

Лесоведение, 2019, № 3, стр. 198-207
Индивидуальная и географическая изменчивость шишек и формы семенных чешуй ели сибирской в сибирской части ареала
М. Н. Казанцева a, b, С. П. Арефьев a, b, П. П. Попов a, *
a Институт проблем освоения Севера Тюменского НЦ СО РАН
625003 Тюмень, ул. Малыгина, 86,
Россия
b Тюменский государственный университет
625003 Тюмень, ул. Володарского, 6,
Россия
* E-mail: ipospopov@mail.ru
Поступила в редакцию 25.12.2017
После доработки 05.03.2018
Принята к публикации 05.06.2018
Аннотация
Ель сибирская (Picea obovata Ledeb.) на территории Сибири в современный период характеризуется некоторыми особенностями: у нее сравнительно короткие шишки; показатели формы семенных чешуй существенно отличаются от ели европейской и популяций ели сибирской в европейской части ареала. Внутрипопуляционная (индивидуальная) изменчивость длины шишек несколько больше, чем географическая изменчивость, величина коэффициента сужения (Cn) значительно больше, а коэффициента вытянутости верхней части семенных чешуй (Cp) меньше, чем у ели европейской. Географическая изменчивость их очень низкая (около 4%), а внутрипопуляционная изменчивость в 2.5–3 раза выше. На всей территории региона показатель Cn больше показателя Cp, а разность их, как комплексного показателя признака, в среднем равна 25% при довольно высокой внутрипопуляционной изменчивости и почти вдвое меньшей географической. Отрицательная корреляция показателей Cn и Cp в пределах популяций среднего уровня (–0.328–0.694) прямолинейная. Сходство (корреляция) в географической изменчивости средних показателей длины шишек и формы семенных чешуй почти того же уровня. Результаты исследования могут быть полезны для решения вопросов популяционно-географической дифференциации ели сибирской с точки зрения лесоводства.
Ель сибирская (Picea obovata Ledeb.) за Уралом распространена до Охотского побережья (Атлас …, 1973; Соколов и др., 1977). При этом леса, образованные этим видом, составляют в структуре лесного фонда сравнительно небольшую часть (Цепляев, 1961) и распространены крайне неравномерно (Атлас …, 1973). В лесоводческом отношении ель сибирская в сибирской части ареала до сих пор изучена значительно меньше, чем близкая к ней ель европейская. Это обусловлено, прежде всего, обширностью территории и труднодоступностью участков. В.Н. Сукачев (1938) отмечал крайнюю желательность изучения ели сибирской и высказывал предположение, что при более детальном ее исследовании, вероятно, придется выделить географические расы, различающиеся не менее, чем ели европейская и сибирская.
Давно сложилось мнение, что ель сибирская мало отличается от ели европейской по многим признакам, и ряд ботаников, вслед за Ф.А. Теплоуховым (Teplouhoff, 1868; Теплоухов, 1872), рассматривают их в качестве одного вида – ели обыкновенной (Schmidt-Vogt, 1977). По имеющимся данным ель в Сибири в значительной степени отличается от ели европейской по параметрам морфологических, физиологических (Попов, 1999), генетических признаков (Кравченко и др., 2008, 2013; Гончаренко, Падутов, 2001), структуре хромосомного аппарата (Круклис, 1971; Муратова, Владимирова, 2001). Наиболее существенные отличия этих двух видов заключаются в форме семенных чешуй и длине шишек, давно используемых в качестве диагностических признаков. В ботанической и лесоводческой литературе обычно указывается, что длина шишек ели сибирской составляет 6–8 см, а форма семенных чешуй характеризуется так, как описал ее еще в 1843 г. К.Ф. Ледебур (K.F. Ledebour): “шишки цилиндрические, чешуйки овальные, на конце закругленные и цельные, снаружи несколько выпуклые” (цит. по (Теплоухов, 1872; Кеппен, 1885). Однако на таких больших пространствах, как Сибирь, возможно, ель не так однообразна, как предполагается. Несомненно, что помимо географической изменчивости ели сибирской имеется изменчивость внутрипопуляционная, т.е. индивидуальная (Мамаев, 1972) между деревьями в пределах популяций. На необходимость дальнейшего изучения ели сибирской указал и Л.И. Милютин (2015). Целью настоящей работы является изучение индивидуальной (внутрипопуляционной) и межпопуляционной (географической) изменчивости длины шишек и формы семенных чешуй на большей части ареала ели сибирской в Сибири.
