

Генетика, 2021, T. 57, № 2, стр. 205-213
Генетическая структура популяции голштинизированного черно-пестрого скота по микросателлитным локусам
О. С. Шаталина 1, *, И. В. Ткаченко 1, А. А. Ярышкин 1
1 Уральский федеральный аграрный научно-исследовательский центр
Уральского отделения Российской академии наук
620142 Екатеринбург, Россия
* E-mail: shatalinao@list.ru
Поступила в редакцию 12.03.2020
После доработки 01.05.2020
Принята к публикации 25.08.2020
Аннотация
Приведены результаты исследования генетической структуры популяции крупного рогатого скота по 12 микросателлитным локусам. Объект исследования – 702 особи крупного рогатого скота голштинизированной черно-пестрой породы. Установлено, что в среднем гетерозиготность животных по локусам составляет 78.3%. Среднее число аллелей на локус составляет 7.0 ± 0.08. Наибольшее распространение в популяции получил аллель 248 пн локуса SPS115 – 0.644. Генотип 248/248 локуса SPS115 имеет наибольшую частоту встречаемости – 0.446. В популяции выявлено 169 редких генотипов с частотой встречаемости меньше 0.010.
Популяционная генетика представляет большой интерес для биологов, так как дает новые знания о генетической структуре популяций различных видов животных, селекционных процессах, дрейфе генов, степени инбридинга, оценке генетических расстояний между семействами, генетическом разнообразии. Межпородное скрещивание приводит к тому, что в популяцию животных привносятся новые гены. Закрепление ценных хозяйственно полезных признаков при помощи инбридинга способствует увеличению количества гомозиготных особей в популяции.
Генетическая структура популяции исследуется по генам и микросателлитным локусам. Изучается сцепленное наследование генов, локусов, изменчивость и ассоциации с хозяйственно полезными признаками. Так, Н.В. Ковалюк с соавт. [1] изучена генетическая изменчивость гена BoLA‑DRB3 крупного рогатого скота. С.Р. Хатами с соавт. [2] исследован полиморфизм генов пролактина и гормона роста и их взаимосвязь с хозяйственно полезными признаками. Также учеными изучены фрагменты ДНК, фланкированных микросателлитными локусами с целью выявления породных маркеров крупного рогатого скота, овец и других животных [3–6].
В последнее десятилетие широкое распространение получили исследования микросателлитных локусов. Т.Ю. Киселева с соавт. [7] выявили неравновесие по сцеплению локусов крупного рогатого скота. Проведены исследования генетического разнообразия крупного рогатого скота различных пород по микросателлитным локусам [8–11]. Изучены количественные показатели аллелей. Выявлено, что число аллелей на локус варьирует от 7 до 18 [12–16]. Также исследования проведены на популяции лошадей, овец, лосей и оленей [17–20].
Цель настоящей работы – определение генетической структуры популяции голштинизированного черно-пестрого скота по микросателлитным локусам.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование проведено в Уральском НИИСХ – филиале ФГБНУ УрФАНИЦ УрО РАН в 2019 г. Объектом исследования служил голштинизированный черно-пестрый скот (Bos taurus) (n = 702). ДНК выделяли из цельной крови с антикоагулянтом при помощи набора ДНК–Экстран 1 (ООО “НПФ Синтол”, Россия) согласно методике изготовителя. ПЦР проводили на амплификаторе PCR-9700. Определены 12 микросателлитных локусов, рекомендованных Международным обществом генетики животных (ISAG): BM1824, BM2113, ETH3, ETH10, ETH225, INRA23, SPS115, TGLA53, TGLA122, TGLA126, TGLA227, BM1818, с использованием набора CorDIS Cattle (ООО “Гордиз”, Россия). Микросателлитный профиль животных установлен при использовании генетического анализатора Genetic Analyzer АВ 3130 и программ Data Collection v.3.1 и GeneMapper v.4.0. В ходе изучения генетической структуры популяции крупного рогатого скота выявлены: количество гомозиготных и гетерозиготных животных по локусам, а также аллели и генотипы по каждому локусу. Информативными считали аллели, имеющие частоту встречаемости выше 5%. Статистический анализ проведен при помощи программ IBM SPSS Statistics 23 и MicrosoftExcel.
