

Лесоведение, 2019, № 3, стр. 188-197
Изменчивость показателей плодов и однолетних сеянцев редкого вида каркас кавказский
Б. М. Магомедова *
Горный ботанический сад Дагестанского НЦ РАН
367000 Махачкала, ул. М. Гаджиева, 45,
Россия
* E-mail: bary_m@mail.ru
Поступила в редакцию 24.03.2017
После доработки 30.01.2018
Принята к публикации 03.12.2018
Аннотация
Одним из редких и уязвимых древесных растений, занесенных в Красную книгу Дагестана, является каркас кавказский Celtis caucasica Willd. Проведено выявление лимитирующих факторов уменьшения численности данного вида, изучена внутривидовая изменчивость природной популяции, исследовано влияние экологических условий на начальные этапы онтогенеза каркаса кавказского на примере группы особей, произрастающих в Кайтагском районе на окраине с. Маджалис (42°08′49.6″ с.ш., 47°53′26.2″ в.д., 315 м над ур. моря). Жизненность особей высокая, плодоношение обильное, возраст около 30–35 лет, средняя высота деревьев 6.5–7.5 м. Особи находятся в средневозрастном генеративном состоянии, сеянцев, ювенильных и сенильных особей не обнаружено. Полиморфизм параметров листа характеризуется средним уровнем изменчивости. Среди рассматриваемых признаков плода очень низким уровнем изменчивости характеризуются: длина плода (5.6%), ширина плода (7.4%), индекс плода (6.7%), длина косточки (6.1%), ширина косточки (7%), как генетически более стабильные. Изучение коррелятивных связей между анализируемыми признаками позволило выявить высокую положительную корреляцию по признакам плода и косточки, по длине и ширине листа. Лабораторная всхожесть семян C. caucasica составила 81.7%. Темпы роста сеянцев в более благоприятных климатических условиях горно-долинного высотного пояса (1100 м над ур. моря) реализовываются лучше, чем в условиях Гунибского плато (1750 м над ур. моря) и Махачкалы (100 м над ур. моря), что подтверждено и биометрическими показателями развития побеговых систем. В результате проведенной работы установлено, что основной причиной низкой численности C. caucasica в Дагестане на сегодняшний день, видимо, является низкая всхожесть семян в естественных условиях, которая получила экспериментальное подтверждение и отсутствие семенного возобновления в популяции.
Сохранение редких, исчезающих и требующих охраны видов растений является частью глобальной проблемы изучения и сохранения биоразнообразия. Охрана генетического фонда флоры, особенно региональной, имеет исключительно важное значение. Региональные флоры в большинстве случаев являются носителями информации о своей истории в минувшие геологические эпохи, и полное сохранение их фиторазнообразия имеет большое теоретическое и практическое значение. Однако изученность многих редких видов еще недостаточна и не обеспечивает решения задач по их интродукции и охране. При изучении популяций редких видов большой интерес представляют данные по изменчивости морфологических признаков, поскольку наличие внутрипопуляционной изменчивости свидетельствует о разной степени приспособляемости вида к различным экологическим условиям и его большой пластичности (Князева, 2007; Магомедова и др., 2016). Необходимы данные по численности и состоянию популяций, характеру и эффективности процессов возобновления редких видов растений. Изучение экологических условий произрастания, состояния ценопопуляций послужит основой для интродукции редких видов в условиях ботанических садов.
Одним из редких и уязвимых древесных видов, занесенных в Красную книгу Республики Дагестан, является Celtis caucasica Willd. – каркас кавказский. Он встречается единичными экземплярами в полосе аридных предгорных редколесий, чистых монодоминантных зарослей не образует. На Кавказе произрастает в нижнем и среднем горном поясах, приблизительно до 1500–1600 м над ур. моря, редко выше; общий ареал – Кавказ, Юго-западная и Средняя Азия. Исключительная засухоустойчивость этого вида объясняется чрезвычайно мощной развитой корневой системой. Ценится как дерево для эффективного облесения и, в особенности закрепления склонов в сухих местностях с малоплодородными почвами (в частности, участков с осыпями). Совместно с другими видами деревьев пригоден для создания полезащитных (ветроломных) насаждений в засушливых районах. Благодаря сильной облиственности кроны, красивой коре, отмеченным выше засухоустойчивости и жаростойкости, а также долговечности, очень ценен для озеленения (Гулисашвили, 1961).