ОБЪЕКТЫ И МЕТОДИКА
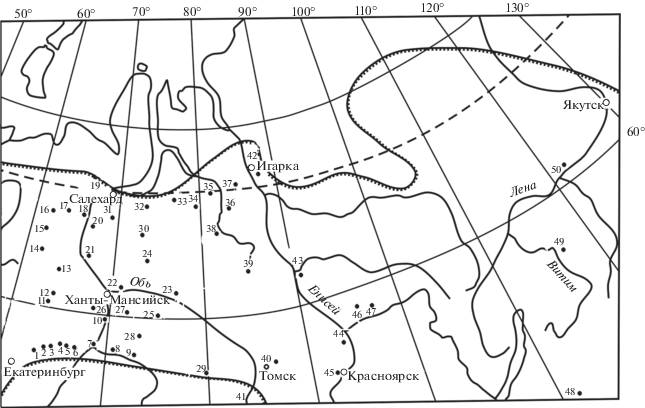
Исходными материалами для изучения изменчивости ели сибирской послужили 50 популяционных выборок шишек на обширной территории – от Урала до Якутии. Большая часть их приходится на Западную Сибирь (рис. 1), где еловые леса распределены сравнительно кучно. В большинстве выборок число деревьев, с которых собраны шишки, 100 и более, а общее их число составляет около 7.7 тыс. шт. Методика сбора и обработки материалов опубликована (Попов, 1999, 2011; Попов и др., 2015а). Длину шишек определяли измерением линейкой, а форму чешуй характеризовали на их проекциях по величине коэффициентов сужения (Cn) и вытянутости (Cp) их верхней (наружной) части. Коэффициент Cn определяется в виде отношения ширины чешуйки (d) на 0.1 наибольшей ее величины (D) от верхнего края к D, обычно в процентах:
Рис. 1.
Расположение пунктов (1–50) сбора материалов для изучения изменчивости ели на территории Сибири: 1 – Юшала, 2 – Тугулым, 3 – Успенское, 4 – Леваши, 5 – Велижаны, 6 – Янтык, 7 – Тобольск, 8 – Вагай, 9 – Дубровное, 10 – Уват, 11 – Куминский, 12 – Междуреченский, 13 – Зеленоборск, 14 – Усть-Манья, 15 – Саранпауль, 16 – Хулга, 17 – Овгорт, 18 – Ямгорт, 19 – Салехард и Лабытнанги, 20 – Полноват, 21 – Октябрьское, 22 – Ханты-Мансийск, 23 – Нижневартовск, 24 – Когалым, 25 – Угут, 26 – Чембакчина, 27 – Салым, 28 – Демьянка, 29 – Кыштовка, 30 – Нумто, 31 – Полуй, 32 – Надым, 33 – Новый Уренгой, 34 – Самбург, 35 – Тазовский, 36 – Красноселькуп, 37 – Сидоровск, 38 – Толька, 39 – Ратта, 40 – Томск, 41 – Горно-Алтайск, 42 – Игарка, 43 – Подкаменная Тунгуска, 44 – Енисейск, 45 – Красноярск, 46 – Бедоба, 47 – Дербина, 48 – Кыра, 49 – Бодайбо, 50 – Олекминск.