РЕЗУЛЬТАТЫ
Исследована гомозиготность крупного рогатого скота голштинизированной черно-пестрой породы по 12 микросателлитным локусам. Результаты отражены в табл. 1.
Таблица 1.
Структура популяции крупного рогатого скота по гомозиготности микросателлитных локусов (n = 702)
| Микросателлитные локусы | Особи, % | |
|---|---|---|
| гетерозиготные | гомозиготные | |
| BM1824 | 78.2 | 21.8 |
| BM2113 | 87.2 | 12.8 |
| ETH3 | 75.4 | 24.6 |
| ETH10 | 75.8 | 24.2 |
| ETH225 | 78.7 | 21.3 |
| INRA023 | 80.5 | 19.5 |
| SPS115 | 56.9 | 43.1 |
| TGLA53 | 93.9 | 6.1 |
| TGLA122 | 86.0 | 14.0 |
| TGLA126 | 70.8 | 29.2 |
| TGLA227 | 87.9 | 12.1 |
| BM1818 | 69.0 | 31.0 |
| Среднее | 78.3 | 21.6 |
Выявлено, что количество гетерозиготных особей значительно превышает количество гомозиготных по всем 12 локусам. По локусу TGLA53 обнаружено наименьшее количество гомозиготных особей – 6.1%. Наибольший процент гомозиготных животных выявлен в локусе SPS115 – 43.1%. Также установлены высокие показатели гомозиготности в локусах BM1818 и TGLA126 – 31 и 29.2% соответственно. В среднем по популяции гетерозиготность составила 78.3%, гомозиготность – 21.6%. Аналогичные данные получены Т.Н. Карымсаковым с соавт. [13].
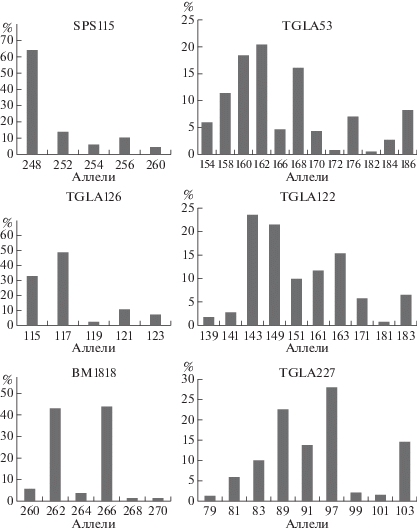
В микросателлитных локусах распространен множественный аллелизм. На рис. 1, 2 представлен полиморфизм аллелей по 12 микросателлитным локусам. В локусах BM1824, SPS115, TGLA126, BM1818 выявлено наименьшее число аллелей – пять. В локусе TGLA53 установлено наибольшее число аллелей – 12, что свидетельствует о высоком генетическом разнообразии [21, 22]. В среднем число аллелей на локус составляет 7.0 ± 0.08. Аналогичные данные получены Н.И. Стрекозовым с соавт., Н.С. Фураевой с соавт. на крупном рогатом скоте сычевской, бурой швицкой и ярославской пород, число аллелей на локус составило 7.4 [11, 23]. Исследования, проведенные учеными на герефордской и симментальской породах, выявили более низкое число аллелей на локус – 3.3 в герефордской породе и 5.3 в симментальской породе [24, 25]. В популяции голштинизированного крупного рогатого скота наибольшую распространенность в общем количестве локусов получил аллель 248 пн локуса SPS115 (0.644), наименьшую – аллель 141 пн локуса BM2113 (0.001).
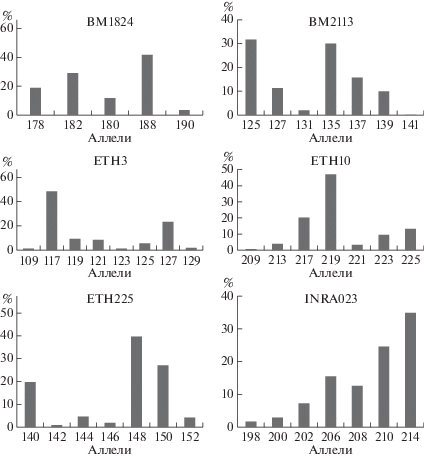
Рис. 2.
Частоты встречаемости (%) аллелей локусов SPS115, TGLA53, TGLA122, TGLA126, TGLA227, BM1818.