Литературные сведения о виде немногочисленны, посвящены в основном распространению и морфологическому описанию (Гроссгейм, 1945; Гулисашвили, 1961; Галушко, 1967; Абдурахманов, 2009).
Целью данной работы является выявление лимитирующих факторов уменьшения численности C. caucasica, изучение внутривидовой изменчивости природной популяции, исследование влияния экологических условий на начальные этапы онтогенеза изучаемого вида.
ОБЪЕКТЫ И МЕТОДИКА
Изучение внутрипопуляционной изменчивости Celtis caucasica проводилось в Кайтагском районе Республики Дагестан (в окрестностях с. Маджалис 42°08′49.6″ с.ш., 47°53′26.2″ в.д., 315 м над ур. моря). В первой декаде сентября в период массового созревания плодов было собрано по 5 однолетних побегов с 10 деревьев. Учитывались следующие признаки: число генеративных метамеров, число вегетативных метамеров, длина листа, ширина листа, индекс листа, число зубцов по краю листа, длина побега, длина плода, ширина плода, индекс плода, длина плодоножки, вес плода, вес косточки, длина косточки, ширина косточки. Индекс плода был вычислен путем деления длины на ширину плода, индекс листа – делением длины на ширину листа. Морфологические признаки изучали путем элементарных измерений. Весовые признаки измерялись на электронных весах “Ohaus” с точностью до 1 мг, размерные – штангенциркулем с точностью до 0.1 мм. Динамика роста и развития однолетних сеянцев изучалась в условиях Горного Ботанического сада Дагестанского НЦ РАН. По результатам измерений проводился статистический анализ.
Посев семян производили по общепринятым методикам (Николаева и др., 1985). Фенологические наблюдения были проведены в соответствии с методикой (Иваненко, 1962; Александрова и др., 1975). Ежемесячно с момента появления всходов линейкой измеряли длину годичного побега. Для оценки зимостойкости сеянцев применена пятибалльная шкала, адаптированная нами применительно к изучаемому виду, с использованием подходов, изложенных в коллективной монографии “Древесные растения Главного ботанического сада АН СССР” (1975): 1 – растение не обмерзает; 2 – обмерзает до 50% длины побега; 3 – обмерзает более 50% длины побега; 4 – обмерзает вся надземная часть (до корневой шейки); 5 – растения вымерзают целиком.
Посев в открытый грунт производили на трех высотных уровнях (Гунибская и Цудахарская экспериментальные базы Горного ботанического сада, г. Махачкала) в 2 срока: осенью, без предпосевной обработки, и весной, после 3-х месячной стратификации.
Опытный участок в г. Махачкале относится к Низменному Дагестану и расположен на высоте 100 м над ур. моря, между 42°58′51″ с.ш. и 47°22′04″ в.д. Территория сложена песчано–глинистыми толщами морских каспийских осадочных пород. Снеговой покров маломощный, продолжительность залегания снега 20–50 дней. В холодные зимы морозы достигают 20–30°С. Ветры преобладают юго-восточные и северо-западные. Абсолютный минимум – минус 25°, число дней со снеговым покровом 16–25. Среднее количество осадков – 328 мм, средняя годовая температура – 12.25°С. Относительная влажность воздуха – 75%.
Гунибская экспериментальная база Горного ботанического сада находится в Гунибском районе Республики Дагестан, в 3 км к западу от с. Гуниб, на Гунибском плато (плато Большого Кавказа), на высоте 1650–2000 м над уровнем моря. Координаты: 42°24′34″ с.ш., 46°55′75″ в.д., 1630–1980 м над ур. моря. Плато Гуниб находится в 150 км к юго-западу от административного центра Дагестана – Махачкалы. По рельефу и геологическому строению относится к известняковому району, образованному преимущественно меловыми и юрскими известняками. Климат – континентальный, среднегодовая сумма осадков – 680 мм, с летним максимумом. Средняя годовая температура +6.6°, абсолютный минимум –26.0°, абсолютный максимум +36.0°. Относительная влажность воздуха – 65%. В почвенном покрове преобладают субальпийские горно-луговые и черноземовидные почвы (Акаев, 1996).
