

Экология, 2020, № 5, стр. 332-340
Ресурсная регуляция отавности растений альпийских пустошей: факторный эксперимент
М. М. Эркенова a, b, Д. К. Текеев c, Р. Б. Семенова a, А. А. Ахметжанова d, В. Г. Онипченко b, c, d, *
a Северо-Кавказская государственная академия
369001 КЧР, Черкесск, ул. Ставропольская, 36, Россия
b Карачаево-Черкесский госуниверситет им. У. Алиева
369200 КЧР, Карачаевск, ул. Ленина, 29, Россия
c Тебердинский государственный природный биосферный заповедник
369210 КЧР, Теберда, Бадукский переулок, 1, Россия
d Московский госуниверситет им. М.В. Ломоносова
119234 Москва, Ленинские горы, 1-12, Россия
* E-mail: vonipchenko@mail.ru
Поступила в редакцию 11.12.2019
После доработки 16.01.2020
Принята к публикации 18.03.2020
Аннотация
Отрастание надземной биомассы отдельных видов растений после полного или частичного отчуждения (дефолиации) в высокогорьях может зависеть от конкуренции с соседними растениями и степени обеспеченности почвенными ресурсами, в первую очередь элементами минерального питания (особенно N и P). Проведен 3-летний полнофакторный эксперимент по влиянию трех режимов дефолиации (контроль, 1 и 2 укоса на уровне почвы за сезон), обогащению почвы N и P и удалению соседних растений (снижение конкуренции) на численность побегов и надземную биомассу 3 видов растений альпийских пустошей (Anemone speciosa, Festuca ovina и Oxytropis kubanensis). Всего 12 вариантов, повторность 10-кратная. Показано, что в условиях низкопродуктивных сообществ альпийских пустошей снятие конкуренции в отличие от внесения NP удобрений, не улучшает восстановление биомассы после дефолиации. Отавность азотфиксирующего бобового растения не зависела от доступности почвенных ресурсов.
В формировании травяных растительных сообществ, в том числе высокогорных, большая роль принадлежит фитофагам, отчуждающим значительную часть надземной первичной продукции [1–5]. Способность восстанавливать надземную биомассу после повреждений (отавность – см. [6, 7]) – одно из фундаментальных адаптивных свойств растений, используемое при выделении их функциональных типов [8]. Эта способность имеет большое адаптивное значение для поддержания популяций растений в условиях сенокошения, выпаса и других зоогенных нарушений [9], является очень важным признаком, определяющим доминирование видов [10]. Отавность растений зависит как от свойств растительного организма (жизненной формы, структуры надземных и подземных побегов и др.) [11], так и от доступности почвенных ресурсов, обеспечивающих рост [12–14]. К таким ресурсам относятся прежде всего вода и элементы минерального питания (ЭМП), поступление которых зависит и от конкуренции со стороны соседних растений. Вопрос, насколько отавность высокогорных растений определяется непосредственным недостатком ЭМП и конкуренцией, важен для понимания функционирования альпийских фитоценозов, но он остается практически не исследованным.
Альпийские сообщества Кавказа во многом сформированы под воздействием крупных фитофагов, к которым относится домашний скот (преимущественно овцы, лошади, крупный рогатый скот, в последнее время включающий яков) [15, 16], а также дикие копытные (серны Rupicapra rupicapra, туры Capra caucasica, кабаны Sus scrofa), особенно на охраняемых природных территориях [17, 18]. Альпийские луга Кавказа и Альп во многом сформировались под воздействием выпаса, длительное отсутствие которого ведет к снижению роли травянистых растений и усилению роли кустарников, кустарничков, мхов и лишайников [19], т.е. переходу к сообществам, сходным с горными–тундрами и редколесьями, которые широко распространены в горах Сибири и Дальнего Востока. Сильный выпас оленей в тундрах Норвегии приводит к смене кустарничковой тундры злаковыми сообществами [20], а исключение выпаса в тундрах Аляски резко увеличивало участие лишайников [21].
