

Журнал физической химии, 2019, T. 93, № 11, стр. 1746-1755
Доказательства отсутствия транс–цис-фотоизомеризации ридимеров метилоранжа в водной среде и в комплексах включения
Ю. А. Михеев a, *, Ю. А. Ершов b
a Российская академия наук, Институт биохимической физики им. Н.М. Эмануэля
Москва, Россия
b Московский государственный технический университет им. Н.Э. Баумана
Москва, Россия
* E-mail: mik@sky.chph.ras.ru
Поступила в редакцию 17.12.2018
После доработки 17.12.2018
Принята к публикации 15.01.2019
Аннотация
Проведен анализ данных по лазерному ультравысокоскоростному зондированию фотоиндуцированных состояний метилоранжа в водной среде и в комплексах с циклодекстринами, а также транс-аминоазобензола в этаноле, полученных методами ультрабыстрых транзитных линз (UTL) и транзитной абсорбционной спектроскопии (TAS). В основу анализа положена концепция ридимерного строения азокрасителей в их основном состоянии, причем аминоазобензол рассматривается как референтное соединение. Показано, что несоответствие, существующее между UTL-, TAS-сигналами ридимеров метилоранжа и UTL-, TAS-сигналами ридимеров аминоазобензола, обусловлено неспособностью ридимеров метилоранжа к транс–цис-фотоизомеризации в водной среде и в комплексах включения, протекающей вследствие диссоциации ридимеров аминоазобензола в этаноле на мономеры. Один из мономеров, обладающий двумя катионами фениламинильного типа и VIS-поглощением в интервале длин волн 500–750 нм, превращается в цис-изомер. В отличие от этого фотодиссоциация ридимеров метилоранжа на мономеры не приводит к образованию цис-изомеров. Причины такого различия – иммобилизация и ограниченная подвижность ридимеров метилоранжа в аквакапсулах и нанополостях циклодекстринов.
В работах [1–5] установлено, что простые красители на основе аминоазобензола в основном состоянии являются ридберговскими димерами (ридимерами). Межмономерная связь в ридимерах – ковалентная связь нового типа – образуется в результате спаривания электронов, промотированных с sp2-орбиталей атомов N азогрупп на ридберговские 3s-орбитали азогрупп. При этом характерная для азокрасителей цветность связана не с их хиноидными мономерами, как полагали до этого более 100 лет, а с принадлежащими ридимерам катионами фениламинильного типа (CPhAT).
На основе работ [1–5] дана новая трактовка экспериментальных данных по фотохимии аминоазобензольных красителей. Так, в [6] объяснены резонансные рамановские спектры ридимеров аминоазобензола (AAB2), а в [7, 8] раскрыта природа транзитных состояний AAB2, установленных в [9, 10] методами UTL и TAS, и раскрыт механизм транс–цис-фотоизомеризации.
В настоящей работе новая трактовка UTL- и TAS-сигналов AAB2 [7, 8] положена в основу анализа UTL- и TAS-сигналов метилоранжа (MOD), приведенных в работе [11]. Показано, что эти сигналы тоже обусловлены ридимерами MOD2, но не традиционно постулируемыми мономерами строения
где R = CH3, X = ${\text{SO}}_{3}^{ - }$.
Результаты проведенного анализа позволяют также сделать заключение о неспособности находящихся в воде ридимеров MOD2 вступать в транс–цис-изомеризацию, в отличие от AAB2 в этаноле.
УЛЬТРАБЫСТРЫЕ UTL- И TAS-МЕТОДЫ
Авторы [9–11] использовали ультрабыстрые UTL- и TAS-методы с целью уточнения механизма фотопроцессов транс–цис-изомеризации t‑AAB (в этаноле и гептаноле) и MOD (в водной среде и в комплексах с циклодекстринами). При этом они традиционно исходили из представления о мономерном строении аминоазобензольных красителей. Между тем, транзитные VIS-спектры и UTL-сигналы от MOD и AAB имеют отличия, объяснимые только в рамках ридимерной концепции.
Метод ультрабыстрой транзитной абсорбционной спектроскопии (TAS)
Для регистрации TAS-спектров MOD2 и AAB2 использовали импульсную лазерную установку, генерирующую излучение с длиной волны λ = 800 нм и частотой повторения импульсов 1000 Гц [9–11]. Это излучение расщепляли на два пучка, один из которых после преобразования с удвоением частоты в видимый (VIS) свет (λex = 400 нм) использовали в качестве пучка фотовозбуждения (pump beam) ридимеров (мощность этого пучка составляла менее 200 мкДж). Другой пучок после преобразования в континиум белого света использовали в качестве зондового (probe) пучка, ориентированного под малым углом к пучку возбуждения и фокусированного в ту же точку в кювете с образцом, что и пучок возбуждения. Импульсы зондового пучка генерировали через определенное время задержки, причем малую его часть отделяли в канал сравнения. Облучаемые растворы красителей (концентрация 0.3 ммоль/л) пропускали через проточную кварцевую кювету с оптическим путем 0.5 мм.
Авторы [11] предполагали, что в воде фотовозбужденные молекулы MOD претерпевают транс–цис-изомеризацию, наличие которой установили для AAB2 [9, 10] (механизм фотоизомеризации AAB2 дан в [7, 8]). В этой связи они поставили задачу исследовать характер изменения транзитных сигналов комплексов предполагаемых молекул MOD с циклодекстринами (CD) в стерических затрудненных условиях для транс–цис-изомеризации.