Цудахарская экспериментальная база Горного ботанического сада находится в Левашинском районе Республики Дагестан, в 2 км к югу от с. Цудахар, на высоте 900–1100 м над ур. моря. Координаты: 42°19′39″ с.ш., 47°09′53″ в.д. Среднее количество осадков – 440 мм, средняя годовая температура – 6.9°С. Относительная влажность воздуха – 72%. В почвенном покрове преобладают известковые почвы.
Статистическая обработка полученных результатов выполнена с использованием программ Excel, графики – с помощью лицензионной программы Statistica V. 5.5.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Популяция Celtis caucasica, произрастающая в Кайтагском районе Республики Дагестан не образует монодоминантных зарослей. Древесные растения образуют 2 яруса, всего на описываемых участках, помимо Сeltis caucasica произрастает еще 21 вид деревьев и кустарников: Crataegus pentagyna, Ulmus minor, Rhamnus cathartica, Cornus mas, Paliurus spina-christi, Malus orientalis, Mespilus germanica, Quercus robur, Rosa canina, Berberis vulgaris, Acer ibericum, Acer campestre, Prunus divaricata, Ulmus minor, Viburnum lantana, Fraxinus excelsior, Cydonia oblonga, Ligustrum vulgare, Euonymus verrucosus, Rubus sp., Swida australis. Доминирующим видом из древесных растений первого яруса является Crataegus pentagyna. Во втором ярусе численно преобладают Ligustrum vulgare, Rosa canina, Viburnum lantana, подрост Fraxinus excelsior и Crataegus pentagyna. Жизненность особей высокая, плодоношение обильное, средний возраст деревьев составляет 30–35 лет, высота 6.5–7.5 м. Особи находятся в средневозрастном генеративном состоянии, сеянцев, ювенильных и сенильных особей не обнаружено.
Годичные побеги относятся к фундаментальной категории признаков жизненных форм растений, т.е. основная их функция – ассимиляция и репродукция. Соподчиненной единицей такого модуля является лист (Хохряков, Мазуренко, 1993).
Лист, обладая определенной пластичностью в отношении численности, размера, времени и места возникновения, наилучшим образом облегчает сохранение эффективной взаимосвязи между растением и окружающей средой (Романова, Ковригина, 2008). Размеры листовой пластинки отражают мощность развития растений.
Проведенные исследования внутрипопуляционной изменчивости элементов генеративного побега редкого вида C. caucasica показывают, что в объединенной выборке такие признаки, как длина листа (16%), ширина лист” (19.3%), индекс листа (13%), а также длина побега (16.7%) характеризуются средним уровнем изменчивости. Высокий уровень изменчивости наблюдается по числу вегетативных (32.2%) и генеративных метамеров (36.3%) (табл. 1).
Таблица 1.
Статистические параметры признаков побега и плодов Celtis caucasica
| Признак | Значения | |
|---|---|---|
| x ± Sx | C.V., % | |
| Число вегетативных метамеров, шт. | 3.2 ± 0.15 | 32.2 |
| Число генеративных метамеров, шт. | 2.9 ± 0.14 | 36.3 |
| Длина листа, см | 5.5 ± 0.12 | 16.0 |
| Ширина листа, см | 2.5 ± 0.06 | 19.3 |
| Индекс листа | 2.3 ± 0.04 | 13.0 |
| Число зубцов, шт. | 17.2 ± 0.31 | 13.2 |
| Длина побега, см | 10.7 ± 0.24 | 16.7 |
| Длина плода, см | 0.86 ± 0.007 | 5.6 |
| Ширина плода, см | 0.78 ± 0.008 | 7.4 |
| Индекс плода | 1.11 ± 0.009 | 6.7 |
| Длина плодоножки, см | 1.3 ± 0.03 | 18.2 |
| Вес плода, г | 0.25 ± 0.007 | 20.4 |
| Вес косточки, г | 0.14 ± 0.003 | 17.6 |
| Длина косточки, см | 0.62 ± 0.005 | 6.1 |
| Ширина косточки, см | 0.54 ± 0.005 | 7.0 |
Примечание. В табл. 1, 3–5 (x ± Sx) – среднее значение и стандартное отклонение, C.V. – коэффициент вариации.