Одним из типичных растительных сообществ Кавказа являются низкопродуктивные альпийские лишайниковые пустоши (АЛП), развивающиеся на гребнях и обдуваемых склонах гор в условиях незначительного (или отсутствующего) снежного покрова зимой и сильного промерзания почв [18, 22, 23]. Несмотря на суровые условия, в этих флористически богатых сообществах выражена конкуренция между основными доминантами, в то время как между доминирующими и недоминирующими видами растений отмечены положительные связи [24, 25]. Среди растений АЛП абсолютно преобладает стратегия стресс-толерантности [26], т.е. низкая относительная скорость роста и относительно больший вклад ассимилянтов в защитные структуры [8]. В то же время значительная доля подземной биомассы у высокогорных растений [22, 27] делает их устойчивыми к отчуждению надземной биомассы, что было экспериментально показано и для растений альпийских пустошей [28]. Проведенные в предшествующие годы (2001–2006) исследования отавности растений альпийских пустошей не затрагивали вопросов ее ресурсного лимитирования. Они показали, что отавность альпийских растений после одно-, дву- и трехкратного отчуждения надземной биомассы за сезон в целом не компенсировала удаляемую биомассу [28]. Также установлено, что восстановление биомассы уменьшалось при увеличении частоты скашивания, и от первого года эксперимента к третьему численность побегов после дефолиации в целом восстанавливается лучше, чем масса отдельных побегов, а растения альпийских пустошей отличались несколько большей отавностью по сравнению с растениями пестроовсяницевых лугов [28]. Однако зависимость восстановления биомассы от доступности почвенных ресурсов и/или интенсивности конкуренции ранее не была исследована в альпийских фитоценозах.
Продукция альпийских сообществ, как показали результаты серии экспериментов [29, 30], обычно лимитирована доступностью ЭМП в почве, в первую очередь азота и во вторую фосфора. Мы предположили, что как внесение в почву ЭМП, так и снижение конкуренции (а также взаимодействие этих факторов) должны повышать способности растений к восстановлению надземной биомассы после отчуждения (отавность), но реакции растений разных экологических групп на эти факторы могут быть различными. Проверка этой гипотезы и составила основную цель нашей работы. В предлагаемой статье обсуждаются результаты исследований отавности трех видов цветковых растений в факторном эксперименте с внесением ЭМП, удалении соседей (снижении конкуренции) при разных режимах дефолиации.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Полевой эксперимент. Работа проведена на территории Тебердинского государственного биосферного заповедника на стационаре “Малая Хатипара”. В 2009 г. были заложены экспериментальные площадки в альпийских лишайниковых пустошах и предварительно проведен подсчет побегов (вегетативных и генеративных) до экспериментального воздействия. Исследованы три вида травянистых растений – Anemone speciosa (Ranunculaceae), Festuca ovina (Poaceae), Oxytropis kubanensis (Fabaceae) (номенклатура по [31]).
Для каждого вида было заложено 12 вариантов эксперимента в 10-кратной повторности каждого (всего 3 вида × 12 вариантов × 10 повторностей = = 360 круглых площадок). Каждая площадка была огорожена от соседних металлическим кольцом из алюминиевой проволоки на уровне почвы и отмечена биркой с номером варианта. Площадь одной площадки – 314 см2. Для изучения способности растений восстанавливать надземную биомассу после повреждений имитировали отчуждение биомассы путем среза побегов. Перед срезанием проводился подсчет вегетативных и генеративных побегов. Однократное срезание тестовых растений было в конце июля, двукратное – в конце июля и конце августа (табл. 1). Срезание проводили ножницами у поверхности почвы. Укосы помещали в бумажные пакеты, подсушивали на воздухе, а затем в сушильных шкафах при температуре 80°С в течение 7–8 ч. Удобрения вносили каждый год после схода снега с площадок по всей площади круга в сухом виде. В конце июня–начале июля (табл. 2) внесен двойной суперфосфат Ca(H2PO4)2 из расчета 0.5 г в каждый круг и мочевина CO(NH2)2 – 0.6 г в кольцо (расчетные дозы 43 кг/га Р и 90 кг/га N). В варианте “удаление соседей” проводили многократное срезание соседних с тестовым видом растений.
Таблица 1.
Сроки проведения укосов
| Год | Вид | ||
|---|---|---|---|
| Oxytropis kubanensis | Festuca ovina | Anemone speciosa | |
| 2010 | |||
| 1-й укос | 2 августа | 3–4 августа | 1 августа |
| 2-й укос | 21 августа | 20 августа | 20–21 августа |
| 2011 | |||
| 1-й укос | 11–12 августа | 9–10 августа | 8–9 августа |
| 2-й укос | 8 сентября | 8 сентября | 8 сентября |
| 2012 | |||
| 1-й укос | 2 августа | 5 августа | 2 августа |
| 2-й укос | 3 сентября | 3 сентября | 2 сентября |
| 2013 | 24–25 июля | 30–31 июля | 24–30 июля |
Таблица 2.
Сроки внесения элементов минерального питания
| Год | Oxytropis kubanensis | Festuca ovina | Anemone speciosa |
|---|---|---|---|
| 2010 | 26–27 июня | 28 июня | 27 июня |
| 2011 | 26 июня | 26 июня | 26 июня |
| 2012 | 2 июля | 2 июля | 2 июля |
Для оценки итоговой отавности в 2013 г. (без экспериментального воздействия) был проведен подсчет вегетативных и генеративных побегов и однократное срезание надземной биомассы во всех вариантах, включая контроль. Таким образом, результаты учитывали по следующим характеристикам: 1) изменение численности вегетативных и генеративных побегов в 2010–2013 г.; 2) биомасса отдельных укосов и итоговая биомасса по каждому варианту в 2013 г.