TAS-сигналы MOD2 в воде и комплексах включения
Для получения комплексов включения использовали α-, β- и γ-циклодекстрины, молекулы которых построены из шести, семи и восьми глюкопиранозных единиц соответственно [11, 12]. Эти молекулы имеют форму усеченных конусов высотой 7.8 Å с внешней гидрофильной поверхностью и отличаются размерами внутримолекулярных полостей, имеющих гидрофобную поверхность и диаметры у основания соответственно 5.7, 7.8 и 9.5 Å [12]. Желая получить комплексы в соотношении CD/MOD-молекулы = 1 : 1 и 2 : 1, авторы [11] использовали CD-концентрации 0.3 и 10 ммоль/л соответственно. По их мнению, высокая концентрация CD обеспечивает образование преимущественно комплексов 2 : 1, в которых молекулы MOD будут закапсулированы с обоих концов и получат наиболее сильное стеснение. В [11] не сообщено о спектрах TAS для комплексов с α- и β-CD. В то же время было обнаружено изменение TAS-спектров у комплексов MOD2 с γ‑CD относительно спектров MOD2 в водном растворе без γ-CD. Наблюдавшиеся в [11] для нулевого времени задержки спектры TAS приведены на рис. 1а.
Рис. 1.
Сигналы TAS метилового оранжевого красителя в воде(а) и аминоазобензола в этаноле (б) при концентрациях 0.3 ммоль/л в присутствии γ-CD концентрацией: 1 – 0, 2 – 0.3, 3 – 10 ммоль/л (а), и без γ-CD (б). Время задержки зондового пучка, пс: 1 – 0.1, 2 – 0.5, 3 – 1.0, 4 – 5.0 (б). По данным [9–11].

На рис. 1а кривая 1 характеризует TAS-спектр MOD2 в воде без γ-CD, кривая 2 – спектр совместного раствора MOD2 с γ-CD при концентрациях 0.3 ммоль/л, кривая 3 – спектр раствора с содержанием 0.3 ммоль/л MOD2 и 10 ммоль/л γ-CD. Авторы [11] ограничились данными рис. 1а, уделив основное внимание UTL-исследованиям (данные UTL рассмотрены ниже).
При описании TAS-спектров комплексов включения (рис. 1a, спектры 2, 3) принято [11], что в относительно крупные полости γ-CD инклюдируются две молекулы MOD, претерпевая агрегацию с параллельной укладкой. При этом в комплексах “1 : 2” димеры MOD-молекул фиксируются с одного конца (рис. 2б). Этим комплексам приписаны транзитный спектр 2 (рис. 1а) и низкая скорость транс–цис-изомеризации [11]. В комплексах “2 : 2” (рис. 2в), согласно [11], димеры MOD капсулируются с двух концов. Этим комплексам приписаны спектр 3 (рис. 1а) и практическое отсутствие транс–цис-изомеризации.
Рис. 2.
Сигналы UTL метилоранжа (а) в водной среде при концентрации 0.3 ммоль/л и различной концентрации γ-CD: 1 – 0, 2 – 0.3, 3 – 10 ммоль/л; б и в – модели комплексов включения в работе [11].

Следует отметить, что, в соответствии с новыми фактами [1–8], основным состоянием метилоранжа, как и других производных аминоазобензола, являются ридимеры. Далее будет показано, что спектры 2 и 3 на рис. 1а соответствуют транзитным фотовозбужденным состояниям, возникающим при наличии двух ридимеров MOD2, включенных в полость γ-CD.
Что касается ридимеров AAB2, то их TAS-сигналы, полученные для этанольного раствора в кинетическом (time-resolved) варианте в работах [9, 10], приведены на рис. 1б. Они получили свое объяснение в рамках ридимерной концепции в [7, 8] и будут использованы как референтные спектры при обсуждении данных по MOD2.
Метод ультрабыстрых транзитных линз (UTL)
Метод UTL служит для определения динамики молекул растворителя и растворенного соединения по транзитному неоднородному изменению показателя преломления (n) среды. Такое транзитное изменение n относительно n0 возникает в условиях фотовозбуждения растворенных молекул сфокусированным пучком света в результате безызлучательной конверсии энергии возбуждения. В [10, 11] использовали лазерную установку, генерирующую импульсы возбуждения и импульсы зондирования с одинаковой поляризацией электрического вектора. Импульсы зондирования (probe), имеющие λp = 800 нм и частоту повторения 76 МГц, направляли через линию оптической задержки коллинеарно с импульсами возбуждения (λex = 400 нм, частота повторения 1.1 МГц, мощность менее 200 пДж/импульс) и фокусировали оба пучка излучения на ячейку с образцом выпуклой линзой с f = 50 мм. Определяли изменение интенсивности излучения в центральном сечении зондового луча. Центральное сечение выделяли, пропуская зондовый луч через расположенный на удалении от образца фильтр, снабженный “булавочным отверстием” (pinhole).
