий социальный статус, и жирные птицы с низкой мотивацией пополнения энергетических ресурсов будут перемещаться медленнее, чем молодые и тощие особи. Большое количество конспецифичных особей на остановке может снизить скорость перемещений мигрантов из-за увеличения частоты конфликтов или, наоборот, способствовать ее повышению, в случае если птицы будут пытаться найти место с минимальной конкуренцией за пищевые ресурсы. С увеличением продолжительности светового дня в ходе весеннего сезона можно было ожидать уменьшение подвижности птиц.
МАТЕРИАЛ И МЕТОДЫ
Наше исследование основано на результатах отлова зарянок на остановках во время весенней миграции на мысе Росситтен на Куршской косе Балтийского моря. Благодаря географическому положению, большому разнообразию биотопов, удобных для сбора корма и укрытия от хищников, здесь после ночных полетов во время сезонных миграций останавливается большое количество зарянок. В качестве меры скорости перемещений зарянок на остановке мы использовали временной интервал между выпуском птиц, пойманных в первые два утренних часа, и их последующим отловом в тот же день на фиксированном расстоянии – в нескольких сотнях метров от места выпуска.
Зарянки были пойманы в 2004–2017 гг. на полевом стационаре Биологической станции “Рыбачий” (55°09′ с.ш., 20°51′ в.д.). Его территория покрыта кустарниками (в основном ивы Salix sp.), с небольшим количеством деревьев и участков с тростниковыми зарослями. Птицы были отловлены стандартными паутинными сетями (2 × 7 м), расположенными в две линии (общая длина 175 м). Сетки проверяли каждый час от рассвета до заката. За 14 лет в первые два утренних часа во время весенней миграции (27 марта–30 апреля) было поймано 111 зарянок, которые в этот же день были повторно отловлены в контрольной точке на расстоянии 270–300 м от места выпуска. Для каждой пойманной в сети птицы отмечали время поимки, длину крыла (с точностью до 0.5 мм), массу тела (с точностью до 0.1 г), возраст (взрослые – птицы старше одного года; молодые – птицы, родившиеся в предшествующий поимке год [9]).
Погодные параметры в день отлова измеряли в месте проведения исследования. Для оценки температуры воздуха и скорости ветра были использованы средние значения этих параметров в 12:00 и 18:00 по восточно-европейскому летнему времени (GMT + 3), для оценки влияния дождя – данные по количеству осадков (в мм), выпавших в дату отлова в светлую часть суток. Температура воздуха во время поимок зарянок колебалась в пределах 2.3–17.8° C, скорость ветра – 0.2–7.4 м с–1, количество осадков – 0–9.0 мм.
Число птиц на остановке в даты их отлова оценивали по общему количеству зарянок, пойманных за день на стационаре, – оно варьировало от 3 до 292 особей. Календарная дата (КД) от начала к концу весеннего сезона варьировала от 1 до 35 с увеличением дат от 27 марта до 30 апреля, когда соответственно были получены первые и последние данные по зарянкам.
Для оценки энергетического состояния отдельных зарянок был использован индекс состояния тела (ИСТ, г мм–1), который рассчитывали путем деления массы тела птицы в первую утреннюю поимку на ее длину крыла. Несмотря на то, что у зарянок отмечается сильная линейная зависимость между весом тела и визуально оцененными запасами подкожного жира [10–13], мы предпочли использовать ИСТ, поскольку он (i) учитывает структурный размер птиц, (ii) отражает не только запасы жира, но и запасы белка, (iii) не зависит от ошибок в определении подкожных жировых запасов между отдельными кольцевателями. ИСТ зарянок, которых мы использовали для анализа, был умеренно коррелирован с их жировыми запасами, оцененными по 9-балльной системе (коэффициент ранговой корреляциия Спирмена: rs = 0.61, р < 0.0001, n = 111): у отдельных птиц он варьировал от 0.185 до 0.255 г мм–1.