Статистический анализ. Влияние исследуемых факторов на отрастание биомассы и восстановление численности побегов оценивали для каждого вида индивидуально. Эта оценка основана на итоговой биомассе и численности на следующий год (2013) после трехлетнего эксперимента. После логарифмической трансформации (по основанию натурального логарифма) распределение величин биомассы не отличалось от нормального (критерий Колмогорова-Смирнова) и позволило провести 3-факторный дисперсионный анализ. Анализируемые независимые факторы: частота укосов (У: 0 – контроль, 1 – один укос за сезон, 2 – два укоса за сезон), внесение элементов минерального питания – удобрений (ЭМП: 0 – без внесения, 1 – с внесением), удаление соседей (С: 0 – с удалением, 1 – без удаления), зависимая переменная – надземная биомасса. В связи с сильным отклонением от нормальности значений численности в 2013 г., численности и биомассы укосов в предшествующие годы, а также практическим отсутствием взаимодействия между факторами по результатам дисперсионного анализа биомассы влияние каждого фактора было оценено также индивидуально непараметрическим однофакторным дисперсионным анализом (Kruskal-Wallis ANOVA). Анализ для данных 2010 г. (год начала эксперимента) проведен для подтверждения исходной однородности выборки. Все расчеты произведены в программе Statistica 12.0.
РЕЗУЛЬТАТЫ
В год начала экспериментального воздействия (2010) заложенные площадки для разных вариантов значимо не различались между собой по численности побегов (табл. 3–5). Исключение составляли численность вегетативных побегов и суммарная их численность, которая показала значимые различия у Festuca ovina по фактору частоты скашивания (численность исходно была значительно выше на “укосных” вариантах по сравнению с контрольными), а также численность вегетативных побегов по этому же фактору у Anemone speciosa (она была слегка меньше в 2-укосном варианте).
Таблица 3.
Значимость влияния факторов эксперимента на численность побегов и надземную биомассу Festuca ovina (Kruskal-Wallis ANOVA). Полужирным шрифтом выделено значимое влияние (p < 0.05), курсивом – пограничные величины (0.05 < p < 0.1)
| Год | Показатель | Значимость влияния фактора (p) | ||
|---|---|---|---|---|
| удобрение | соседи | укос | ||
| 2010 | Численность генеративных побегов | 1.000 | 1.000 | 0.387 |
| Численность вегетативных побегов | 0.273 | 0.273 | 0.0006 | |
| Суммарная численность побегов | 0.715 | 0.715 | 0.0001 | |
| 2011 | Численность генеративных побегов | 0.0018 | 0.854 | <0.0001 |
| Численность вегетативных побегов | 0.715 | 0.273 | 0.202 | |
| Суммарная численность побегов | 0.144 | 0.144 | 0.0716 | |
| Биомасса 1-го укоса | <0.0001 | 0.655 | 0.670 | |
| Биомасса 2-го укоса | 0.752 | 0.0267 | – | |
| 2012 | Численность генеративных побегов | <0.0001 | 0.846 | <0.0001 |
| Численность вегетативных побегов | 0.144 | 0.0285 | 0.0608 | |
| Суммарная численность побегов | 0.0285 | 0.144 | 0.1225 | |
| Биомасса 1-го укоса | <0.0001 | 0.655 | 0.670 | |
| Биомасса 2-го укоса | 0.0578 | 0.527 | – | |
| 2013 | Численность генеративных побегов | 0.0001 | 0.8541 | <0.0001 |
| Численность вегетативных побегов | <0.0001 | 0.1441 | <0.0001 | |
| Суммарная численность побегов | <0.0001 | 0.1003 | 0.0000 | |
| Биомасса | 0.0001 | 0.259 | 0.0001 | |
Все изученные факторы действовали на отрастание растений практически независимо, поскольку 3-факторный дисперсионный анализ не показал значимых взаимодействий между ними (табл. 1е – электронное приложение). Лишь для Festuca ovina отмечено взаимодействие “удаление соседей” × “частота скашивания” (p = 0.010), при этом на площадках, косимых дважды, восстановление биомассы шло лучше в присутствии соседей, чем при их отсутствии. Это свидетельствует о положительном влиянии фитоценотического окружения на рост растений этого вида при дефолиации. Рассмотрим влияние изученных факторов на численность побегов и биомассу модельных растений.