UTL-сигналы от ридимеров MOD2
Импульсное возбуждение MOD2 [11] и AAB2 [10] вызывает мгновенный рост интенсивности зондового излучения вследствие мгновенного роста показателя преломления, связанного с оптическим эффектом Керра (OKE, nOKE – n0 > 0). Эффект OKE имеет место и в чистых растворителях вследствие мгновенного отклика электронов среды на световое воздействие и появления транзитной линзы с положительным фокусом. Спад интенсивности OKE-сигнала наступает практически при нулевом времени задержки зондового пучка. На его спадающую ветвь налагается индуцированное фотовозбуждением ослабление интенсивности зондового импульса за счет появления дефокусирующей транзитной UTL-линзы с отрицательным фокусом. Появление такой линзы связывают с безызлучательной конверсией энергии возбуждения в колебательно возбужденное основное электронное состояние с последующей передачей конвертированной энергии молекулам среды, в результате чего происходит снижение показателя преломления (n0 – n > 0).
На рис. 2а приведены транзитные UTL-сигналы [11] для водных растворов MOD2 (кривая 1), а также для комплексов включения MOD2 в γ-CD при концентрациях 0.3 ммоль/л обоих компонентов (кривая 2) и при концентрациях 0.3 ммоль/л у MOD2 и 10 ммоль/л у γ-CD (кривая 3). Расчетный анализ кривой 1, проведенный с разделением сигналов OKE и UTL [11], показал, что наблюдаемый вслед за OKE спад сигнала UTL соответствует сумме двух экспоненциальных кривых [10, табл. 1 ]. Одна из этих кривых имеет характеристическое время τ1 = 0.6–0.9 пс, другая –τ2 = 8–13 пс. Время τ1 приписали конверсии энергии исходного возбужденного состояния (трактуемого как S2) молекулы t-MOD в энергию колебательно-возбужденного состояния (S2 ~→ $S_{0}^{v}$), а τ2 – колебательной релаксации $S_{0}^{v}$ ~→ S0.
Сходные компоненты UTL-сигналов с такими же значениями констант τ1, τ2 были установлены для комплексов включения в растворах с концентрацией 0.3 ммоль/л t-MOD (в расчете на молекулы), содержащих 0.3 ммоль/л α-CD, β-CD и 10 ммоль/л β-CD. Всем этим комплексам был приписан состав 1 : 1.
Для UTL-сигналов от комплексов предполагаемых молекул t-MOD с γ-CD при концентрациях 0.3 ммоль/л t-MOD и 0.3 ммоль/л γ-CD авторы получили примерно такие же кинетические константы (τ1 = 0.6–1.2 пс, τ2 = 6–12 пс). Однако, учитывая, что при указанных молярных концентрациях константа комплексообразования в равновесии “t-MOD + γ-CD ↔ (t-MOD, γ-CD)” на четыре порядка больше, чем при таких же концентрациях t-MOD с α- и β-CD, авторы [11] приняли, что комплексы с γ-CD включают две молекулы t-MOD в виде димеров (1 : 2), капсулируемых с одного конца (рис. 2б). Для концентраций 0.3 ммоль/л t-MOD и 10 ммоль/л γ-CD спад UTL-сигналов был более длительным, с τ1 = 1.1–2.1 пс, τ2 = 13–17 пс. Это было объяснено капсулированием димеров t-MOD с двух концов (2 : 2) (рис. 2в).
Следует отметить, что UTL-сигнал находящихся в воде предполагаемых свободных молекул MOD снижается в область отрицательных значений при ~2.5 пс (рис. 2, кривая 1), после чего очень медленно возвращается к нулю. Это свойство характерно также для комплексов с α-CD, β-CD (1 : 1), и авторы связали его с образованием цис-формы MOD, предположив для нее более низкий показатель преломления относительно транс-формы MOD.
Транзитные UTL- и TAS-сигналы ридимеров AAB2
Ридимеры AAB2 в среде этанола и гептанола исследовали с помощью описанных выше установок [9, 10]. При этом оказалось, что полученная для AAB2 картина отличается от картины с MOD2 числом экспоненциальных компонент в UTL-, TAS-сигналах и значениями кинетических констант.
Расчетное разделение наблюдавшихся OKE- и UTL-сигналов показало [10], что для спиртовых растворов AAB2 характерно наличие трех кинетических компонент в UTL-сигнале. Соответствующие им константы 0.3, 0.7 и 2 пс были вычислены с учетом эффекта снижения интенсивности сигнала UTL в области отрицательных значений на длительное время. Такое снижение, как и в случае с MOD2, было приписано образованию цис-формы молекул AAB.
Что касается TAS-сигналов AAB2 в работах [9, 10], то они практически одинаковы для обоих спиртов. На рис. 1б приведены транзитные TAS-спектры для растворов в этаноле, полученные в [9] при временах задержки импульсов зондирующих лучей 0.1, 0.5, 1.0 и 5.0 пс. Здесь спектр 1 имеет широкую полосу TAS в области 410–750 нм и относительно долгоживущую полосу отбеливания (bleachihg) в области поглощения AAB2 при 370–410 нм (ее возвратная эволюция соответствует моноэкспоненциальной функции с характеристическим временем τ = 15 пс).