Повышенным уровнем изменчивости среди изученных признаков плодов C. caucasica обладает вес плода 20.4%. Средний уровень изменчивости имеют такие признаки, как длина плодоножки (18.2%), вес косточки (17.6%) (табл. 1). Очень низким уровнем изменчивости характеризуются длина плода (5.6%), ширина плода (7.4%), индекс плода (6.7%), длина косточки (6.1%), ширина косточки (7%) как генетически более стабильные.
Значение индекса плода, определяющее форму плода каркаса кавказского в природной популяции, составляет 1.1, что говорит о слегка вытянутой или почти округлой его форме.
Для определения силы связей между признаками побега применен корреляционный анализ, который выявил статистически достоверную корреляционную зависимость между некоторыми парами признаков (табл. 2).
Таблица 2.
Коэффициенты корреляции для признаков побега Celtis caucasica
| Признак | Число вегетативных метамеров | Число генеративных метамеров | Длина листа | Ширина листа | Индекс листа | Число зубцов | Длина побега | Длина плода | Ширина плода | Индекс плода | Длина плодоножки | Вес плода | Вес косточки | Длина косточки |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Число генеративных метамеров | –0.65* | |||||||||||||
| Длина листа | 0.38* | –0.14 | ||||||||||||
| Ширина листа | 0.32* | 0.01 | 0.75* | |||||||||||
| Индекс листа | –0.06 | –0.16 | 0.03 | –0.59* | ||||||||||
| Число зубцов | 0.26 | –0.04 | –0.01 | 0.03 | –0.05 | |||||||||
| Длина побега | 0.12 | 0.45* | 0.48* | 0.67* | –0.44* | 0.14 | ||||||||
| Длина плода | 0.25 | –0.04 | 0.36* | 0.56* | –0.45* | 0.01 | 0.41* | |||||||
| Ширина плода | 0.16 | 0.12 | 0.20 | 0.38* | –0.33* | 0.02 | 0.48* | 0.76* | ||||||
| Индекс плода | 0.04 | –0.28* | 0.07 | 0.01 | 0.04 | –0.02 | –0.33* | –0.11 | –0.53* | |||||
| Длина плодоножки | 0.41* | –0.27 | 0.59* | 0.48* | –0.03 | 0.11 | 0.21 | 0.23 | –0.03 | 0.06 | ||||
| Вес плода | 0.32* | –0.07 | 0.32* | 0.48* | –0.36* | 0.08 | 0.57* | 0.66* | 0.77* | –0.31* | 0.04 | |||
| Вес косточки | 0.16 | 0.08 | 0.33* | 0.54* | –0.41* | 0.00 | 0.66* | 0.68* | 0.76* | –0.26 | 0.04 | 0.88* | ||
| Длина косточки | 0.24 | –0.11 | 0.34* | 0.49* | –0.29* | 0.04 | 0.38* | 0.77* | 0.68* | –0.15 | 0.09 | 0.65* | 0.69* | |
| Ширина косточки | 0.01 | 0.29* | 0.19 | 0.41* | –0.41* | –0.20 | 0.60* | 0.58* | 0.65* | –0.26 | –0.06 | 0.60* | 0.80* | 0.56* |
Обнаружена положительная корреляция между весовыми и линейными признаками плода и косточки. Значительная корреляционная зависимость наблюдается между всеми признаками побега с длиной и шириной листа и по признакам плода. Так, для признаков “вес косточки” и “вес плода” (0.88); “длина листа” и “ширина листа” (0.75); “длина косточки” и “длина плода” (0.77); “вес плода” и “ширина плода” (0.77); “вес косточки” и “ширина плода” (0.76); “ширина косточки” и “вес косточки” (0.80); “ширина плода” и “длина плода” (0.76) наблюдается высокая достоверная положительная корреляционная связь. Положительная заметная достоверная связь имеется между признаками “длина плодоножки” и “длина листа” (0.59); “длина побега” и “ширина листа” (0.67); “длина плода” и “ширина листа” (0.56); “вес косточки” и “ширина листа” (0.54); “вес плода” и “длина побега” (0.56); “вес косточки” и “длина побега” (0.66); “ширина косточки” и “длина побега” (0.60); “вес плода и длина плода” (0.66); “вес косточки” и “длина плода” (0.68); “ширина косточки” и “длина плода” (0.58); “длина косточки и вес плода” (0.65); “ширина косточки” и “вес плода” (0.60); “длина косточки” и “вес косточки” (0.69); “ширина косточки” и “длина косточки” (0.56).