Cp определяется отношением расстояния (h) от верхнего края до положения наибольшей ширины (D) к ней же:
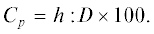
Исходные абсолютные размеры чешуй на их проекции (рис. 2) определяли с точностью 0.1 мм. Параметр 0.1D при определении коэффициента Cn используется в результате подбора, который подходил бы к чешуйкам разной формы. Отношение d и h к ширине семенных чешуй точнее характеризует форму их верхней части, чем отношение h к общей длине чешуй, которое использовалось некоторыми исследователями (Borchetti et al., 1988) при определении коэффициента вытянутости верхней (наружной) части чешуй. Разность показателей Cn и Cp или их отношение (Попов, 1999), как комплексный показатель признака полнее отражает его изменчивость. Подобный прием введен в широкую практику.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для изучения индивидуальной (внутрипопуляционной) изменчивости длины шишек и показателей формы семенных чешуй использовали часть выборок (16 пунктов) из разных географических районов: южных (№№ 3, 4, 9, 29, 45, 49), северных (19, 32, 42, 50), и из средней части региона (21, 26, 40, 43, 44, 48). Для характеристики изменчивости признаков приведены два показателя: предельные, или крайние значения (Lim.) и коэффициент вариации (Cv). Максимальное значение длины шишек (Lc) в кроне деревьев (около 90 мм в среднем по 16 выборкам) превосходит минимальное (около 45 мм) в 2 раза (табл. 1). Средние значения коэффициента вариации составляют 11–15%. Изменчивость показателей Cn и Cp заметно меньше. Здесь максимальное значение превосходит минимальное в 1.62–1.65 раза. Коэффициент вариации показателя Cn составляет 6–14, а показателя Cp – 8–11%, т.е. вдвое меньше, чем в европейской части ареала (Попов, 1999). Изменчивость показателя Cn в юго-западной части региона несколько выше, чем на всей остальной территории. Скорее всего, это обусловлено, хотя и слабым, влиянием ели (в результате переноса пыльцы), произрастающей на Урале и в Предуралье, поскольку в западных и особенно юго-западных районах Тюменской области в составе популяций встречаются особи, которые по форме семенных чешуй могут быть отнесены к промежуточной (гибридной) форме ели европейской и сибирской. Внутрипопуляционная изменчивость комплексного показателя формы семенных чешуй (Cn–Cp) составляет около 30%. Дисперсионный анализ (табл. 2) показал относительно большую величину влияния (около 55%) фактора длины шишек и формы семенных чешуй и меньшую – фактора географического положения участка и взаимодействия факторов, хотя все они статистически достоверны (Fф> F0.05).
Таблица 1.
Индивидуальная (внутрипопуляционная) изменчивость показателей длины шишек и формы семенных чешуй ели в сибирской части ареала
| № на рис. 1 | n | Lc | Cn | Cp | Cn–Cp | Корреляция Cn и Cp | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lim | X ± Sx | Cv | Lim | X ± Sx | Cv | Lim | X ± Sx | Cv | Lim | X ± Sx | Cv | η ± Sη | R ± Sr | ||
| 3 | 250 | 50–90 | 71 ± 0.5 | 11 | 42–75 | 60 ± 0.4 | 11 | 34–57 | 45 ± 0.3 | 11 | –17 + 36 | 15 ± 0.6 | 32 | 0.560 ± 0.0526 | 0.525 ± 0.0541 |