Спад интенсивности TAS в интервале λ = 600–725 нм описывается тремя экспонентами [9]. Наиболее быстрая компонента TAS с λmax = = 625 нм [9, рис. 4] и τ1 = 0.2 пс фактически соответствует компоненте UTL с τ1 = 0.3 пс [10]. Эта компонента в области 500–600 нм практически не определяется. Она отсутствует и в TAS-спектрах MOD2 работы [11] (рис. 1а, кривые 1–3). Относящиеся к AAB2 две другие TAS-компоненты охватывают одну и ту же спектральную область 410–750 нм и имеют один общий максимум 525 нм [9, рис. 4] (рис. 1б). Они спадают медленнее, характеризуются константами τ2 = 0.6 пс и τ3 = 1.9 пс и, несмотря на почти трехкратное различие констант, их спектры TAS имеют одинаковую форму, не меняющуюся в ходе их исчезновения. В конце TAS-эволюции наблюдается спектр раствора, обедненного AAB2 и обогащенного цис-формой AAB. Регенерация t-AAB достигает ~75%, тогда как ~25% превращается в c-AAB [9].
Что касается быстрой транзитной TAS-компоненты AAB2 (0.2 пс в области спектра 600–750 нм), то авторы [9] приписывают ее процессу конверсии энергии возбужденных молекул с S2-уровня (π,π*-состояние) на S1-уровень (“n,π*-состояние”). Наличие двух компонент спада, идущих в спектральной области 410–750 нм с τ2 = = 0.6 пс и τ3 = 1.9 пс и имеющих одинаковую форму спектров, они связывают с существованием двух разных путей спада “n,π*-состояния“ в основное S0-состояние молекул t-AAB.
Очевидно, что описание механизма конверсии энергии возбуждения с S2-уровня, данное в [9] для мнимых молекул t-AAB (а реально ридимеров AAB2 [1–8]), не согласуется с числом компонент в UTL-сигналах [11] предполагаемых молекул MOD (а реально ридимеров MOD2 [2]) и способом объяснения их природы.
Фотопревращения ридимеров AAB2 и MOD2
Процессу фотоизомеризации AAB предшествует диссоциация AAB2 на мономеры [7, 8]. Этого следует ожидать и для MOD2 как естественного свойства фотовозбужденных ридимеров. Рассмотрим в краткой форме механизм фотолиза AAB2 [7, 8], принимаемый в качестве референтного образца.
Фотоизомеризация ридимеров AAB2
Для фотопревращения аминоазокрасителей наиболее значимы e-конфигурации [7, 8]:
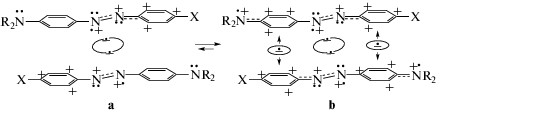
В схеме 1 символы R, X обозначают H в AAB2, точки под атомами N обозначают электроны на sp2-орбиталях, а точки над N – электроны на pz‑орбиталях. Между азогруппами мономеров показаны ридберговские ковалентные связи из двух электронов, промотированных с sp2-орбиталей азогрупп на R3s-орбитали (R3s–R3s-связи). В каждом мономере sp2-орбиталь, потерявшая электрон, наделяет свой атом N положительным зарядом. На pz-орбиталь этого атома N стягивается электрон с pz-орбитали соседнего атома N. Данная ситуация имеет место в каждом мономере ридимера. Потерявшие sp2-электроны атомы N мономеров индуцируют положительные заряды на pz-орбиталях соседних атомов N, которые поляризуют электронные pz-системы в сопряженных кольцах с образованием катионов фениламинильного типа (CphAT).
Ридимеры (1b) в схеме 1 образуются вследствие того, что атомы азота аминных групп отдают по электрону в соседние фениленовые кольца, которые становятся донорами электронов для CPhAT (Ph+N+) в противолежащих мономерах. В итоге связь между мономерами в ридимере усиливается, и возникают наиболее устойчивые e-таутомеры (1b). В них между противолежащими катионами обоих мономеров существуют одноэлектронные связи [3–8], фиксирующие четыре катиона фениламинильного типа с интенсивной полосой поглощения. В AAB2 такая полоса захватывает область 330–420 нм (λmax ~ 382 нм, растворитель ацетонитрил [6]). У растворенных в воде ридимеров MOD2, содержащих группы N(СH3)2 и ${\text{SO}}_{3}^{ - }$ в фенильных кольцах, такая полоса смещена в область 360–525 нм, λmax ~ 463 нм [2].
Из изложенного выше следует, что импульсное лазерное излучение с λex = 400 нм [9–11] должно взаимодействовать преимущественно с e‑таутомером (1b), создавая франк-кондоновские (FK) состояния (полное описание в [7, 8]):

здесь R = H. Схема 2 отражает процесс расщепления ридимера (2b) с образованием неодинаковых FK-пар мономеров (2сFK) и (2dFK) примерно в равных количествах. При этом гомогенная пара (2сFK) является результатом того, что разрыв R3s–R3s-связи фотовозбужденного ридимера (2b) приводит к возвращению освободившихся электронов на sp2-орбитали атомов N обоих мономеров. Эта пара является промежуточным FK-состоянием, ее последующее превращение рассмотрено ниже.