Таким образом, изучение коррелятивных связей между анализируемыми признаками позволило выявить положительную корреляцию по признакам плода.
Выявленные недостоверные положительные связи показывают наличие определенной автономности в реализации органами морфогенетических потенций.
Достоверная отрицательная связь существует между признаками числом генеративных и вегетативных метамеров (–0.65), индексом и шириной плода (–0.53), индексом и шириной листа (‒0.59). Отрицательные достоверные связи, на наш взгляд, выражают степень перераспределения энергопластических веществ между достаточно сильно сопряженными элементами конструкции побега. В некоторых случаях отрицательная корреляционная связь возникает между размером и числом частей в результате компенсационного роста. В других случаях они могут быть связаны с возрастом растения, так как известно, что на более поздних стадиях онтогенеза возрастает роль экзогенной регуляции: под влиянием условий внешней среды система корреляций несколько “расшатывается”, могут изменяться или исчезать старые и возникать новые связи между признаками, и наоборот, первые этапы онтогенеза весьма жестко регулируются внутренними факторами и уровень корреляции возникающих признаков очень высок (Шмидт, 1985). Этим, вероятно, объясняется обнаруженное высокое число положительных достоверных связей.
В естественных условиях нами не выявлено семенного возобновления в популяции, в связи с чем проведены опыты по изучению всхожести семян и изучены начальные этапы онтогенеза C. caucasica в различных экологических условиях.
Осенний посев семян C. caucasica показал низкую всхожесть (42%), что может быть связано с несоответствием экологического оптимума прохождения глубокого покоя семян, ходом зимней температуры в природных условиях и водным балансом. Весенний посев стратифицированных семян показал более высокие результаты (60%), т.е. предпосевная обработка способствует лучшему проявлению их биологического потенциала. Лабораторная всхожесть составила 81.7%. Прорастание у семян C. caucasica надземное, гипокотиль (30–50 мм) более развитый, чем эпикотиль (15–18 мм). Семядольные листья (длина 10–12 мм) почти сидячие, прямоугольные, на верхушке с широкой выемкой шириной 7–10 мм, на очень коротких черешках, почти сидячие. У семядолей хорошо выражено жилкование.
Сравнительная характеристика динамики роста сеянцев вида C. caucasica в условиях Дагестана на трех высотных уровнях (Махачкала, Цудахарская и Гунибская экспериментальные базы) представлена в табл. 3.
Таблица 3.
Динамика прироста сеянцев Celtis caucasica в разных экологических условиях
| Место произрастания | Длина побега, см | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| май | июнь | июль | август | сентябрь | ||||||
| x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | |
| Махачкала | 6.7 ± 0.36 | 17.6 | 8.7 ± 0.73 | 28.5 | 17.9 ± 1.31 | 21.8 | 23.2 ± 1.25 | 16.9 | 31.3 ± 2.46 | 23.8 |
| Цудахарская база | 2.3 ± 0.25 | 12.5 | 6.8 ± 0.27 | 21.6 | 30.6 ± 1.15 | 11.6 | 45.6 ± 2.45 | 11.8 | 65.3 ± 3.45 | 17.8 |
| Гунибская база | 1.5 ± 0.08 | 6.7 | 1.9 ± 0.05 | 9.8 | 7.3 ± 0.43 | 8.4 | 11.5 ± 0.73 | 15.4 | 21.9 ± 1.32 | 14.8 |
Примечание. Обозначения см. табл. 1.