По локусу BM1824 наиболее часто присутствует аллель 188 пн, его частота встречаемости составляет 0.405. Редким аллелем данного локуса является аллель 190 – 0.026. В локусе установлено четыре информативных аллеля – 178, 180, 182, 188 пн и один низкоинформативный аллель – 190 пн. По локусу BM2113 распространены аллели 135 и 125 пн (0.298 и 0.314 соответственно). Редким аллелем в данном локусе является 141 пн – 0.001, возможно данный аллель появился вследствие мутационного процесса. В локусе присутствуют пять информативных аллелей –125, 127, 135, 137, 139 пн и два низкоинформативных аллеля – 131 и 141 пн. В ярославской породе у животных имеется большее разнообразие по данному локусу – 10 аллелей, в черно-пестрой голштинизированной породе – семь аллелей [23]. В локусе ETH3 получил распространение аллель 117 пн – 0.480. Редко встречаются аллели 123 и 109 пн, по 0.013 каждый. По локусу выявлены пять информативных аллелей – 117, 119, 127, 121, 125 пн и три аллеля с низкой информативностью – 129, 123 и 109 пн. В ярославской породе данный локус представлен меньшим генетическим разнообразием – шесть аллелей, в то время как в черно-пестрой голштинизированной породе – восемь аллелей [23]. По локусу ETH10 часто встречается аллель 219 пн – 0.472, наиболее редким в данном локусе является аллель 209 – 0.008. В локусе ETH10 присутствуют четыре информативных аллеля – 219, 223, 225 и 217 пн и три низкоинформативных – 221, 213, 209 пн.
Локус TGLA53 представлен большим числом аллелей – 12, в ярославской породе животные по данному локусу имеют 10 аллелей [23]. Наибольшее распространение получили аллели 168, 160 и 162 пн (0.161, 0.183 и 0.200 соответственно). Редкими аллелями в данном локусе являются 182 и 172 пн – 0.005 и 0.008 соответственно. По данному локусу выявлено семь информативных аллелей – 154, 158, 160, 162, 168, 176 и 186 пн и пять аллелей с низкой информативностью – 166, 172, 182, 184 и 170 пн. В локусе TGLA122 животных черно-пестрой голштинизированной породы наблюдается большое генетическое разнообразие – 10 аллелей, в симментальской породе в данном локусе выявлено шесть аллелей [25]. При этом наибольшее распространение имеют аллели 149 и 143 пн – 0.215 и 0.235 соответственно, хотя их концентрация в популяции ниже, чем у других распространенных аллелей локусов. Редкими аллелями локуса являются 181 и 139 пн – 0.008 и 0.019. В локусе TGLA122 установлено семь информативных аллелей – 149, 143, 171, 183, 163, 151 и 161 пн. Низкоинформативными показали себя аллели с длиной 181, 139 и 141 пн. Распространенным аллелем локуса ETH225 признан аллель 148 (0.397), редким – 146 пн (0.024). В данном локусе присутствуют три информативных аллеля – 140, 148 и 150 пн, причем аллель 148 является наиболее информативным. Аллели 142, 144, 152 и 146 пн обладают низкой информативностью.
По локусу INRA023 наиболее распространен аллель 214 пн. Редко встречается аллель 198 – 0.019. По локусу INRA023 выявлено пять информативных аллелей – 202, 206, 208, 210 и 214 пн и два низкоинформативных – 198 и 200 пн. В локусе SPS115 установлен самый распространенный аллель – 248 пн, который обладает высокой информативностью. Редких аллелей (менее 0.02) в данном локусе не выявлено. Информативными аллелями локуса являются 248, 256, 254 и 252 пн. В симментальской породе также данный аллель 248 является самым распространенным, его частота встречаемости составляет 0.778 [25]. По локусу TGLA126 наиболее распространен аллель 117 пн – 0.486. Редким аллелем локуса является аллель 119 пн – 0.020. Данный аллель – единственный неинформативный аллель локуса TGLA126. В локусе TGLA227 наблюдается большое разнообразие – девять аллелей. Наибольшей распространенностью обладает аллель 97 пн – 0.28. Редкими аллелями локуса являются 79 и 101 пн – 0.013 и 0.016 соответственно. В данном локусе присутствуют шесть информативных аллелей – 89, 91, 83, 97, 81 и 103 пн. Три аллеля являются неинформативными – 101, 79 и 99 пн. В локусе BM1818 лидирующее положение заняли аллели 262 и 266 – 0.430 и 0.444 соответственно. Реже других встречаются аллели 268 и 270 пн – 0.14 каждый. Аллели 262 и 266 пн данного локуса можно считать высокоинформативными, аллели 268 и 270 пн – низкоинформативными. В популяции крупного рогатого скота голштинизированной черно-пестрой породы число аллелей на локус варьировало от пяти до 12. Необходимо отметить, что в бурой швицкой породе крупного рогатого скота аллель 262 локуса BM1818 – самый распространенный, его частота встречаемости составляет 0.680 [11], в то время как в черно-пестрой голштинизированной породе самый распространенный аллель 248 – 0.644. Кроме того, несмотря на меньшее число аллелей на локус в герефордской породе, в локусе BM1818 присутствуют девять аллелей, а в черно-пестрой голштинизированной – шесть аллелей [24]. При этом в симментальской породе данный локус представлен меньшим разнообразием аллелей – четыре аллеля на локус [25].