Образование гетерогенной пары (2d) связано с тем, что только одна $R_{{{\text{3}}s}}^{{\underset{\raise0.3em\hbox{$\smash{\scriptscriptstyle-}$}}{ \bullet } }}$-орбиталь разорванной при фотолизе R3s–R3s-связи возвращает электрон на sp2-орбиталь атома N. Вторая одноэлектронная $R_{{{\text{3s}}}}^{{\underset{\raise0.3em\hbox{$\smash{\scriptscriptstyle-}$}}{ \bullet } }}$-орбиталь захватывает электрон из одноэлектронной связи ридимера (2b). В итоге появляется пара (2dFK), состоящая из мономера t-AAB и поляризованного мономера с двумя катионами фениламинильного типа и с сильно ослабленной связью ${\text{N}}_{{ \bullet + }}^{{ + \bullet }}{\kern 1pt} - {\kern 1pt} {\text{N}}_{{ \bullet + }}^{{ \bullet + }}$ в азогруппе. Заряды поляризованного мономера уравновешиваются двумя отрицательными зарядами электронов, находящимися на ридберговской орбитали $R_{{{\text{3s}}}}^{{\underset{\raise0.3em\hbox{$\smash{\scriptscriptstyle-}$}}{ \bullet } {\kern 1pt} \underset{\raise0.3em\hbox{$\smash{\scriptscriptstyle-}$}}{ \bullet } }}$.
Вступать в транс–цис-изомеризацию способны только поляризованные мономеры из пары (2dFK), но не электронейтральные мономеры t‑AAB (в парах 2сFK и 2dFK) с прочной двойной связью N=N. Поляризованные мономеры содержат группу ${\text{N}}_{{ \bullet + }}^{{ + \bullet }}{\kern 1pt} - {\kern 1pt} {\text{N}}_{{ \bullet + }}^{{ \bullet + }}$ и ослабленную азосвязь в сочетании с сильным кулоновским отталкиванием положительно заряженных атомов азота. Именно их изомеризацией объясняют наличие субпикосекундных сигналов TAS (с τ1 = 0.2 пс [7]) и UTL (с τ1 = 0.3 пс [8]) и тот факт [7, 8], что доля образующихся молекул цис-изомеров (c-AAB) составляет, согласно [9, 10], всего ~25% от числа всех мономеров, входящих в AAB2. Остальные 75% мономеров рекомбинируют в AAB2 [7, 8].
Превращение FK-гомогенных пар AAB2
FK-пары из неполяризованных мономеров (2сFK) превращаются в FK-ридимеры с транзитной полосой при λmax = 525 нм по схеме [7, 8]:

(X, R = H у ААВ2). В схеме 3 каждый мономер в сFK-паре отдает на создание межмономерной связи по электрону, промотированному на ридберговскую орбиталь. Образующиеся при этом FK-ридимеры ${\text{a}}_{{\mathbf{r}}}^{{\mathbf{v}}}{\text{ и b}}_{{\mathbf{r}}}^{{\mathbf{v}}}$ несут на себе бинарные резонаторы из катионов ${\text{XP}}{{\hbar }^{ + }}{\text{N}}_{{ \bullet \bullet }}^{ + }$ (${\text{3a}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) и ${{{\text{R}}}_{{\text{2}}}}{{{\text{N}}}^{{ \bullet \bullet }}}{\text{P}}{{\hbar }^{ + }}{\text{N}}_{{ \bullet \bullet }}^{ + }$ (${\text{3b}}_{{\mathbf{r}}}^{{\mathbf{v}}}$ ).
В каждой паре таких резонаторов происходит расщепление π*-уровней одиночных резонаторов (механизм Симпсона), и это приводит к появлению суммарной широкой транзитной полосы VIS-поглощения в области 500–750 нм с λmax ~ ~ 525 нм (рис. 1б, кривые 2, 3) [7, 8]. В момент своего образования ридимеры (${\text{3a}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) ↔ (${\text{3b}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) имеют некоторый избыток колебательной энергии. Сначала этот избыток достаточно высок (“горячее колебательное состояние”), и стадию исчезновения TAS-сигнала с константой τ2 = 0.6 пс “горячих” структур (${\text{3a}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) ↔ (${\text{3b}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) связывают [7, 8] с их переходом в основное электронное, но термически неравновесное (1аT) состояние 1а в схеме 1. Такому переходу соответствует UTL-сигнал с τ2 = 0.7 пс [10].
Снижение степени колебательного возбуждения релаксирующих структур (${\text{3a}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) ↔ (${\text{3b}}_{{\mathbf{r}}}^{{\mathbf{v}}}$) приводит к более медленной стадии (в TAS τ3 = 1.9 пс [7], в UTL τ3 = 2.0 пс) их перехода в основное неравновесное электронное состояние (1аT). После этого происходит релаксация структур типа (1аТ) в состояние равновесия (1аT → 1а → 1b) с восстановлением исходных ридимеров AAB2 (схема 1).
Сходные стадии фотопревращений MOD2 и AAB2
Из изложенного выше следует, что в актах фотовозбуждения MOD2 должны участвовать наиболее устойчивые ридимеры типа (2b), причем их возбужденные e-конфигурации должны завершиться расщеплением как в случае AAB2 [7, 8]:

В схеме 4 FK-состояние – гомогенная пара (4сFK). В ней не приведены FK-гетерогенные пары типа (2dFK), так как среди TAS-сигналов MOD2 [11] в интервале длин волн 550–650 нм нет полосы поглощения с λmax = 625 нм (рис. 1а, кривые 1–3), характерной для поляризованных мономеров с ослабленной связью ${\text{N}}_{{ \bullet + }}^{{ + \bullet }}{\kern 1pt} - {\kern 1pt} {\text{N}}_{{ \bullet + }}^{{ \bullet + }}$ (рис. 1б, кривая 1). В суммарных UTL-сигналах MOD2 [11] тоже отсутствует высокоскоростная компонента, которая могла бы свидетельствовать в пользу транс–цис-изомеризации.