| 4 | 235 | 45–90 | 66 ± 0.6 | 14 | 33–79 | 60 ± 0.6 | 14 | 33–62 | 43 ± 0.3 | 11 | –20 + 45 | 17 ± 0.8 | 27 | 0.702 ± 0.0466 | 0.694 ± 0.0472 |
| 9 | 300 | 40–95 | 67 ± 0.5 | 15 | 39–78 | 63 ± 0.4 | 12 | 27–54 | 42 ± 0.2 | 11 | –15 + 50 | 21 ± 0.5 | 29 | 0.567 ± 0.0832 | 0.512 ± 0.0867 |
| 19 | 195 | 35–75 | 51 ± 0.6 | 15 | 45–83 | 68 ± 0.5 | 10 | 30–55 | 39 ± 0.4 | 11 | 0–50 | 29 ± 0.7 | 29 | 0.620 ± 0.0560 | 0.553 ± 0.0600 |
| 21 | 100 | 50–90 | 66 ± 0.7 | 11 | 54–78 | 67 ± 0.5 | 8 | 33–50 | 40 ± 0.4 | 10 | 7–41 | 27 ± 0.8 | 29 | 0.494 ± 0.0878 | 0.463 ± 0.0895 |
| 26 | 150 | 43–90 | 64 ± 0.6 | 12 | 48–78 | 63 ± 0.3 | 11 | 33–54 | 41 ± 0.3 | 10 | 5–45 | 22 ± 0.8 | 45 | 0.584 ± 0.0667 | 0.552 ± 0.0685 |
| 29 | 110 | 45–90 | 68 ± 0.9 | 14 | 48–78 | 64 ± 0.5 | 9 | 33–51 | 41 ± 0.4 | 10 | 5–45 | 23 ± 0.8 | 37 | 0.568 ± 0.0792 | 0.552 ± 0.0802 |
| 32 | 117 | 43–85 | 61 ± 0.9 | 15 | 57–82 | 69 ± 0.5 | 7 | 30–48 | 39 ± 0.4 | 11 | 5–48 | 30 ± 0.8 | 31 | 0.567 ± 0.0768 | 0.502 ± 0.0806 |
| 40 | 150 | 50–90 | 65 ± 0.6 | 12 | 55–82 | 66 ± 0.4 | 7 | 32–53 | 42 ± 0.3 | 9 | 5–50 | 24 ± 0.6 | 31 | 0.578 ± 0.0671 | 0.567 ± 0.0677 |
| 42 | 210 | 35–85 | 62 ± 0.6 | 15 | 55–84 | 69 ± 0.4 | 8 | 30–53 | 41 ± 0.3 | 11 | 2–52 | 28 ± 0.6 | 30 | 0.601 ± 0.0554 | 0.556 ± 0.0576 |
| 43 | 170 | 50–95 | 64 ± 0.7 | 15 | 51–84 | 67 ± 0.4 | 8 | 33–57 | 42 ± 0.3 | 10 | 5–50 | 25 ± 0.7 | 31 | 0.637 ± 0.0595 | 0.595 ± 0.0620 |
| 44 | 130 | 50–95 | 67 ± 0.8 | 14 | 48–80 | 66 ± 0.5 | 8 | 33–57 | 43 ± 0.4 | 11 | –5–42 | 23 ± 0.8 | 30 | 0.568 ± 0.0727 | 0.551 ± 0.0737 |
| 45 | 140 | 45–85 | 64 ± 0.7 | 14 | 51–80 | 64 ± 0.5 | 9 | 36–60 | 43 ± 0.4 | 11 | –14 + 45 | 21 ± 0.8 | 27 | 0.695 ± 0.0612 | 0.603 ± 0.0679 |
| 48 | 130 | 35–65 | 54 ± 0.6 | 13 | 54–82 | 68 ± 0.5 | 8 | 31–50 | 41 ± 0.3 | 10 | 6–50 | 27 ± 0.7 | 28 | 0.352 ± 0.0827 | 0.328 ± 0.0835 |
| 49 | 150 | 45–80 | 62 ± 0.6 | 12 | 54–81 | 66 ± 0.3 | 7 | 31–50 | 41 ± 0.3 | 9 | 7–44 | 25 ± 0.5 | 26 | 0.429 ± 0.0742 | 0.360 ± 0.0767 |
| 50 | 125 | 50–90 | 66 ± 0.8 | 14 | 58–77 | 67 ± 0.4 | 6 | 41–48 | 41 ± 0.3 | 8 | 12–38 | 26 ± 0.7 | 25 | 0.474 ± 0.0794 | 0.389 ± 0.0831 |
Примечание. Здесь и в табл. 3–4: n – число деревьев в выборке; Lc – длина шишек, мм; Cn – коэффициент сужения и Cp – коэффициент вытянутости верхней части семенных чешуй, %; Lim – крайние значения в выборке, X ± Sx – среднее значение и его ошибка, Cv – коэффициент вариации; η ± Sη – корреляционное отношение и его ошибка, R ± Sr – коэффициент корреляции и его ошибка.
Таблица 2.
Результаты двухфакторного дисперсионного анализа популяционно-географической изменчивости показателей длины шишек и формы семенных чешуй ели сибирской в сибирской части ареала
| Дисперсия | Сумма квадратов | Степень свободы | Средний квадрат | Fф | F0.05 |
|---|---|---|---|---|---|
| Общая | 20 655 | 159 | – | – | – |
| Фактор A | 11 340 | 1 | 11 340 | 218.20 | 3.94 |
| Фактор B | 956 | 7 | 136.57 | 2.63 | 2.10 |
| AB | 875 | 7 | 125.07 | 2.41 | 2.10 |
| Остаток | 7484 | 144 | 51.97 | – | – |
Таблица 3.