Как видно из табл. 3, на низменности (100 м над ур. моря) наблюдается постепенное увеличение биометрических показателей развития побеговой системы, к концу сентября средняя длина побега составила 31.3 см. В условиях горно-долинного (Цудахарская база, 1100 м над ур. моря) и среднегорного (Гунибская база, 1750 м над ур. моря) высотных поясов темпы роста сеянцев в первый месяц вегетации низкие; в более благоприятных климатических условиях горно-долинного пояса в последующем они реализовываются лучше, чем в условиях Гунибского плато, что подтверждено биометрическими показателями развития побеговых систем (табл. 3). Интенсивность ростовых процессов сеянцев была высокой с июля до наступления осенних заморозков.
Таблица 4.
Сравнительная характеристика изменчивости морфологических признаков листа однолетних сеянцев Celtis caucasica вдоль высотного градиента*
| Высота над ур. моря |
Длина черешка (C), мм | Длина листовой пластинки (А), мм | Общая длина листа (AC), мм | Максимальная ширина пластинки (В), мм | Индекс формы листа (В)/(А), % | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | |
| 100 | 5.8 ± 0.54 | 19.5 | 56.2 ± 2.45 | 12.8 | 62.0 ± 2.56 | 12.0 | 33.2 ± 1.25 | 9.3 | 59.1 ± 1.23 | 6.7 |
| 1100 | 6.9 ± 0.27 | 16.5 | 89.7 ± 2.17 | 8.5 | 96.6 ± 2.89 | 6.5 | 55.3 ± 2.19 | 12.8 | 61.6 ± 1.34 | 7.5 |
| 1700 | 4.1 ± 0.39 | 15.1 | 55.8 ± 2.30 | 8.9 | 59.9 ± 2.45 | 7.9 | 35.3 ± 1.87 | 11.6 | 63.3 ± 1.15 | 4.3 |
* Выборка n = 10. Примечание. Обозначения см. табл. 1.
Относительная изменчивость показателей динамики роста характеризовалась низкими и средними значениями в периоды незначительного прироста (май, август) до высокой и весьма высокой в периоды интенсивного роста (июнь, июль и сентябрь). Наши исследования показали, что выращивание сеянцев вида C. caucasica в разновысотных условиях Дагестана выявляет многовариативные программы развития растений при воздействии экзогенных и эндогенных факторов.
Основной причиной низкой численности C. caucasica в Дагестане на сегодняшний день является низкая всхожесть семян в естественных условиях, что было подтверждено нами экспериментально. Иными словами, высокий биологический потенциал семян этого растения недостаточно полно реализуется в природных условиях Предгорного Дагестана, что может быть следствием недостаточной экологической пластичности растений на первых этапах жизненного цикла.
Для древесных растений-интродуцентов важным условием адаптации является поддержание определенной “физиологической нормы” ассимиляционных структур. В то же время вопрос о том, насколько широки границы изменений, не выходящих за пределы нормы реакций, заслуживает изучения для каждого отдельно взятого вида (Кавеленова и др., 2008). Изучение изменчивости морфологических признаков листа традиционно считается важнейшей составной частью эколого-морфологических исследований в экологии видов. Морфометрия листа является дополнительным информативным признаком в изучении пластичности видов вдоль средовых градиентов. В связи с этим нами было проведено исследование основных закономерностей изменчивости морфологических признаков листьев ювенильных растений рассматриваемого вида вдоль высотного градиента.
Однолетние сеянцы наибольшие размеры листа формируют в условиях Цудахарской экспериментальной базы, наименьшие – в условиях Гунибской экспериментальной базы. Ряд авторов (Гамалей, Шийревдамба, 1988; Kozlowski et al., 1991) отмечают, что в засушливых условиях растения формируют более мелкие листья. В связи с этим часто обнаруживают положительную корреляцию площади листа с количеством осадков (Мигалина и др., 2009).