Влияние рассмотренных нами факторов на скорость поисковых перемещений птиц на миграционной остановке в течение весеннего дня изучали с помощью множественного регрессионного анализа. Разница во времени в часах между утренним выпуском птицы и ее последующей поимкой в тот же день в контрольной точке в 270–300 м была зависимой переменной. Независимыми переменными были: 1) возраст, 2) КД, 3) число пойманных зарянок, 4) ИСТ, 5) температура воздуха, 6) скорость ветра, 7) количество выпавших осадков. Возраст и количество выпавших осадков были категориальными переменными (0 – молодые, 1 – взрослые птицы; 0 – количество осадков 0–0.5 мм, 1 – более 0.5 мм), в то время как остальные переменные были непрерывными. Тест Колмогорова-Смирнова на нормальное распределение показал, что распределения переменных “число птиц на остановке” и “скорость ветра” значимо отличались от нормального. Логарифмическое преобразование этих переменных привело к их нормальному распределению. Выбор модели осуществлялся пошаговым исключением предикторов. Переменная вводилась в модель, если наблюдаемый уровень значимости ее F-значения был меньше заданного порога включения (0.05), и исключалась, если этот уровень значимости был больше порога исключения (0.1). В результате выбрали модель, в которой все переменные были значимыми.
Все приведенные в тексте средние значения включали стандартное отклонение.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
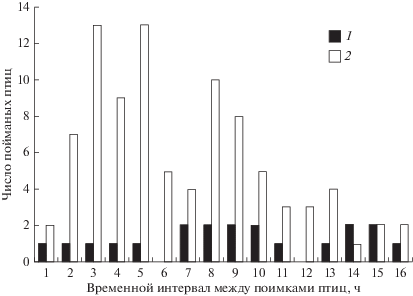
Весной пойманные и выпущенные на миграционной остановке в первые два утренних часа зарянки были повторно отловлены на расстоянии 270–300 м от места выпуска через 1–16 ч (в среднем 7.1 ± 3.9 ч, n = 111; рис. 1 ). Молодые птицы имели значимо более высокую скорость перемещений, чем взрослые. Время между выпуском и последующим отловом в контрольной точке у молодых птиц было в среднем 6.7 ± 3.8 ч (n = 91), тогда как у взрослых 9.0 ± 4.5 ч (n = 20) (t-тест: t = 2.39, df = 109, p = = 0.018). Сравнивая средние значения ИСТ, КД, число зарянок на остановке и три рассматриваемых погодных параметра при поимке молодых и взрослых птиц, мы не нашли достоверных различий между двумя группами. Это позволяет предполагать, что различия в скорости перемещений между молодыми и взрослыми птицами были обусловлены их различным поведением.
Рис. 1.
Число взрослых (1) и молодых (2) зарянок, пойманных впервые рано утром во время поисковых перемещений, а затем в тот же день через разные временные интервалы на расстоянии 270–300 м от места выпуска.
В результате регрессионного анализа была получена модель с тремя значимыми предикторами:
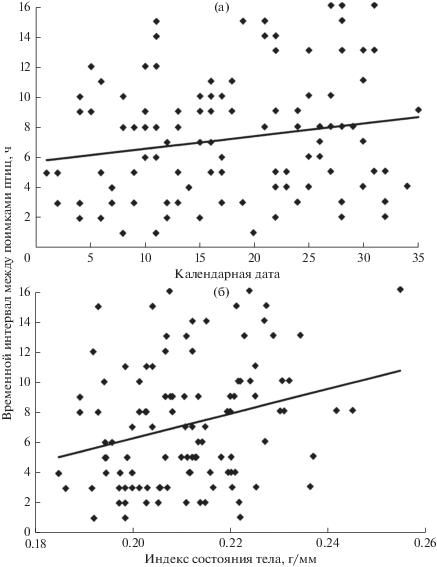
Полученная достоверная положительная связь индекса состояния тела и календарной даты со скоростью перемещений зарянок на остановке свидетельствует о том, что чем меньше птицы имели энергоресурсов и чем меньше была продолжительность дня во время остановки, тем быстрее они передвигались (рис. 2 ).
Рис. 2.