Также исследованы генотипы микросателлитных локусов популяции крупного рогатого скота (рис. 3). По локусу TGLA122 у особей крупного рогатого скота установлено 70 генотипов, что свидетельствует о высоком полиморфизме локуса. Генетическая структура популяции по данному локусу состоит из 61 гетерозиготного и восьми гомозиготных генотипов: 149/149, 143/143, 151/151, 163/163, 141/141, 183/183, 139/139 и 171/171. Наибольшая частота встречаемости отмечена у генотипа 143/149 – 0.108. В популяции обнаружено большое число редких генотипов – 41. Установлено 15 генотипов с частотой встречаемости 0.001 (143/181, 161/163, 153/149, 161/181, 173/183, 171/173, 171/181, 147/149, 139/149, 151/153, 149/175, 141/153, 153/171, 139/139 и 161/169), а также 13 генотипов с частотой встречаемости 0.002 (153/183, 147/163, 151/181, 143/175, 181/183, 149/173, 141/161, 183/183, 139/141, 141/163, 143/147, 141/151, 171/171). В популяции крупного рогатого скота по локусу TGLA122 выявлено шесть генотипов с частотой встречаемости 0.004 (141/171, 143/173, 153/163, 141/141, 163/181, 139/143) и три генотипа с частотой встречаемости 0.005 (139/161, 139/151, 139/163). Частота встречаемости генотипов 171/183 и 139/183 составляет 0.006, 141/149 и 141/183 – 0.008.
Рис. 3.
Процентное соотношение распространенных генотипов локусов TGLA122, TGLA227, TGLA126, BM1818.
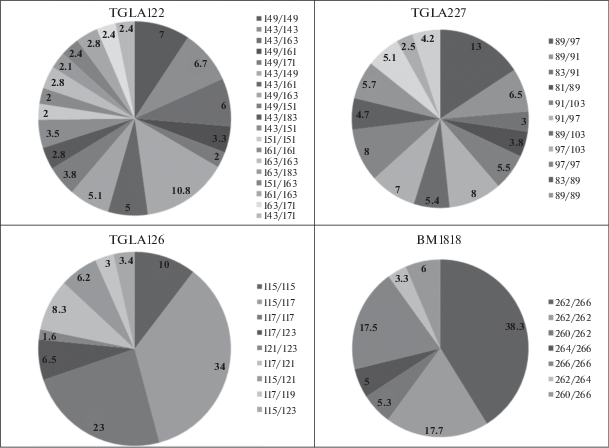
Локус TGLA227 представлен большим генетическим разнообразием – в нем установлено 53 генотипа, что также позволяет предположить о его высоком полиморфизме. При этом гомозиготных генотипов по локусу всего четыре – 97/97, 89/89, 81/81 и 83/83 и 49 генотипов – гетерозиготны. Самый распространенный генотип данного локуса – 89/97 (0.130). Также в популяции крупного рогатого скота по локусу TGLA227 имеется большое число генотипов с частотой встречаемости менее 0.01. Так, 11 генотипов имеют частоту встречаемости 0.001 – 99/103, 79/91, 77/83, 97/101, 77/91, 79/97, 89/101, 81/87, 91/95, 83/101 и 99/101. У 8 генотипов установлена частота встречаемости 0.002 – 91/101, 101/103, 87/103, 81/99, 81/81, 87/91, 83/99, 87/93. Выявлены пять генотипов с частотой встречаемости 0.004 – 79/103, 83/87, 81/93, 77/89, 77/103. Частота встречаемости генотипов 83/93, 89/99, 91/93, 87/89, 93/103 и 83/83 составляет 0.005. Генотипы 97/99, 87/97, 81/83 и 81/91 имеют частоту встречаемости 0.008, генотипы 81/83 и 77/97 – 0.007, а генотип 103/103 – 0.009.