Географическая изменчивость средних показателей длины шишек и формы семенных чешуй ели в сибирской части ареала
| Признак | Показатель | ||||||
|---|---|---|---|---|---|---|---|
| Lim | X ± Sx | Cv | A ± Sa | t | E ± Se | t | |
| Lc | 50–71 | 64 ± 0.7 | 7.9 | –0.893 ± 0.330 | 2.71 | 0.442 ± 0.622 | 0.71 |
| Cn | 60–70 | 66 ± 0.4 | 4.7 | –0.315 ± 0.328 | 0.96 | –1.295 ± 0.623 | 2.08 |
| Cp | 39–45 | 41 ± 0.2 | 3.7 | 0.606 ± 0.329 | 1.84 | –0.241 ± 0.618 | 0.39 |
| Cn–Cp | 15–31 | 25 ± 0.6 | 17.0 | –0.476 ± 0.428 | 1.45 | –0.817 ± 0.624 | 1.31 |
Примечание. A ± Sa – показатель асимметрии и его ошибка, E ± Se – показатель эксцесса и его ошибка, t – показатель достоверности (t0.05 = 2.01). Остальные обозначения см. табл. 1.
Таблица 4.
Географическая корреляция средних показателей длины шишек и формы семенных чешуй ели в сибирской части ареала
| Коррелирующие признаки | η ± Sη | tη | R ± Sr | tr | Kk |
|---|---|---|---|---|---|
| Cn и Cp | 0.726 ± 0.0993 | 7.31 | –0.693 ± 0.1040 | 6.66 | 0.046 |
| Cn и Lc | 0.544 ± 0.1212 | 4.49 | –0.525 ± 0.1228 | 4.27 | 0.020 |
| Cp и Lc | 0.448 ± 0.1290 | 3.47 | +0.426 ± 0.1306 | 3.26 | 0.019 |
| Cn–Cp и Lc | 0.543 ± 0.1212 | 4.48 | –0.530 ± 0.1224 | 4.33 | 0.015 |
Примечание. Kk – показатель криволинейности связи (η2–R2). Остальные обозначения см. табл. 1.
Определенный интерес представляет изучение характера и уровня связи показателей Cn и Cp в пределах популяций. Оказалось, что корреляционное отношение и коэффициент отрицательной корреляции этих показателей во всех случаях достоверны и мало различаются по своей величине: в среднем корреляционное отношение равно 0.562, а коэффициент корреляции составляет –0.519. Близость числовых значений этих коэффициентов указывает на прямолинейность связи, поскольку величина коэффициента криволинейности связи недостоверна (η2–R 2 = = 0.046 < 0.1). Таким образом, чем больше коэффициент сужения чешуй (Cn), тем меньше “вытянутость” их верхней части (Cp), и наоборот. Поскольку корреляция показателей только среднего уровня, имеются отклонения от этой закономерности. В целом же уровень и характер связи показателей Cn и Cp такой же, как в популяциях ели на европейской части ареала (Попов, 2005). Связь показателей формы семенных чешуй с длиной шишек в пределах популяций отсутствует (Попов, 2012).
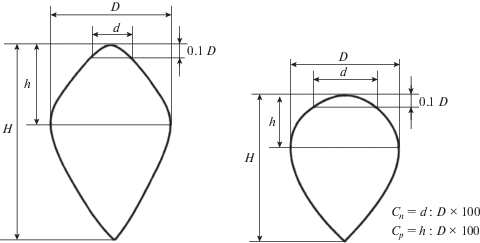
Географическая изменчивость средних показателей длины шишек (рис. 3) и формы семенных чешуй (Cn, Cp, Cn–Cp) определена по всем 50 выборкам. С точки зрения общей географической изменчивости признака елей европейской и сибирской эти шишки ближе к мелким (Попов, 2011):
Рис. 3.
Географическая изменчивость средней длины шишек ели на территории Сибири в пунктах сбора материала (см. рис. 1).