Зависимость скорости поисковых перемещений зарянок, выраженной временным интервалом между поимкой птиц рано утром и их повторной поимкой в 270–300 м от места выпуска в тот же день, от календарной даты (а) , где 1 – 27 марта, 2 – 28 марта и т.д.; б – индекс состояния тела.
Во многих случаях целью дневных перемещений воробьиных – ночных мигрантов на остановках, после завершения ночного полета является поиск безопасных мест для отдыха и кормежки. До недавнего времени по методическим причинам сведения о том, как особи мелких мигрирующих ночью воробьиных после ночного броска осуществляют выбор места остановки и какие факторы при этом влияют на его поиск, отсутствовали или были крайне ограничены. В последние несколько десятилетий, благодаря массовому кольцеванию, анализу данных повторных отловов окольцованных птиц и прослеживанию отдельных особей с помощью микропередатчиков, появилось большое количество новых и интересных фактов об этом явлении (см. обзор [6]).
Мы исследовали, как влияют на подвижность зарянок во время поиска места остановки днем такие факторы, как их возраст и энергетическое состояние, количество птиц этого вида на остановке, погода (температура воздуха, скорость ветра и количество осадков), различия в продолжительности дня. Несмотря на то, что для птиц этого вида имеется большое количество данных по экологии и поведению на миграционных остановках [6–8, 14–17], сведения о влиянии внешних и эндогенных факторов на их подвижность во время остановки практически отсутствовали. Мы использовали данные по времени перемещений достаточно большого числа зарянок, которые сначала впервые были пойманы рано утром, а затем в тот же день повторно отловлены в контрольной точке (270–300 м от места выпуска). Наблюдения на Росситенском мысу показали, что место, где проводились исследования, было крайне привлекательным для посадки зарянок после завершения ночного миграционного полета, и птицы начинали совершать дневные перемещения в ранние утренние часы, связанные с поиском места остановки и кормежки [7, 8].
Мы не обнаружили, что температура воздуха, скорость ветра и осадки влияют на скорость дневных передвижений зарянок во время поисковых перемещений на миграционной остановке весной после завершения ночного миграционного полета. Зарянки на остановках в этот сезон питаются в основном пауками, многоножками, жесткокрылыми, муравьями, паразитическими перепончатокрылыми [18], которых они собирают на земле. Погодные условия, возможно, не очень сильно влияют на количество и активность объектов питания зарянок, а также на их доступность для птиц во время питания при перемещениях в поиске постоянного места остановки. Подвижность зарянок на остановке также не зависела от количества конспецифичных особей. Во время поисковых перемещений после ночного полета птицы, скорее всего, стараются избегать конфликтных ситуаций, чтобы не тратить на них время и энергию.
С увеличением длины светового дня скорость поисковых перемещений зарянок уменьшалась (см. рис. 2). Это могло быть вызвано как улучшением кормовых условий с прогрессом весеннего сезона, так и тем, что к концу весенней миграции мотивация поиска благоприятного места для остановки могла снижаться в связи с завершением миграционного состояния у отдельных особей.
По данным наблюдений в естественных условиях [4, 6, 19–21] известно, что во время сезонных миграций тощие воробьиные – ночные мигранты, днем часто проявляют более высокую двигательную активность, чем птицы с высокими запасами жира. Наши данные подтверждают такую связь для зарянок во время поисковых перемещений: птицы с низкими ИСТ были более подвижными, чем с высокими (см. рис. 2 ). Еще одним фактором, влияющим на подвижность зарянок, был возраст птиц (см. рис. 1 ). Тот факт, что взрослые зарянки на остановке были в среднем менее подвижными, чем молодые, согласуется с гипотезой о том, что социальное доминирование и предыдущий опыт, накопленный взрослыми птицами, оказывают определенное влияние на их мобильность [22]. Подобное поведение у зарянок отмечается уже на зимовках [23, 24]. К сожалению, из-за того, что осенью на остановках на Куршской косе встречаются в основном молодые зарянки [8, 25], мы не смогли проверить, как различия в возрасте зарянок в этот сезон сказываются на их подвижности. В осенний сезон в исследуемом районе для молодых зарянок мы не нашли ни одного фактора, включая погодные параметры, число птиц на остановке, различия в продолжительности светового дня и энергетическом состоянии мигрантов, которые бы значимо влияли на скорость их перемещений после ночного миграционного полета.