Вместе с тем, TAS-сигналы MOD2 (рис. 1а, кривые 1–3), содержат транзитную полосу с λmax = 535 нм, которая почти дублирует транзитные сигналы ридимеров AAB2 (рис. 1б, спектры 2, 3). На этом основании естественно предполагать, что транзитные ридимеры MOD2 должны появляться в соответствии со схемой 4. Следует также отметить, что TAS-сигнал 1 на рис. 1а включает в себя не только транзитную VIS-полосу с λmax = = 535 нм, как у AAB2 (рис. 1б, спектры 2, 3), но одновременно и полосу отбеливания (bleaching) исходного ридимера (область 400–500 нм) в соответствии со схемой 4, общей для обоих красителей. Таким образом, имеется определенное качественное сходство превращений обоих красителей, за исключением факта отсутствия транс–цис-изомеризации у MOD2.
Особенности фотопревращений MOD2
Сопоставление данных [9, 10] и [11] показало, что находящийся в водной среде краситель MOD2 не обладает UTL- и TAS-сигналами поляризованных мономеров с ослабленной азогруппой ${\text{N}}_{{ \bullet + }}^{{ + \bullet }}{\kern 1pt} - {\kern 1pt} {\text{N}}_{{ \bullet + }}^{{ \bullet + }}$, необходимой для транс–цис-изомеризации [7, 8]. При этом практическое совпадение кинетических констант UTL-сигналов (τ1 = 0.6–0.9 пс и τ2 = 8–13 пс) от ридимеров MOD2, находящихся в воде без CD и в комплексах включения с α-CD и β-CD (CD/MOD = 1 : 1 [11]), свидетельствует о том, что в этих системах действуют механизмы расщепления ридимеров MOD2 по схеме 4 и механизмы регенерации исходных ридимеров по схеме 3.
Отметим, что в экспериментах [11] соотношение исходных компонентов не соответствует молекулярным комплексам состава 1 : 1, так как ридимерная концентрация в 2 раза меньше молекулярной. В связи с этим возникает неопределенность: реально в растворе могут находиться как комплексы CD/MOD2 = 1 : 1, так и MOD2 вне CD. Вместе с тем, отмеченный выше факт совпадения кинетических констант UTL-сигналов свидетельствует о том, что ридимеры MOD2 в воде испытывают примерно такую же стерическую стесненность, как и в комплексах с α-CD и β-CD. В то же время установленные в [11] значительные отличия UTL- и TAS-сигналов у комплексов MOD2 с γ-CD свидетельствуют о наличии в этих комплексах структурных особенностей, которые были отмечены [11], но не получили адекватного объяснения.
Причины отсутствия транс–цис-изомеризации у MOD2
Как изложено выше, степень стесненности MOD2 в водной фазе примерно соответствует таковой в нанополостях α- и β-CD. Это можно объяснить гидрофобностью ридимеров. В противном случае не было бы движущих сил для образования комплексов включения. Определенная степень гидрофобности придает ридимерам MOD2 способность не только инклюдироваться в гидрофобные полости циклодекстринов [11], но и образовывать кластеры в нейтральной воде [2]. Дело в том, что растворение (в той или иной степени) гидрофобных молекул (в данном случае MOD2) в воде идет с формированием вокруг них супрамолекулярных аквакапсул, прочность которых превышает прочность сетки из водородных связей [13, 14]. При этом в MOD2 электроны азогрупп, находящиеся на 3s-ридберговских орбиталях, не только образуют ридберговские связи R3s–R3s, но также имеют некоторую вероятность пребывания в зоне третьих пучностей 3s-орбиталей азогрупп, выходя за пределы молекулярного каркаса. Они не только компенсируют положительные заряды ридимеров, но также способны взаимодействовать с молекулами воды, увеличивая прочность аквакапсулы. При этом действующее между аквакапсулой и иммобилизованным ридимером давление вызывает уменьшение ван-дер-ваальсовского размера и повышает его потенциал Гиббса.
Согласно [11], кинетические константы релаксации UTL-сигналов, генерируемых ридимерами MOD2 в аквакапсулах и комплексах α- и β-CD (1 : 1, молекулы), практически одинаковы. Они отражают условия иммобилизации и стерического стеснения, исключающие динамику фенильных колец, необходимую для транс–цис-изомеризации. Вследствие этого FK-пара (${\text{2d}}_{{{\text{FK}}}}^{{}}$) способна превратиться только в FK-пару (2cFK) за счет рекомбинации обоих электронов орбитали $R_{{{\text{3s}}}}^{{ \bullet \bullet }}$ с положительными зарядами обеих sp2-орбиталей азогруппы. Одновременно исчезают оба CPhAT:
Следует также отметить, что схема 4 образования фотовозбужденного FK-состояния MOD2 исключает рассмотренный ранее [4] вариант с хиноидной структурой фотовозбужденного MOD2.
Структура и фотопревращения комплексов γ-CD + MOD2
Для понимания механизма фотопревращения комплексов γ-CD + MOD2 важно, что их сигналы TAS (рис. 1а, кривые 2, 3) не содержат полосы отбеливания (bleaching), характерной для сигнала TAS ридимеров MOD2 в воде (рис. 1а, кривая 1). Сама полоса отбеливания является результатом вычитания спектра раствора MOD2, зафиксированного зондирующим пучком перед фотовозбуждением, из транзитного спектра, полученного после фотовозбуждения. И ее наличие у MOD2 в воде свидетельствует о том, что фотовозбуждение ридимеров MOD2 создает мономерные FK-пары, обладающие слабым поглощением VIS-света в VIS-полосе ридимеров (схемы 3, 4).