В большей части выборок (42 шт.) длина шишек составляет 60–71 мм и только в 8 выборках она равна 50–59 мм. Во всем ряду выборок среднее значение их 64.2 ± 0.71 мм, коэффициент вариации – 7.9%, медиана – 65 мм, коэффициент асимметрии –0.893 ± 0.3298 при t = 2.71; коэффициент эксцесса – 0.442 ± 0.0.6225 при t = 0.71 (табл. 3). Наименьшая длина шишек характерна для популяций, располагающихся в основном вдоль Северного полярного круга. Вся остальная территория занята популяциями ели с длиной шишек, равной 60–70 мм.
Примерно в тех же пределах изменчивость признака определена Г.М. Говориным (1992) в бассейне Енисея. Аналогичные оценки длины шишек ели на восточном пределе ареала ели сибирской даны в работах А.М. Науменко (1964), С.А. Андреева (1986), О.Н. Арбузовой (1994). На северном пределе распространения ели в Восточной Сибири (от Туруханска до Верхоянска) средняя длина шишек равна 50–51 мм, а в центральной части ареала (Алтай, Томск, Иркутская область, Бурятия) она оказалась значительно больше (76–79 мм), чем в других районах Сибири (Коропачинский и др., 2012). В целом же нет четкой зависимости изменения средней длины шишек в направлении север–юг, как на европейской части ареала (Попов, Александров, 2010; Попов, 2011). Шишки длиной 60–70 мм в относительно благоприятных условиях произрастания ели содержат примерно 80–100 полных семян (Попов, 1996). Но на северном пределе распространения ели в Сибири шишки (несколько меньшей длины) на большей части деревьев содержат пустые семена, а полные характеризуются пониженной и очень низкой всхожестью (Норин, 1958; Попов и др., 2015в; Арефьев и др., 2016).
Среднее значение показателя Cn равно 66 (60–70), показателя Cp – 41 (39–45), их разности (Cn–Cp) – 25 (15–31%). В популяциях ели европейской показатель Cn (25–40%) больше показателя Cp (70–80%), а их разность (–55…–40%) имеет отрицательный знак (Попов, 2005). Соответствующие коэффициенты географической (межпопуляционной) вариации составляет 4.7, 3.7, 17.0%. Для всей территории бывшего СССР эти показатели равны 26, 20.5, 40%. Характер распределений анализируемых признаков в целом близок к нормальному. Здесь наблюдается достоверная отрицательная величина эксцесса (–1.295 ± 0.623; t = = 2.08) только по показателю Cn, поскольку во всем ряду не оказалось ни одной выборки с величиной признака, равной 65%, разделяющей ряд на две примерно равные части. Но это явление, скорее всего, случайное. Отсутствие достаточно четких отклонений величины эксцесса по анализируемым признакам (Lc, Cn, Cp, Cn–Cp) от нормального распределения свидетельствует об относительно большой географической однородности популяций ели в регионе. Хотя величина достоверных различий средних значений в выборках составляет по длине шишек около 2 мм, по показателям формы семенных чешуй – 2%. Достоверные различия некоторых выборок по длине шишек и форме семенных чешуй, вероятно, обусловлены особенностями популяций, связанными со значительным расстоянием между участками их расположения.
Степень сходства в географической изменчивости анализируемых показателей на сибирской части ареала характеризуется средним уровнем их прямолинейной корреляции (табл. 4). Например, уравнения связи среднего показателя длины шишек и комплексного показателя формы семенных чешуй (Cn–Cp) имеют вид:
Ошибка уравнений 1.17. Расчетные значения признаков составляют примерно следующие величины, которые в целом соответствуют показателям их географической изменчивости:
Корреляция эмпирических и расчетных величин Lc (η = 0.670 ± 0.1071; R = 0.537 ± 0.1217) и Cn–Cp (η = 0.550 ± 0.1205; R = 0.546 ± 0.1209) имеет тот же уровень. Корреляция по Lc достоверно отличается от прямолинейной.
В связи со слабой географической зависимостью анализируемых признаков не обнаруживается и их достаточно четкой соответствующей динамики. Только по показателю Cn–Cp при градации в 5–6% (15–20; 21–25; 26–30%) видна достаточно ясная картина его географического изменения в Западной Сибири (Попов и др., 2015б). В данном случае популяции с меньшими показателями Cn–Cp располагается также в самой юго-западной части региона, с наибольшими – в северной. Сюда же относятся выборки из района Горно-Алтайска (Cn–Cp = 30%) и Забайкалья (Cn–Cp = 27%). Вся остальная территория занята популяциями с промежуточными значениями этого показателя (21–25%).