Фактор дефолиации (срезания надземных побегов) после 3 лет эксперимента оказал значимое негативное влияние на все исследованные параметры растений, за исключением численности вегетативных побегов у Oxytropis kubanensis, которая в отличие от биомассы значимо не снижалась даже в варианте двукратной дефолиации за сезон. Наиболее резко отчуждение надземной биомассы сказалось на численности генеративных побегов, которая многократно снизилась (табл. 3–5, рис. 1а). Например, численность генеративных побегов в 2013 г. (восстановление после 3 лет эксперимента) в контроле составила 4.9 ± 1.9 (среднее и его ошибка, N = 10, здесь и далее) на площадку, в варианте только с двукратным скашиванием генеративные побеги полностью отсутствовали (табл. 2е – электронное приложение). После 3 лет дефолиации более сильное снижение биомассы отмечено у Festuca ovina (при одном срезании примерно в 4 раза), а наиболее устойчивым оказалась Anemone speciosa (снижение биомассы примерно на треть) (табл. 2е – электронное приложение, рис. 1б).
Таблица 4.
Значимость влияния факторов эксперимента на численность побегов и надземную биомассу Anemone speciosa (Kruskal-Wallis ANOVA). Полужирным шрифтом выделено значимое влияние (p < 0.05)
| Год | Показатель | Значимость влияния фактора (p) | ||
|---|---|---|---|---|
| удобрение | соседи | укос | ||
| 2010 | Численность генеративных побегов | 0.2709 | 0.7136 | 0.6676 |
| Численность вегетативных побегов | 0.5834 | 0.3607 | 0.0447 | |
| Суммарная численность побегов | 0.7148 | 0.2731 | 0.3554 | |
| 2011 | Численность генеративных побегов | 1.000 | 0.4561 | 0.0172 |
| Численность вегетативных побегов | 0.1997 | 0.0992 | 0.7906 | |
| Суммарная численность побегов | 0.0679 | 0.1441 | 1.000 | |
| Биомасса 1-го укоса | 0.0253 | 0.654 | 0.905 | |
| Биомасса 2-го укоса | 0.0331 | 0.361 | – | |
| 2012 | Численность генеративных побегов | 0.464 | 0.714 | 0.194 |
| Численность вегетативных побегов | 0.166 | 0.843 | 0.962 | |
| Суммарная численность побегов | 0.439 | 0.121 | 0.490 | |
| Биомасса 1-го укоса | 0.0003 | 0.180 | 1.000 | |
| Биомасса 2-го укоса | 0.0016 | 0.527 | – | |
| 2013 | Численность генеративных побегов | 0.0053 | 0.5774 | 0.0001 |
| Численность вегетативных побегов | 0.3580 | 0.5813 | 0.0091 | |
| Суммарная численность побегов | 0.0001 | 0.6733 | 0.0072 | |
| Биомасса итоговая | 0.0001 | 0.159 | 0.0001 | |
Таблица 5.
Значимость влияния факторов эксперимента на численность побегов и надземную биомассу Oxytropis kubanensis (Kruskal-Wallis ANOVA). Полужирным шрифтом выделено значимое влияние (p < 0.05)
| Год | Показатель | Значимость влияния фактора (p) | ||
|---|---|---|---|---|
| удобрение | соседи | укос | ||
| 2010 | Численность генеративных побегов | 0.3071 | 0.0788 | 0.9393 |
| Численность вегетативных побегов | 0.2692 | 0.8514 | 0.8731 | |
| Суммарная численность побегов | 0.2012 | 0.8551 | 0.4345 | |
| 2011 | Численность генеративных побегов | 0.6854 | 1.000 | <0.0001 |
| Численность вегетативных побегов | 0.7150 | 0.2733 | 0.2725 | |
| Суммарная численность побегов | 1.000 | 0.2709 | 0.9039 | |
| Биомасса 1-го укоса | 0.6547 | 0.1797 | 0.9048 | |
| Биомасса 2-го укоса | 0.2059 | 0.2059 | – | |
| 2012 | Численность генеративных побегов | 0.0673 | 0.0281 | 0.0002 |
| Численность вегетативных побегов | 0.7861 | 0.4067 | 0.8977 | |
| Суммарная численность побегов | 0.1441 | 0.4652 | 0.4966 | |
| Биомасса 1-го укоса | 1.000 | 0.0253 | 0.4066 | |
| Биомасса 2-го укоса | 0.5271 | 1.000 | – | |
| 2013 | Численность генеративных побегов | 0.187 | 0.187 | 0.0002 |
| Численность вегетативных побегов | 0.0446 | 0.2012 | 0.3557 | |
| Суммарная численность побегов | 0.0169 | 0.581 | 0.0377 | |
| Биомасса итоговая | 0.302 | 0.194 | <0.0001 | |
Рис. 1.
Численность генеративных побегов (а) Festuca ovina, Anemone speciosa, Oxytropis kubanensis и надземная биомасса (б) этих видов растений на экспериментальных площадках в 2013 г. по вариантам частоты скашивания. Показаны средние значения и ошибка среднего, n = 40.