Отсутствие полосы отбеливания у комплексов γ-CD + MOD2 (рис. 1а, кривые 2, 3) можно объяснить взаимным наложением двух явлений, связанных с фотовозбуждением комплексов. Так, разностные спектры зондирующих пучков отражают: 1) образование FK-пар, слабо поглощающих свет в VIS-полосе ридимера b, (схемы 3, 4); 2) появление другого ридимера MOD2, имевшего в γ-CD до возбуждения измененную структуру, не поглощающую свет с λex = 400 нм. Другими словами, следует учитывать факт образования комплексов γ-CD + 2MOD2 и 2 γ-CD + 2MOD2. Это можно ожидать в связи с тем, что константы связи для комплексов с γ-CD (lg K = 7.26 в расчете на молекулы) значительно превосходят таковые для α-CD (lg K = 3.95) и β-CD (lg K = 3.3) [11].
Следует также отметить, что ван-дер-ваальсовские расстояния между соседними молекулами в кристаллах ароматических соединений равны 3.4 Å, тогда как для образования химических связей между мономерами в топохимических реакциях необходимо, чтобы расстояние (d) между ними стало меньше 3 Å [15]. Между тем, ридимеры (1b) уже имеют R3s–R3s-связь между азогруппами и две одноэлектронных связи между катионами CphAT. Эти связи обеспечивают высокую стабильность ридимеров даже в парообразном состоянии [3], что свидетельствует о наличии в них d < 3 Å и о возможности инклюдирования двух ридимеров в γ-CD.
Вместе с тем для объяснения TAS-сигналов на рис. 1а (кривые 2, 3) необходимо принять, что инклюдирование двух ридимеров MOD2 в γ-CD приводит к изменению e-конфигурации одного или обоих ридимеров. Так, в комплексах γ-CD + + 2MOD2 (преобладающих при концентрации [γ‑CD] = 0.3 ммоль/л) один из ридимеров, устойчивый в обычном равновесии (1b, схема 1), превращается в менее устойчивый ридимер (1а). Это сочетание представлено (со схематическим очертанием стенок полости) в схеме 5 структурой (5ab):
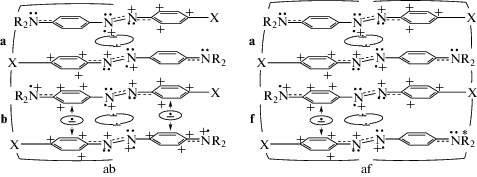
При концентрации [γ-CD] = 10 ммоль/л пара ридимеров капсулируется с обоих концов (комплексы 2γ-CD + ${\text{2MO}}{{{\text{D}}}_{2}}$, схема 5, af). В этом случае стесненность усиливается, и к образовавшейся структуре (а) присоединяется конформация f, вызванная деформированием H3С–N-связей группы N(CH3)2. Эти связи (соответствующая группа NR2 обозначена звездочкой) стремятся принять пирамидальную форму, характерную для электронной конфигурации азота 1s22s22p3. Данная ситуация ведет к потери sp2-гибридизации атома N и его сопряжения с фениленовым кольцом. Одновременно исчезает катион CphAT, и на аминный атом N возвращается электрон из одноэлектронной связи.
Представленные на схеме 5 сочетания ридимеров MOD2 необходимы и достаточны для объяснения происхождения TAS-сигналов на рис. 1а, спектры 2, 3. Дело в том, что ридимерная форма (a) в схеме (5) обладает VIS-поглощением при λmax = 535 нм (рис. 1a; 1б, кривые 2, 3), но практически не поглощает света в VIS-полосе ридимеров 1б и соответственно не испытывает фотовозбуждения в пучке лазерного света с λex = 400 нм. В акты фотовозбуждения вступают только ридимерные формы (b) и (f), несущие на себе четыре и три катиона CphAT, соответственно.
Согласно изложенному выше, фотопревращение пары (5ab) индуцируется возбуждением ридимера (b) (схема 4) с последующим появлением FK-состояний (схема 3), которые ответственны за широкую полосу VIS-поглощения от 470 до 650 нм с λmax = 535 нм (рис. 1а, кривая 2). При этом, несмотря на исчезновение ридимера (1b), кривая 2 (рис. 1а) не содержит полосы отбеливания. Это можно объяснить тем, что фотовозбуждение ридимера (b) в ридимерной паре (5ab) приводит к образованию колебательно возбужденных FK-состояний, передающих энергию колебаний на соседнюю форму (а) и на капсулу γ-CD. Как видно, получаемая парой (а) энергия достаточна для перевода ее в пару (b). Таким образом, появление новой пары (b), компенсирует исчезновение фотовозбужденной пары (b), тем самым, компенсируя и полосу отбеливания. Одновременно с этим потеря VIS-полосы с λmax = = 535 нм, связанная с исчезновением пары (a), компенсируется появлением такой же полосы, принадлежащей FK-состояниям ${\text{3a}}_{{\mathbf{r}}}^{{\mathbf{v}}} \leftrightarrow {\text{3b}}_{{\mathbf{r}}}^{{\mathbf{v}}}$ (схема 3).