В локусе TGLA126 представлено 16 генотипов, среди них – четыре гомозиготных: 115/115, 117/117, 121/121 и 123/123 и 12 гетерозиготных. Наибольшее распространение получили четыре генотипа – 115/115, 117/121, 117/123 и 115/121 – 0.100, 0.083, 0.065 и 0.063 соответственно. Наиболее редкие генотипы данного локуса: 107/117, 111/121 – по 0.001 каждый; 123/123 – 0.002; 119/123, 119/121 – по 0.004 каждый.
В локусе BM1818 установлено 17 генотипов. Наиболее распространенными являются генотипы 262/266, 262/262 и 266/266 – 0.383, 0.177 и 0.175 соответственно. Редкими генотипами по данному локусу являются 260/268, 260/260 и 258/262 – по 0.001 каждый; 264/268 – 0.002; 258/268 – 0.005 и 260/264–0.007. Частота встречаемости данных генотипов меньше одного процента. В популяции по локусу BM1818 выявлено три гомозиготных генотипа – 262/262, 266/266, 260/260 и 14 гетерозиготных.
Локус BM1824 представлен 15 генотипами. Гомозиготных генотипов по данному локусу четыре – 182/182, 180/180, 188/188 и 178/178. Наибольшую частоту встречаемости имеют генотипы 182/188 и 188/188 – 0.181 и 0.180 соответственно. Также выявлено три генотипа с низкой встречаемостью – 174/182 (0.001), 180/190 (0.007) и 178/190 (0.008) (рис. 4).
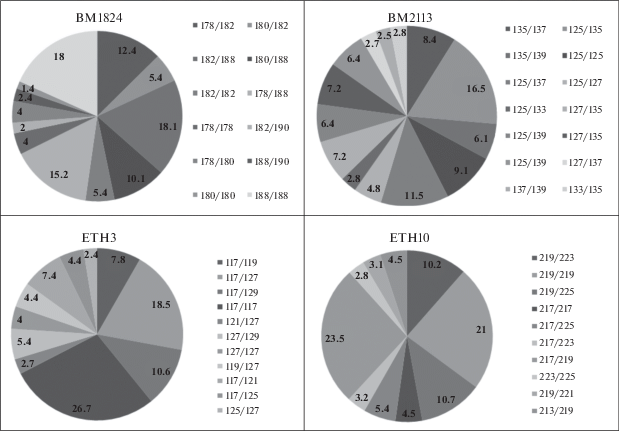
В популяции крупного рогатого скота по локусу BM2113 установлено 19 генотипов. Наибольшее распространение получил генотип 125/135 – 0.165, наименьшее – генотип 133/139 – 0.007. Установлено три гомозиготных по данному локусу генотипа – 125/125, 137/137 и 127/127.
Локус ETH3 в популяции представлен 18 генотипами. Гомозиготные генотипы в данной популяции – 117/117, 119/119, 129/129 и 121/121. Наибольшую частоту встречаемости имеет генотип 117/117 – 0.267. Также установлены пять генотипов с низкой частотой встречаемости: 119/119 и 121/121 – 0.004, 119/125 – 0.005, 129/129 и 121/125 – 0.008 каждый.
По локусу ETH10 выявлен 21 генотип, среди них четыре гомозиготных – 219/219, 217/217, 225/225 и 223/223. Наибольшее распространение имеет генотип 217/219 – 0.235. Также выявлены пять редко встречающихся генотипов – 209/225 и 213/223 – 0.002, 209/217 – 0.004, 223/223 и 209/219 – 0.008 каждый.