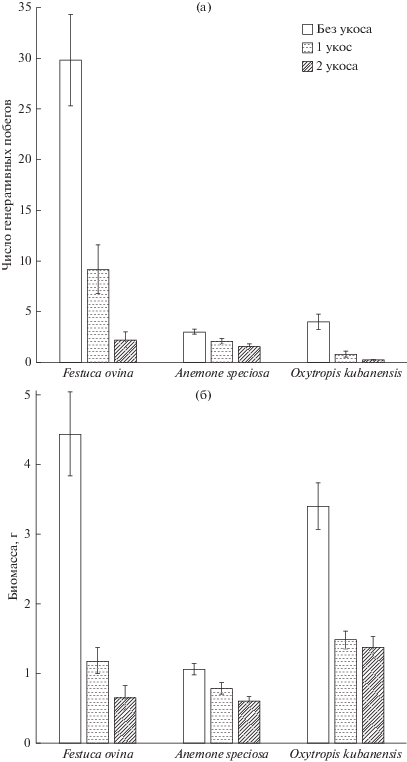
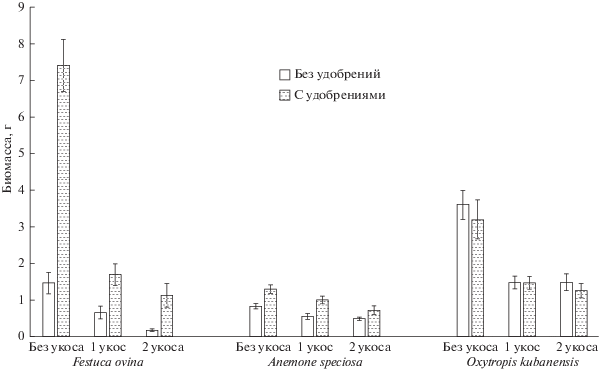
Фактор “удобрения” к концу эксперимента имел значимое положительное влияние на численность побегов и биомассу Festuca ovina (биомасса в 2013 г. в контроле 1.1 ± 0.4 г/площадку, в варианте только с внесением удобрений – 8.3 ± ± 1.1), численность генеративных побегов и биомассу Anemone speciosa, но отрицательное – на итоговую численность побегов Oxytropis kubanensis. Таким образом, обогащение почвы элементами минерального питания увеличивало надземную биомассу и улучшало ее восстановление во всех вариантах независимо от режима дефолиации у двух из трех исследованных видов (рис. 2).
Рис. 2.
Влияние удобрений на биомассу растений в различных вариантах кошения (итоговая надземная биомасса в 2013, г/площадку, среднее и его ошибка, n = 40).
В большинстве случаев фактор “удаление соседних растений” не оказал значимого влияния на восстановление численности и биомассы растений после дефолиации. Для Festuca ovina отмечено увеличение отрастания надземной массы после первого укоса при наличии соседних растений на второй год эксперимента, а также увеличение численности вегетативных побегов в присутствии соседей на третий год эксперимента (табл. 3). Таким образом, растения этого вида, скорее, испытывают положительное, чем конкурентное, влияние соседних растений. Аналогично для Oxytropis kubanensis на третий год эксперимента мы наблюдали положительное влияние соседних растений на численность генеративных побегов и биомассу первого укоса (табл. 4). Значимого влияния фитоценотического окружения на Anemone speciosa не отмечено (табл. 5). Необходимо подчеркнуть, что ни в одном случае, вопреки априорной гипотезе, мы не обнаружили улучшения роста растений при снижении конкуренции.
ОБСУЖДЕНИЕ
Нами исследована реакция трех типичных видов растений низкопродуктивных альпийских пустошей на ежегодное одно- и двукратное удаление надземной биомассы (дефолиацию). Для всех видов и большинства изученных параметров интенсивность дефолиации оказывала отрицательное воздействие на дальнейшее отрастание растений в текущем и последующем сезонах, что согласуется с результатами экспериментов как в высокогорных сообществах [28, 32–34], так и в травяных экосистемах равнин [35, 36]. Наиболее резко при дефолиации снижается образование генеративных побегов, что связано с их закладкой в годы, предшествующие цветению [19, 37], что также было отмечено во многих работах [32, 33, 38, 39].
Исследованные нами виды растений существенно различались по отавности при снижении конкурентных воздействий (удалении соседних растений) и внесении элементов минерального питания. Наименее отзывчивым на эти воздействия был представитель бобовых – Oxytropis kubanensis, на восстановление побегов которого присутствие соседних растений оказывало благоприятное влияние. Таким образом, для этого вида недостаток почвенных ресурсов не влиял на рост и накопление биомассы,что, видимо, связано с его способностью эффективно фиксировать азот клубеньковыми симбионтами [40].