Поскольку в географической изменчивости длины шишек и показателей формы семенных чешуй наблюдается большое сходство, то попробуем разделить всю совокупность популяций на группы по классам формы семенных чешуй и длины шишек. При этом установлено, что в группе популяций с наибольшей заостренностью (угловатостью) чешуй оказываются только “крупные” шишки (64 мм и более). В группе с несколько меньшей угловатостью чешуй – преимущественно такие же шишки и только в двух выборках шишки класса меньшей длины. Группа популяций ели с наибольшей притупленностью семенных чешуй (26% и более) “содержит” популяции со всеми классами шишек. Средние показатели длины шишек по классам формы семенных чешуй (Cn–Cp) в значительной степени отражают особенности географической дифференциации популяций ели на территории Сибири. Группы популяций со средней и большой величиной показателя Cn–Cp несколько ближе друг к другу, чем группы со средней и меньшей величиной шишек.
Особенности географической изменчивости и дифференциации популяций ели в Сибири можно объяснить отчасти условиями произрастания и характером ее расселения в прошлом. Такой характер распространения вида, скорее всего, остаточный от предшествующего периода, когда он был распространен более широко. Ель, произрастая часто на небольших участках, иногда среди заболоченных пространств, “эволюционирует” как бы сама в себе без внешнего притока генов и формирует из поколения в поколение признаки почти одинаковых параметров, часто отличающихся от европейских популяций ели (Schmidt-Vogt, 1977; Borchetti et al., 1988).
Заключение. Ель сибирская на большей части Сибири в современный период имеет сравнительно короткие шишки (50–70, иногда до 80 мм), во внутрипопуляционной изменчивости амплитуда признака несколько больше (45–90 мм). Географическая изменчивость средних значений коэффициента сужения (Cn) небольшая и находится в пределах 60–70%, коэффициента вытянутости чешуй – 39–45%. Показатель разности их в среднем составляет около 25% при довольно высокой внутрипопуляционной изменчивости и значительно меньшей – географической. Результаты исследования могут быть полезны для решения вопросов популяционно-географической дифференциации ели сибирской с точки зрения лесоводства.
Список литературы
Андреев С.А. Ель сибирская в верховьях р. Гилюй // Лесоведение. 1986. № 2. С. 80–86.
Арбузова О.Н. Picea и Larix (Pinaceae) в плейстоценовых флорах района реки Берелех (Бассейн реки Колымы) // Ботанический журн. 1994. Т. 79. № 5. С. 1–10.
Арефьев С.П., Казанцева М.Н., Попов П.П. Некоторые особенности в изменчивости семян ели сибирской в северных районах ареала (Ямало-Ненецкий автономный округ) // Вестник Тюменского гос. университета. Экология и природопользование. 2016. Т. 2. № 4. С. 96–107.
Атлас лесов СССР. М.: ГУГК, 1973. 222 с.
Говорин Г.М. Изменчивость ели сибирской в бассейне Енисея // Лесоведение. 1992. № 5. С. 56–60.
Гончаренко Г.Г., Падутов В.Е. Популяционная и эволюционная генетика елей Палеарктики. Гомель: Институт леса Национальной АН Беларуси, 2001. 197 с.
Кеппен Ф. Географическое распространение хвойных деревьев в Европейской России и на Кавказе. СПб.: Типография Императорской Академии наук, 1885. 634 с.
Коропачинский И.Ю., Потемкин О.Н., Рудиковский А.В., Кузнецова Е.В. Полиморфизм и структура популяций ели сибирской (Picea obovata Ledeb.) на северном пределе распространения вида // Сибирский экологический журн. 2012. № 2. С. 175–184.
Кравченко А.Н., Ларионова А.Я., Милютин Л.И. Генетический полиморфизм ели сибирской (Picea obovata Ledeb) в Средней Сибири // Генетика. 2008. Т. 44. № 1. С. 45–53.
Кравченко А.Н., Ларионова А.Я., Экарт А.К. Генетический полиморфизм ели сибирской Picea obovata (Pinaceae) в азиатской части ареала // Вестник Северо-Восточного НЦ ДВО РАН. 2013. № 2. С. 74–85.