Локус ETH225 представлен в данной популяции 18 генотипами. Установлено три гомозиготных генотипа (150/150, 140/140 и 148/148) и 15 гетерозиготных. Наибольшее распространение получил генотип 148/150 – 0.301. Также установлено три редко встречающихся генотипа – 146/152 (0.002), 140/142 (0.005) и 142/150 (0.007) (рис. 5).
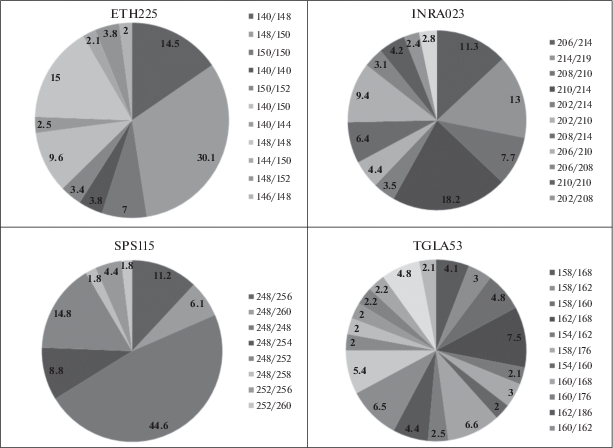
Исследованы особи крупного рогатого скота по локусу INRA023. Всего установлено 29 генотипов, среди них четыре гомозиготных (202/202, 210/210, 208/208 и 206/206) и 25 гетерозиготных. Наибольшую частоту встречаемости имеет генотип 210/214 – 0.182. Также выявлено 11 генотипов с частотой встречаемости меньше 0.01. Два генотипа имеют частоту встречаемости 0.002 (198/202, 212/214), пять генотипов (198/208, 198/206, 202/204, 200/208, 204/208) – частоту встречаемости 0.004, и генотип 200/206 – 0.008.
Локус SPS115 в популяции крупного рогатого скота черно-пестрой породы представлен 16 генотипами. Установлено четыре гомозиготных генотипа (248/248, 252/252, 256/256 и 260/260) и 12 гетерозиготных. Наибольшее распространение получил генотип 248/248 – 0.446. Также установлено пять редких генотипов. Частота встречаемости генотипов 254/258 и 260/260 составляет 0.002, 256/258 и 256/260 – 0.004 и 256/256 – 0.005.
В популяции крупного рогатого скота по локусу TGLA53 обнаружено 80 генотипов. Данный локус ввиду значительного количества аллелей и генотипов возможно отнести к высокополиморфным. Число гомозиготных генотипов у особей – восемь (168/168, 162/162, 160/160, 176/176, 186/186, 154/154, 170/170, 166/166), гетерозиготных – 72. Наибольшую частоту встречаемости имеет генотип 162/168 – 0.075. Также в популяции установлено 48 редких генотипов. У исследованных животных присутствуют 18 генотипов с частотой встречаемости 0.001 (172/184, 154/174, 168/174, 178/184, 186/186, 154/184, 158/182, 154/154, 172/186, 158/174, 170/170, 166/172, 162/172, 160/172, 164/176, 168/178, 176/182, 154/164) и 13 генотипов с частотой встречаемости 0.002 (160/182, 162/174, 154/178, 166/170, 184/186, 164/184, 160/174, 158/178, 164/186, 166/166, 158/164, 166/176, 158/158). Также выявлено семь генотипов с частотой встречаемости 0.004 (160/164, 154/170, 160/184, 164/168, 166/182, 154/166, 162/178) и три генотипа с частотой встречаемости 0.005 (170/184, 176/184 и 178/186). Частота встречаемости генотипов 158/184 и 154/176 составляет 0.007, 176/176 и 170/176 – 0.008 и 170/186, 168/170 и 158/170 – 0.009.