Конкуренция в альпийских лишайниковых пустошах между доминирующими видами сосудистых растений хорошо выражена [25], в то время как доминанты обычно оказывают благоприятное воздействие на более редкие виды [24]. В нашем эксперименте мы практически не выявили отрицательного влияния соседних растений на биомассу изученных видов, в то время как для Festuca ovina отмечено положительное влияние со стороны соседей на биомассу растений при двукратном скашивании в 2011 г. и численность вегетативных побегов в 2012 г. Это свидетельствует о меньшем влиянии конкуренции на отавность растений в изученном сообществе, возможно, в связи с незначительным обогащением почвы при удалении соседей и медленным разложением их подземных органов [41, 42]. Исследованные виды высокогорий отличаются от многих других растений, у которых снижение конкуренции положительно сказывалось на восстановлении надземной биомассы и семенной продукции после дефолиации или повреждения фитофагами [13, 43–45].
Напротив, при прямом обогащении почвы ЭМП существенно увеличились как рост контрольных растений, так и отавность двух из трех изученных видов (Anemone speciosa, Festuca ovina). Эти данные подтверждают большую роль богатства почвы для образования биомассы растений в условиях альпийского пояса, что было отмечено многими исследователями и в других условиях: для древесных растений [46], вереска [13], злаков [47], тундровых растений [48] и т.п. У Poa bulbosa компенсационный рост после дефолиации наблюдался только при высоком уровне богатства почвы и отсутствовал при низком [49]. В то же время внесение NP снижало отавность Eriophorum vaginatum по отношению к неудобренным растениям, но улучшало для Hierochloe alpina в тундрах [50].
Интересно отметить, что для Festuca ovina показано значительное снижение корневой биомассы при дефолиации [51], и это хорошо объясняет лучшую отавность вида при обогащении почвы ЭМП. В ряде случаев дефолиация может вызвать увеличение численности побегов за счет пробуждения спящих почек на фоне снижения их общей массы [52], однако мы не наблюдали этого явления для изученных видов.
Таким образом, проведенные нами эксперименты впервые показали, что в условиях низкопродуктивных сообществ альпийских пустошей снятие конкуренции в отличие от внесения NP удобрений не улучшает восстановление биомассы после дефолиации. Изученные факторы (влияние соседних растений и обогащение почвы ресурсами) принципиально по-другому влияли на рост и отавность азотфиксирующего бобового растения: увеличение доступности почвенных ресурсов снижало численность его побегов и не оказывало влияния на прирост биомассы. Нами не отмечено случаев положительного влияния удаления соседних растений (частичное снятие конкуренции) на численность и биомассу изученных видов, напротив, в отдельных вариантах для двух из трех видов мы наблюдали большую численность или надземную биомассу в присутствии соседних растений. Обогащение почвы доступными формами азота и фосфора улучшало прирост небобовых растений независимо от режима дефолиации.
Работа выполнена при финансовой поддержке РНФ (проект № 19-14-00038).
Список литературы
Zamora R., Hodar J.A., Gomes J.M. Plant-herbivore interaction: beyond a binary vision // Handbook of functional plant ecology / Pugnaire F.I., Valladares F. Eds. New York, Basel: Marcel Dekker, 1999. P. 677–718.
Schowalter T.D. Insect ecology. An ecosystem approach. 2-nd ed. Amsterdam e.a.: Elsevier AP, 2006. 572 p.
Olofsson J., de Mazancourt C., Crawley M.J. Contrasting effects of rabbit exclusion on nutrient availability and primary production in grasslands at different time scales // Oecologia, 2007. V. 150. № 4. P. 582–589.
Абатуров Б.Д., Дмитриев И.А., Жаргалсфйхан Л., Омаров К.З. Утилизация фитомассы и отложение экскрементов копытными млекопитающими на степных пастбищах Восточной Монголии // Изв. РАН. Серия биологич. 2008. № 3. С. 350–359.
Agrawal A.A. Current trends in the evolutionary ecology of plant defense // Functional Ecology. 2011. V. 25. № 2. P. 420–432.
Работнов Т.А. Луговедение. Изд. 2-е. М.: Изд-во МГУ, 1984. 320 с.
Горчакова А.Ю. Об отавности злаков лесостепной зоны Среднего Поволжья // Экологич. вестн. Северного Кавказа. 2015. Т. 11. № 2. С. 62–74.
Grime J.P. Plant strategies, vegetation processes, and ecosystem properties. 2-nd ed. Chichester: John Wiley and Sons, 2001. 417 p.
Benot M.-L., Mony C., Puijalon S. et al. Responses of clonal architecture to experimental defoliation: a comparative study between ten grassland species // Plant Ecology. 2009. V. 201. № 2. P. 621–630.