Круклис М.В. Кариологические особенности Picea obovata Ledeb. // Лесоведение. 1971. № 2. С. 76–84.
Мамаев С.А. Формы внутривидовой изменчивости древесных растений. М.: Наука, 1972. 284 с.
Милютин Л.И. О таксономическом статусе и внутривидовой изменчивости ели сибирской (Picea obovata) // Ботанический журн. 2015. Т. 100. № 1. С. 33–38.
Муратова Е.Н., Владимирова О.С. Добавочные хромосомы кариотипа ели сибирской P. obovata // Цитология и генетика. 2001. № 4. С. 38–44.
Науменко А.М. Picea obovata Ledeb. на крайнем северо-восточном пределе своего ареала // Ботанический журн. 1964. Т. 49. № 7. С. 1008–1013.
Норин Б.Н. К познанию семенного и вегетативного возобновления древесных пород в лесотундре // Растительность Крайнего Севера и ее освоение. М.; Л.: Изд-во АН СССР, 1958. С. 154–244.
Попов П.П. Предварительная оценка выхода семян из шишек ели // Лесное хозяйство. 1996. № 6. С. 35–36.
Попов П.П. Ель на востоке Европы и в Западной Сибири: Популяционно-географическая изменчивость и ее лесоводственное значение. Новосибирск: Наука, 1999. 169 с.
Попов П.П. Ель европейская и сибирская: Структура, интерградация и дифференциация популяционных систем. Новосибирск: Наука, 2005. 231 с.
Попов П.П. Популяционно-географическая изменчивость шишек ели европейской и сибирской // Лесоведение. 2011. № 5. С. 54–60.
Попов П.П. Соотношение показателей длины шишек и формы семенных чешуй в популяциях Picea abies и P. obovata (Pinaceae) // Ботанический журнал. 2012. Т. 97. № 6. С. 729–734.
Попов П.П., Александров А.Х. Изменчивость длины шишек и формы семенных чешуй ели европейской в зависимости от географической широты // Лесоведение. 2010. № 4. С. 66–73.
Попов П., Александров А., Недкова Е. Географическая изменчивость и фенотипическая структура популяций ели обыкновенной по форме семенных чешуй // Наука за гората (Forest science). 2015а. Кн. 2. С. 1–15.
Попов П.П., Арефьев С.П., Гашева Н.А., Казанцева М.Н. Географическая изменчивость и фенотипическая структура популяций Picea obovata (Pinaceae) в Западной Сибири // Ботанический журн. 2015б. Т. 100. № 9. С. 927–938.
Попов П.П., Арефьев С.П., Гашева Н.А., Казанцева М.Н. Качество семян Picea obovata (Pinaceae) на северной границе ареала (Ямало-Ненецкий автономный округ) // Растительные ресурсы. 2015в. Т. 51. № 4. С. 512–519.
Соколов С.Я., Связева О.С., Кубли В.А. Ареалы деревьев и кустарников СССР. Л.: Наука, 1977. 163 с.
Сукачев В.Н. Дендрология с основами лесной геоботаники. Л.: Гослестехиздат, 1938. 576 с.
Теплоухов Ф.А. Известия о деятельности Лесного общества (Сообщение о коллекции еловых шишек в Лесном отделе Политехнической выставки) // Лесной журн. 1872. Вып. 6. С. 86–91.
Цепляев В.П. Леса СССР. М.: Сельхозиздат, 1961. 216 с.
Borchetti M., Giannini R. and Menozzi P. Geographic Variation in Cones of Norway Spruce (Picea abies (L.) Karst.) // Silvae genetica. 1988. V. 37. № 5–6. P. 178–184.
Schmidt-Vogt H. Die Fichte. Taxonomie. Verbreitung. Morphologie. Waldgesellschaften. Hamburg; Berlin: Paul Parey, 1977. V. 1. 647 c.
Teplouchoff Th. Ein Beitrag zur Kenntniss der sibirischen Fichte Picea obovata Ledeb // Bulletin de la Societe Imperiale des Naturalistes de Moscou. 1868. V. 41. № 3. C. 244−252.
Дополнительные материалы отсутствуют.