ОБСУЖДЕНИЕ
В ходе проведенных исследований генетической структуры популяции крупного рогатого скота черно-пестрой породы установлено, что гомозиготность по микросателлитным локусам варьирует от 6.1% по локусу TGLA53 до 43.1% по локусу SPS115. Число аллелей на локус в голштинизированной черно-пестрой породе варьирует от пяти до 12 и составляет в среднем семь аллелей. В сычевской, бурой швицкой и ярославской породах получены аналогичные данные по среднему числу аллелей на локус, в то время как в герефордской и симментальской породах данный показатель снижен до 3.3 и 5.3 аллеля на локус соответственно. Наибольшую распространенность среди аллелей 12 локусов в популяции имеет аллель 248 пн локуса SPS115 – 0.644. Редкими аллелями популяции являются аллель 141 пн локуса BM2113 – 0.001 и аллель 182 пн локуса TGLA53 – 0.005. Всего по 12 микросателлитным локусам выявлено 52 информативных аллеля. Наиболее распространенным генотипом является генотип 248/248 локуса SPS115 – 0.446. Высокая частота встречаемости данного генотипа позволяет назвать его характерным для голштинизированной черно-пестрой породы. Также у исследованных особей установлено 169 генотипов с частотой встречаемости меньше 0.01. Установлено три высокополиморфных локуса: TGLA53, TGLA227, TGLA122. Гомозиготные генотипы присутствуют как среди распространенных, так и среди редко встречающихся, что свидетельствует о том, что гомозиготное состояние генотипа не связано с его частотой встречаемости. Установлено, что при увеличении общего числа генотипов локуса с 15–18 до 70–80 возрастает количество редких аллелей в данных локусах с 0.20–0.30 до 0.58–0.68, что обусловлено большим генетическим разнообразием. Популяция крупного рогатого скота черно-пестрой голштинизированной породы представлена значительным числом аллелей и генотипов, что позволяет делать предположения об уровне инбридинга в данной популяции. Симментальская и ярославская породы крупного рогатого скота имеют меньшее генетическое разнообразие. По изученным микросателлитным локусам они имеют разницу на 1–2 аллеля на локус по сравнению с черно-пестрой голштинизированной породой.
Источником финансирования являлось выполнение государственного задания по теме: “Разработать селекционно-генетические и теоретические основы сохранения и эффективного использования генофонда крупного рогатого скота в Уральском регионе с применением современных биотехнологий”.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Ковалюк Н.В., Сацук В.Ф., Волченко А.Е. Изменчивость гена BoLA-DRB3 у крупного рогатого скота молочного направления продуктивности и его влияние на параметры жизнеспособности // Генетика. 2012. Т. 48. № 8. С. 962–965.
Хатами C.Р., Лазебный О.Е., Максименко В.Ф., Сулимова Г.Е. ДНК-полиморфизм генов гормона роста и пролактина у ярославского и черно-пестрого скота в связи с молочной продуктивностью // Генетика. 2005. Т. 41. № 2. С. 229–236.
Глазко В.И. Геномное распределение ISSR-маркеров (AG)9C и (GA)9C у видов Bovinae и Caprinae // С.-х. биология. 2009. Т. 44. № 4. С. 31–36.
Глазко В.И., Столповский Ю.А. Структурно-функциональные особенности микросателлитов в геномах крупного рогатого скота и овец // Докл. РАСХН. 2011. № 1. С. 41–45.
Столповский Ю.А., Ахани Азари M., Евсюков А.Н. и др. Сравнительный анализ полиморфизма ISSR-маркеров у пород крупного рогатого скота // Генетика. 2011. Т. 47. № 2. С. 213–226.
Столповский Ю.А., Ахани Азари M., Кол Н.В и др. Дифференциация генофонда пород крупного рогатого скота по ISSR-PCR-маркерам // Изв. Тимирязевской с.-х. академии. 2009. № 3. С. 89–97.
Киселева Т.Ю., Kantanen J., Воробьев Н.И. и др. Неравновесие по сцеплению микросателлитных локусов у шести локальных популяций крупного рогатого скота // Генетика. 2014. Т. 50. № 4. С. 464–473.
Волкова В.В., Денискова Т.Е., Костюнина О.В. и др. Характеристика аллелофонда локальных пород крупного рогатого скота России по микросателлитным маркерам // Генетика и разведение животных. 2018. № 1. С. 3–10. https://doi.org/10.31043/2410-2733-2018-1-3-10
Копылов К.В., Шелёв А.В., Копылова К.В., Березовский А.В. Генетическая структура популяций украинской черно-пестрой и украинской красно-пестрой молочных пород по полиморфизму QTL и STR маркеров // Вестник Сумского национ. аграрного ун-та. 2014. № 7. С. 31–37.
Нурбаев С.Д., Омбаев А.М., Карымсаков Т.Н. и др. Определение чистопородности популяций крупного рогатого скота мясного направления продуктивности по микросателлитным ДНК // Зоотехния. 2017. № 8. С. 10–13.