Suding K.N., Goldberg D.E., Hartman K.M. Relationship among species traits: separating levels of response and and identifying linkages to abundance // Ecology. 2003. V. 84. № 1. P. 1–16.
De Bello F., Leps J., Sebastia M.-T. Predictive value of plant traits to grazing along a climatic gradient in the Mediterranean // J. of Applied Ecology. 2005. V. 42. № 5. P. 824–833.
Vesk P.A., Westoby M. Predicting plant species' responses to grazing // J. of Applied Ecology. 2001. V. 38. № 5. P. 897–909.
Sardans J., Penuelas J., Roda F. The effects of nutrient availability and removal of competing vegetation on resprouter capacity and nutrient accumulation in the shrub Erica multiflora // Acta Oecologica. 2006. V. 29. № 2. P. 221–232.
Fanselow N., Schönbach P., Gong X.Y. et al. Short-term regrowth responses of four steppe grassland species to grazing intensity, water and nutrient in Inner Mongolia // Plant and Soil. 2011. V. 340. № 1-2. P. 279–289.
Энеев С.Х. Этология яков в Кабардино-Балкарии // Труды Кубанского СХИ. 1985. Вып. 264. С. 50–54.
Онипченко В.Г., Ежелев З.С. Экологические последствия выпаса яков в Карачаево-Черкесии // Экологич. вестн. Северного Кавказа. 2010. Т. 6. № 1. С. 45–57.
Соколов В.Е., Темботов А.К. Млекопитающие: Копытные. М.: Наука, 1993. 528 с.
Onipchenko V.G. (ed.) Alpine ecosystems in the Northwest Caucasus. Dordrecht e.a.: Kluwer Academic Publishers, 2004. 407 p.
Körner C. Alpine plant life: functional plant ecology of high mountain ecosystems. 2-nd ed. Berlin e.a.: Springer, 2003. 349 p.
Olofsson J. Short- and long-term effects of changes in reindeer grazing pressure on tundra heath vegetation // J. of Ecology. 2006. V. 94. № 2. P. 431–440.
Gough L., Shrestha K., Johnson D.R., Moon B. Long-term mammalian herbivory and nutrient addition alter lichen community structure in Alaskan dry heath tundra // Arctic, Antarctic, and Alpine Research. 2008. V. 40. № 1. P. 65–73.
Онипченко В.Г. Структура, фитомасса и продуктивность альпийских лишайниковых пустошей // Бюл. МОИП. Отд. биол. 1985. Т. 90. № 1. С. 59–66.
Onipchenko V.G. The structure and dynamics of alpine plant communities in the Teberda Reserve, the Northwestern Caucasus // Oecologia Montana. 1994. V. 3. № 1. P. 40–50.
Aksenova A.A., Onipchenko V.G., Blinnikov M.S. Plant interaction in alpine tundra: 13 years of experimental removal of dominant species // Ecoscience. 1998. V. 5. № 2. P. 258–270.
Onipchenko V.G., Blinnikov M.S., Gerasimova M.A. et al. Experimental comparison of competition and facilitation in alpine communities varying in productivity // J. of Vegetation Science. 2009. V. 20. № 4. P. 718–727.
Онипченко В.Г., Дудова К.В., Ахметжанова А.А. и др. Какие стратегии растений способствуют их доминированию в альпийских сообществах? // Журн. общ. биол. 2020. Т. 81. № 1. С. 37–46.
Онипченко В.Г. Фитомасса альпийских сообществ Северо-Западного Кавказа // Бюл. МОИП. Отд. биол. 1990. Т. 95. № 6. С. 52–62.
Текеев Д.К., Онипченко В.Г. Реакция альпийских растений малоснежных местообитаний на экспериментальную дефолиацию // Экологич. вестн. Северного Кавказа. 2007. Т. 3. № 2. С. 5–13.
Soudzilovskaia N.A., Onipchenko V.G., Cornelissen J.H.C., Aerts R. Biomass production, N:P ratio and nutrient limitation in a Caucasian alpine tundra plant community // J. of Vegetation Science. 2005. V. 16. № 4. P. 399–406.
Soudzilovskaia N.A., Onipchenko V.G. Experimental investigation of fertilization and irrigation effects on an alpine heath, northwest Caucasus, Russia // Arctic, Antarctic, and Alpine Research. 2005. V. 37. № 4. P. 602–610.
Зернов А.С., Алексеев Ю.Е., Онипченко В.Г. Определитель сосудистых растений Карачаево-Черкесской Республики. М.: КМК, 2015. 459 с.
Wyka T. Carbohydrate storage and use in an alpine population of the perennial herb, Oxytropis sericea // Oecologia, 1999. V. 120. № 2. P. 198–208.