В Дагестане приморская низменность характеризуется аридными условиями, соответственно оптимум роста многих видов вдоль высоты над уровнем моря будет находиться в среднегорном поясе. В дальнейшем с возрастанием высоты над уровнем моря усиливается лимитирующее действие низких температур. Это приводит к замедлению роста побегов и листьев, что мы наблюдали в условиях Гунибского плато. Наглядно это можно увидеть на рисунке.
Относительная изменчивость размеров листа вдоль климатических градиентов основана на адаптации фотосинтеза и водного режима растения к условиям среды. Такие признаки листьев, как длина черешка, длина и ширина листовой пластинки, общая длина листа, характеризуются средним уровнем варьирования.
Индекс формы листа в целом характеризовался низким уровнем варьирования, что говорит о стабильности данного признака. Стоит также отметить, что относительная изменчивость сильно варьировала в разных экологических условиях. Уровень изменчивости в целом был выше на тех высотных уровнях, где формировались более крупные размеры листьев, т.е. в более благоприятных условиях, а наименьшая вариабельность наблюдалась в неблагоприятных условиях. Фактор условия произрастания оказал значительное влияние на изменчивость всех учтенных признаков, за исключением индекса формы листа.
Сеянцы древесных растений вынуждены проходить различные экобиоморфологические состояния – сначала травянистую, после кустарниковую или древесную жизненную форму. Причем одревеснение побегов у древесных видов должно произойти в первый вегетационный период. Если этого не произойдет, надземная часть однолетних сеянцев после перезимовки может погибнуть – частично либо до основания.
Известно, что вовремя наступившее прекращение роста и переход в длительный и глубокий покой обеспечивают устойчивость растения в зимний период, т.е. зимостойкие растения отличаются интенсивным и коротким периодом роста, свидетельствующим о своевременной и достаточной подготовленности к вегетации (Сергеев, 1953; Туманов, 1979).
У изучаемого вида в первый год развития наблюдалось подмерзание побегов (табл. 5), но нами не было отмечено растений, получивших 5-балльную оценку (т.е. полностью погибших). В условиях Махачкалы зимостойкость составила 2 балла (обмерзает до 50% длины побега), а на Гунибской и Цудахарской базах – 3 балла (обмерзает более 50% длины побега).
Таблица 5.
Сравнительная оценка сеянцев Celtis caucasica по признакам подмерзания
| Место произрастания | Общая длина сеянца, см | Подмерзание от общей длины сеянца, см | Доля подмерзания от общей длины сеянца, % | Зимо- стойкость, балл | |||
|---|---|---|---|---|---|---|---|
| x ± Sx | C.V., % | x ± Sx | C.V., % | x ± Sx | C.V., % | ||
| Махачкала | 31.3 ± 2.46 | 23.8 | 2.85 ± 0.35 | 39.5 | 9.1 ± 0.28 | 14.3 | 2 |
| Цудахарская база | 65.3 ± 3.45 | 17.8 | 50.0 ± 3.75 | 26.9 | 76.7 ± 2.25 | 9.2 | 3 |
| Гунибская база | 21.9 ± 1.32 | 14.8 | 20.9 ± 1.05 | 14.6 | 95.4 ± 0.52 | 1.3 | 3 |
Примечание. Обозначения см. табл. 1.
Рис. 1.
Изменчивость морфологических признаков листа ювенильных растений Celtis caucasica вдоль высотного градиента. ◼ – длина черешка, △ – длина листа, ◇ – ширина листа, ⚪ – общая длина листа. Общая длина листа соответствует длине листовой пластины вместе с черешком.
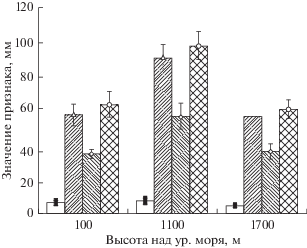
Корреляционный анализ показал, что имеется достоверная, на 0.05-м уровне значимости, сопряженность и взаимосвязь между длиной годичного прироста сеянцев и подмерзанием, т.е. чем более сильный годичный прирост, тем выше процент подмерзания от общей длины сеянца (табл. 5). Ранее в нашей работе уже была описана данная взаимосвязь (Магомедова и др., 2012), сейчас она нашла новое подтверждение.