ЗАКЛЮЧЕНИЕ
Полученная нами модель подвижности зарянок, выраженная через время преодоления 270–300-метрового фиксированного участка на весенней миграционной остановке, имела низкую долю дисперсии зависимой переменной (15.2%). Отсутствие связи скорости поисковых перемещений с температурой воздуха, скоростью ветра, осадками, числом птиц на остановке и очень слабое влияние на подвижность птиц таких факторов, как возраст и энергетическое состояние мигрантов, продолжительность светлой части суток, указывают на то, что на поведение птиц во время поисковых перемещений могут оказывать влияние другие, более сильные факторы.
Мы предполагаем, что одним из таких наиболее важных факторов могут быть различия в эндогенных программах поиска места остановок у отдельных птиц. Об этом явно свидетельствуют результаты радиотелеметрического прослеживания зарянок на Куршской косе, которые показали большое разнообразие поведения птиц в первый день миграционной остановки. По данным мониторинга за перемещениями зарянок с передатчиками, одни птицы днем после ночного полета смещались на определенное расстояние, а затем до конца дня перемещались на ограниченной территории – временном индивидуальном участке. Другие особи после начального периода направленного движения и оседания на индивидуальном участке совершали новое направленное движение с последующим оседанием на втором индивидуальном участке либо двигались вплоть до темноты [8].
В последнее время появилось много фактов о том, что индивидуальные особенности птиц оказывают значительное влияние на поведение зарянок не только во время поиска места остановки, но и на продолжительность остановок, начало и завершение ночного миграционного полета [6, 8, 14, 15, 17]. Чтобы в той или иной степени оценить влияние эндогенных пространственно-временных программ на подвижность птиц в местах остановок, как, впрочем, и на многие другие формы поведения птиц во время миграции, необходимо знать предшествующую миграционную историю каждой отдельной особи. Эта информация должна включать в себя данные о том, когда птица начала миграцию, на каком участке пути относительно всей трассы миграции она находится, в каких ландшафтных и погодных условиях проходил ее ночной полет до остановки. К сожалению, в настоящее время получить такие сведения пока не представляется возможным. Однако есть надежда, что такая возможность вскоре появится с развитием новых технологий в радиотелеметрии, позволяющих длительное время и на больших дистанциях прослеживать небольших по размеру мигрирующих птиц [26].
Мы благодарим всех наших коллег, студентов и волонтеров, которые участвовали в отловах и кольцевании птиц на полевом стационаре Биологической станции “Рыбачий” Зоологического института РАН. Мы признательны двум анонимным оппонентам, чьи замечания позволили значительно улучшить текст. Данная работа была выполнена при поддержке Зоологического института РАН (зарегистрированный исследовательский проект № АААА-A19-119021190073-8) и гранта РФФИ (№ 19-44-390002 р-а).
Список литературы
Hedenstrom A., Alerstam T. Optimal fuel loads in migratory birds: distinguishing between time and energy minimization // J. Theor. Biol. 1997. V. 189. P. 227–234.
Wikelski M., Tarlow E.M., Raim A. et al. Cost of migration in free-flying songbirds // Nature. 2003. V. 423. P. 704.
Aborn D.A., Moore F.R. Pattern of movement by summer tanagers (Piranga rubra) during migratory stopover: a telemetry study // Behaviour. 1997. V. 134. P. 1077–1100.
Moore F.R., Aborn D.A. Mechanisms of en route habitat selection: how do migrants make habitat decisions during stopover? // Stud. Avian Biol. 2000. V. 20. P. 34–42.
Chernetsov N. Spatial behavior of medium and long-distance migrants at stopovers studied by radio tracking // Annals New York Acad. Sci. 2005. V. 1046. P. 242–252.
Chernetsov N. Passerine migration. Stopovers and flight. Berlin: Springer, 2012. 184 p.