Все описанные выше трансформации ридимеров (а) повторяются и в ридимерной паре (5af) при действии света с λex = 400 нм. Нетрудно показать, что фотовозбуждение ридимерной пары (f), содержащей три катиона CphAT, тоже ведет к ее расщеплению на FK-пары. При этом энергия возбуждения пары (f) достаточна для изменения электронной конфигурации атома N из состояния 1s22s22p3 в состояние с sp2-гибридизацией (N*R2 → NR2), а также для восстановления его сопряжения с фениленовым кольцом и, в итоге, появления фотовозбужденного ридимера (b), претерпевающего далее превращения в соответствии со схемами (3, 4). В данном случае важно, что взамен исчезающих трех катионов CPhAT в ридимерной паре (5af) появляются четыре катиона CphAT в результате перехода (a) → (b). Этим можно объяснить появление относительно сильного транзитного сигнала TAS с полосой VIS-поглощения в области исходного ридимера (b) с λmax ~ 460 нм (рис. 1а, кривая 3).
В [11] отмечено, что этот транзитный сигнал релаксирует очень медленно. Данный процесс можно связать с тем, что капсулы 2γ-CD + + 2MOD2 (схема 5, af), получив некоторое структурное искажение при фотовозбуждении, постепенно восстанавливают механическое давление на ридимерную пару и соответственно возвращают состояние с электронной конфигурацией азота 1s22s22p3. Сходный, но значительно менее интенсивный сигнал TAS при λmax ~ 460 нм наблюдается и на кривой 2, рис. 1а. Отсюда можно заключить, что и при низкой концентрации γ‑CD (3 ммоль/л) одновременно с комплексами (5ab) присутствует некоторое количество комплексов (5af).
Выше отмечалось, что наличие отрицательных значений у сигналов UTL от MOD в воде и от комплексов с α- и β-CD авторы [11] объясняли образованием c-MOD. Проведенный в настоящей работе анализ показал отсутствие транс–цис-изомеризации MOD2 в экспериментах [11]. По нашему мнению, отрицательные и медленно релаксирующие к нулевой интенсивности сигналы UTL [11] следует объяснять в соответствии с представлениями работы [16].
Согласно [16], имеются два вида сигналов UTL в водных растворах. UTL-сигналы со значениями τ порядка нескольких пс обусловлены образованием транзитных линз с отрицательным фокусом за счет практически мгновенного повышения температуры без изменения плотности среды в зонах релаксации энергии фотовозбуждения. Затем появляются другие линзы с отрицательным фокусом, связанные с изменением плотности нагретой среды и имеющие значительно более медленную динамику релаксации, ~500 пс.
Таким образом, в результате анализа TAS- и UTL-сигналов установлено строение комплексов включения MOD2 в γ-CD. Кроме того, на основании совпадения кинетических компонент UTL-сигналов от MOD2 в водной среде и в комплексах с α- и β-CD получено подтверждение факта формирования аквакапсул вокруг гидрофобных молекул и ридимеров. В конечном итоге результаты проведенного анализа позволяют утверждать, что концепция ридимерного строения создает адекватный фундамент для объяснения не только спектроскопии аминоазобензольных красителей и закономерностей фотохимического превращения аминоазобензола, но также и фотопревращения его сульфопроизводного MOD2.
Список литературы
Михеев Ю.А., Гусева Л.Н., Ершов Ю.А. // Журн. физ. химии. 2017. Т. 91. № 4. С. 672.
Михеев Ю.А., Гусева Л.Н., Ершов Ю.А. // Там же. 2017. Т. 91. № 10. С. 1683.
Михеев Ю.А., Ершов Ю.А. // Там же. 2018. Т. 92. № 2. С. 267.
Михеев Ю.А., Ершов Ю.А. // Там же. 2018. Т. 92. № 8. С. 1251.
Михеев Ю.А., Ершов Ю.А. // Там же. 2018. Т. 92. № 10. С. 1552.
Михеев Ю.А., Ершов Ю.А. // Там же. 2019. Т. 93. № 2. С. 369.
Михеев Ю.А., Ершов Ю.А. // Там же. 2019. Т. 93. № 6. С. 946.
Михеев Ю.А., Ершов Ю.А. // Там же. 2019. Т. 93. № 7. С. 1111.
Hirose Ya., Yui H., Sawada Ts. // J. Phys. Chem. A. 2002. V. 106. № 13. P. 3067.
Hirose Ya., Yui H., Fujinami M., Sawada Ts. // Chem. Phys. Letters. 2001. № 341. № 1–2. P. 29.
Takei M., Yui H., Hirose Ya., Sawada Ts. // J. Phys. Chem. A. 2001. V. 105. № 51. P. 11395.
Szejtli J. // Chem. Rev. 1998. V. 98. № 5. P. 1743.
Михеев Ю.А., Гусева Л.Н., Ершов Ю.А. // Журн. физ. химии. 2005. Т. 79. № 4. С. 583.
Михеев Ю.А., Гусева Л.Н., Давыдов Е.Я., Ершов Ю.А. // Там же. 2007. Т. 81. № 12. С. 2119.
Симон Ж., Андре Ж.-Ж. Молекулярные полупроводники. М.: Мир, 1988. С. 275.
Terazima M. // J. Chem. Phys. 1996. V. 105. № 16. P. 6587.
Дополнительные материалы отсутствуют.
Инструменты
Журнал физической химии