Стрекозов Н.И., Зиновьева Н.А., Горелов П.В. и др. Генетическая характеристика созданных типов скота бурой швицкой и сычевской пород с использованием полиморфизма микросателлитных локусов // С.-х. биология. 2009. Т. 44. № 2. С. 10–15.
Гайнуллина К.П. Некоторые аспекты применения микросателлитных маркеров в сельскохозяйственной практике // Изв. Оренбургского гос. аграрного ун-та. 2018. № 5. С. 232–234.
Карымсаков Т.Н., Гладырь Е.А., Нурбаев С.Д. и др. Сравнительная характеристика аллелофонда крупного рогатого скота трех родственных пород черно-пестрого корня, разводимых в республике Казахстан // Молочное и мясное скотоводство. 2017. № 3. С. 11–14.
Кольцов Д.Н., Волкова В.В., Гладырь Е.А. и др. Характеристика аллелофонда сычевской породы крупного рогатого скота по ДНК микросателлитам // Достижения науки и техники АПК. 2012. № 8. С. 56–57.
Модоров М.В., Ткаченко И.В., Грин А.А. Генетическая изменчивость голштинизированного черно-пестрого скота на территории Свердловской области // Вопр. нормативно-правового регулирования в ветеринарии. 2019. № 4. С. 111–113. https://doi.org/10.17238/issn2072-6023.2019.4.111
Часовщикова М.А., Шевелева О.М. Генетическая характеристика крупного рогатого скота герефордской породы тюменской области с использованием микросателлитных ДНК-маркеров // Вестник ИРГСХА. 2018. № 88. С. 141–150.
Чысыма Р.Б., Храброва Л.А., Зайцев А.М. и др. Оценка генетического разнообразия в популяциях тувинских лошадей по локусам систем крови и микросателлитным ДНК // С.-х. биология. 2017. Т. 52. № 4. С. 679–685. https://doi.org/10.15389/agrobiology.2017.4.679eng
Озеров М.Ю., Тапио М., Марзанов Н.С. и др. Микросателлитный анализ эволюционно-генетических связей у различных пород овец // Докл. РАСХН. 2006. № 2. С. 30–33.
Марзанов Н.С., Девришов Д.А., Марзанова С.Н. и др. Популяционно-генетическая характеристика лосей по локусам микросателлитов // Пробл. биологии продуктивных животных. 2018. № 1. С. 75–82. https://doi.org/10.25687/1996-6733.prodanimbiol.2018. 1.75-82
Баранова А.И., Холодова М.В., Сипко Т.П. Генетическая структура дикого северного оленя (Rangifer tarandus) России на основании полиморфизма микросателлитных локусов: Материалы Всерос. науч. конф., посвященной 70-летнему юбилею кафедры “Зоология и экология” Пензенского гос. ун-та и памяти профессора В.П. Денисова (1932–1997) “Актуальные вопросы современной зоологии и экологии животных”. Пенза, 2016. 21 с.
Марзанов Н.С., Озеров М.Ю., Насибов М.Г., Марзанова Л.К. Микросателлиты и их использование для оценки генетического разнообразия животных (обзор иностранной литературы) // С.-х. биология. 2004. Т. 39. № 2. С. 104–111.
Митютько В.И., Грачев В.С. Генетическое разнообразие у сельскохозяйственных животных и механизмы его изменения // Изв. Санкт-Петербургского гос. аграрного ун-та. 2011. № 23. С. 140–146.
Фураева Н.С., Ганченкова Т.Б., Кертиев Р.М., Калашникова Л.А. Генетическая гетерогенность быков-производителей ярославской породы по маркерам ДНК // Молочное и мясное скотоводство. 2016. № 6. С. 2–5.
Седых Т.А., Глазырь Е.А., Долматова И.Ю. и др. Полиморфизм микросателлитных локусов крупного рогатого скота герефордской породы различных эколого-генетических генераций // Вестник АПК Ставрополья. 2014. № 3(15). С. 121–128.
Денискова Т.Е., Волкова В.В., Костюнина О.В. и др. Характеристика аллелофонда популяции симментальского и помесного скота Поволжья с использованием микросателлитов // Вестник Алтайского гос. аграрного ун-та. 2019. № 3(173). С. 100–106.
Дополнительные материалы отсутствуют.