Chernetsov N., Mukhin A. Spatial behaviour of European robins Erithacus rubecula during migratory stopovers: a telemetry study // Wilson J. Ornithol. 2006. V. 118. P. 364–373.
Цвей А.Л. Стратегии миграции зарянки (Erithacus rubecula) в восточной Прибалтике: Дис. канд. биол. наук. СПб: ЗИН РАН, 2008. 201 с.
Svensson L. Identification guide to passerines. Fourth, revised and enlarged edition. Stockholm, 1992. 368 p.
Pettersson J., Hasselquist D. Fat deposition and migration capacity of robins Erithacus rubecula and goldcrests Regulus regulus at Ottenby, Sweden // Ring. & Migr. 1985. V. 6. P. 66–76.
Karlsson L., Persson K., Pettersson J., Walinder G. Fat-weight relationship and migratory strategies in the Robin Erithacus rubecula at two stopover sites in South Sweden // Ring. & Migr. 1988. V. 9. P. 160–168.
Ehnbom S., Karlsson L., Ylven R., Akesson S. A comparison of autumn migration strategies in robins, Erithacus rubecula, at a coastal and an inland site in southern Sweden // Ring. & Migr. 1993. V. 14. P. 94–93.
Zimin V.B. Body mass variability in juvenile Robins Erithacus rubecula in the Ladoga area // Avian Ecol. Behav. 2003. V. 10. P. 1–31.
Bulyuk V.N., Tsvey A. Timing of nocturnal autumn migratory departures in juvenile European robins (Erithacus rubecula) and endogenous and external factors // J. Ornithol. 2006. V. 147. P. 298–309.
Bulyuk V.N., Tsvey A. Regulation of stopover duration in the European Robin Erithacus rubecula // J. Ornithol. 2013. V. 154. P. 1115–1126.
Bolshakov C.V., Chernetsov N., Mukhin A. et al. Time of nocturnal departures in European robins, Erithacus rubecula, in relation to celestial cues, season, stopover duration and fat stores // Anim. Behav. 2007. V. 74. P. 855–865.
Bulyuk V.N. Influence of fuel load and weather on timing of nocturnal spring migratory departures in European robins, Erithacus rubecula // Behav. Ecol. and Sociobiol. 2012. V. 66. P. 385–395.
Чернецов Н.С., Титов Н.В. Питание и стратегии весенней миграции зарянки, Erithacus rubecula (Aves, Turdidae), в юго-восточной Прибалтике // Зоол. журн. 2003. Т. 82. Вып. 12. С. 1525–1529.
Dierschke V. Predation hazard during migratory stopover: are light or heavy birds under risk? // J. Avian Biol. 2003. V. 24. P. 24–29.
Wang Y., Moore F.R. Long-distance bird migrants adjust their foraging behavior in relation to energy stores // Acta Zool. Sin. 2005. V. 51. P. 12–23.
Cohen E.B., Moore F.R., Fischer R.A. Fuel stores, time of spring, and movement behavior influence stopover duration of Red-eyed Vireo Vireo olivaceus // J. Ornithol. 2014. V. 155. P. 785–795.
Woodrey M. Age-dependent aspects of stopover biology of passerine migrants // Study Avian Biol. 2000. V. 20. P. 43–52.
Telleria J.L., Perez-Tris J., Ramirez A. et al. Distribution of Robins Erithacus rubecula in wintering grounds: effects of the conspecific density, migratory status and age // Ardea. 2001. V. 89. P. 363–373.
Catry P., Campos A., Almanda V., Cresswell W. Winter segregation of migrant European robins Erithacus rubecula in relation to sex, age and size // J. Avian Biol. 2004. V. 35. P. 204–209.
Bulyuk V.N. Age structure of Robins Erithacus rubecula at the eastern Baltic coast during autumn migration and wind conditions // Avian Ecol. Behav. 2013. V. 23. P. 23–40.
Wikelski M., Kays R.W., Kasdin N.J. et al. Going wild: What a global small-animal tracking system could do for experimental biologists // J. Experim. Biol. 2007. V. 210. P. 181–186.
Дополнительные материалы отсутствуют.