Аджиева Р.Б., Онипченко В.Г. Реакция растений альпийских ковров и гераниево-копеечниковых лугов на разные режимы дефолиации // Бюл. МОИП. Отд. биол. 2006. Т. 111. Вып. 1. С. 24–36.
Текеев Д.К., Семенова Р.Б., Онипченко В.Г. Интегральная оценка отавности альпийских растений // Вестник МГУ. Сер. 16. Биология. 2012. № 1. С. 36–40.
Green R.A., Detling J.A. Defoliation-induced enhancement of total aboveground nitrogen yield of grasses // Oikos. 2000. V. 91. № 2. P. 280–284.
Ferraro D.O., Oesterheld M. Effect of defoliation on grass growth. Aquantitative review // Oikos. 2002. V. 98. № 1. P. 125–133.
Нахуцришвили Г.Ш., Гамцемлидзе З.Г. Жизнь растений в экстремальных условиях высокогорий (на примере Центрального Кавказа). Л.: Наука, 1984. 123 с.
Mueggler W.F. Response of Mountain grassland vegetation to clipping in southwestern Montana // Ecology. 1967.V. 48. № 6. P. 946–949.
Brys R., Shefferson R.P., Jacquemyn H. Impact of herbivory on flowering behaviour and life history trade-offs in a polycarpic herb: a 10-year experiment // Oecologia. 2011. V. 166. № 2. P. 293–303.
Макаров М.И., Малышева Т.И., Ермак А.А. и др. Симбиотическая азотфиксация в сообществе альпийской лишайниковой пустоши Северо-Западного Кавказа (Тебердинский заповедник) // Почвоведение. 2011. № 12. С. 1504–1512.
Воронина И.Н., Онипченко В.Г., Игнатьева О.В. Компоненты биологического круговорота на альпийских лишайниковых пустошах Северо-Западного Кавказа // Почвоведение. 1986. № 1. С. 29–37.
Elumeeva T.G., Onipchenko V.G., Akhmetzhanova A.A. Stabilization versus decomposition in alpine ecosystems of the Northwestern Caucasus: the results of tea bag burial experiment // J. of Mountain Science. 2018. V. 15. № 8. P. 1633–1641.
Briske D.D. Strategies of plant survival in grazed systems: a functional interpretation // Foraging strategies of grazing animals / Hodgson J., Illius A.W. Eds. Wallingford: CAB International, 1996. P. 37–67.
del-Val E., Crawley M.J. Interspecific competition and tolerance to defoliation in four grassland species // Canadian J. of Botany. 2004. V. 82. № 7. P. 871–877.
Hartnett D.C., Fay P.A. Plant populations: patterns and processes // Grassland dynamics / Knapp A.K., Briggs J.M., Hartnett D.C., Collins S.L. Eds. New York, Oxford: Oxford University Press, 1998. P. 81–100.
Hanley M.E. Seedling herbivory, community composition and plant life history traits // Perspectives in Plant Ecology, Evolution and Systematics. 1998. V. 1. № 2. P. 191–205.
Kohyani P.T., Bossuyt B., Bonte D., Hoffmann M. Differential herbivory tolerance of dominant and subordinate plant species along gradients of nutrient availability and competition // Plant Ecology. 2009. V. 201. № 2. P. 611–619.
Olofsson J. Effects of simulated reindeer grazing, trampling, and waste products on nitrogen mineralization and primary production // Arctic, Antarctic, and Alpine Research. 2009. V. 41. № 3. P. 330–338.
van Staalduinen M.A., Dobarro I., Peco B. Interactive effects of clipping and nutrient availability on the compensatory growth of a grass species // Plant Ecology. 2010. V. 208. № 1. P. 55–64.
Johnson D.R., Gough L. Two arctic graminoids differ in tolerance to herbivory when grown with added soil nutrients // Botany. 2013. V. 91. № 2. P. 82–90.
Dawson L.A., Thornton B., Pratt S.M., Paterson E. Morphological and topological responses of roots to defoliation and nitrogen supply in Lolium perenne and Festuca ovina // New Phytologist. 2003. V. 161. № 3. P. 811–818.
Lee W.G., Fenner M., Loughnan A., Lloyd K.M. Long-term effects of defoliation: incomplete recovery of a New Zealand alpine tussock grass, Chionochloa pallens, after 20 years // J. of Applied Ecology. 2000. V. 37. № 2. P. 348–355.
Дополнительные материалы
- скачать ESM.docx
- Таблица 1е. Результаты трехфакторного дисперсионного анализа изменения надземной биомассы исследованных видов через 3 года после экспериментального воздействия (дефолиация, внесение удобрений, удаление соседних растений).
Таблица 2е. Средняя биомасса и численность побегов растений после 3 лет экспериментального воздействия (2013 г.) во всех вариантах эксперимента (приведены значения среднего и его ошибки, во всех случаях N=10)