C. caucasica в настоящее время произрастает на Цудахарской и Гунибской экспериментальных базах. Пятилетние саженцы на Цудахарской базе в настоящее время достигают высоты 2–2.5 м. На Гунибском плато в связи с ежегодным подмерзанием высота сеянцев не превышает 0.5 м.
Заключение. В объединенной выборке все признаки листа характеризуются средним уровнем изменчивости. Среди изученных признаков плодов C. caucasica такие признаки, как длина плодоножки (18.2%), вес плода (20.4%), вес косточки (17.6%), относятся к группе со средним коэффициентом вариации. Очень низким уровнем изменчивости характеризуются длина плода (5.6%), ширина плода (7.4%), индекс плода (6.7%), длина косточки (6.1%), ширина косточки (7%) как генетически более стабильные.
Основной причиной низкой численности C. caucasica в Дагестане на сегодняшний день является низкая всхожесть семян в естественных условиях (что подтверждено нами экспериментально) и отсутствие семенного возобновления в популяции. Лабораторная всхожесть семян C. caucasica составила 81.7%, т.е. высокий биологический потенциал семян этого растения недостаточно полно реализуется в природных условиях Предгорного Дагестана, что может быть следствием недостаточной экологической пластичности растений на первых этапах жизненного цикла. Темпы роста сеянцев в более благоприятных климатических условиях горно-долинного высотного пояса (1100 м над ур. моря) реализуются лучше, чем в условиях Гунибского плато (1750 м над ур. моря) и Махачкалы (100 м над ур. моря), что подтверждено также биометрическими показателями развития побеговых систем. Фактор условия произрастания оказал значительное влияние на изменчивость всех учтенных морфологических признаков листа, за исключением индекса формы листа. Корреляционный анализ свидетельствует, что имеются достоверные, на 0.05-м уровне значимости, сопряженность и взаимосвязь между длиной годичного прироста сеянцев и подмерзанием: чем выше годичный прирост, тем больше процент подмерзания от общей длины сеянца.
Изучение коррелятивных связей между анализируемыми признаками позволило выявить положительную корреляцию по признакам плода и косточки, длине и ширине листа.
Для редкого и уязвимого вида C. caucasica полученные морфологические показатели проростков необходимы для оценки успешности семенного размножения, как с целью практического использования, так и для реинтродукции в природные места обитания. К настоящему времени достоверно установлено, что семенной способ размножения при интродукции в целом является более эффективным, так как обеспечивает лучшую адаптацию к новым условиям произрастания (Некрасов, 1980).
Работа выполнена на уникальной научной установке “Система экспериментальных баз Горного ботанического сада ДагНЦ РАН”.
Список литературы
Абдурахманов Г.М. Красная книга Республики Дагестан. Махачкала: ИПЭ РД, 2009. 552 с.
Акаев Б.А., Атаев З.В., Гаджиев Б.С. Физическая география Дагестана. М.: Школа, 1996. 382 с.
Александрова М.С., Булыгин Н.Е., Ворошилов В.Н. Методика фенологических наблюдений в Ботанических садах СССР. М.: Наука, 1975. 28 с.
Галушко А.И. Деревья и кустарники Северного Кавказа // Дикорастущие, культивируемые и перспективные для интродукции. Нальчик: Кабардино-Балкарский научно-исследовательский институт, Кабардино-Балкарский республиканский ботанический сад, 1967. 535 с.
Гамалей Ю.В., Шийревдамба Ц. Структурные типы пустынных растений // Пустыни Заалтайской Гоби: характеристика растений-доминантов. Л.: Наука, 1988. С. 45–66 (217 с.).
Гроcсгейм А.А. Флора Кавказа. Saururaceae – Caryophyllaceae. Баку: Изд-во АзФАН, 1945. Т. III. 321 с.
Гулисашвили В.З. Дендрофлора Кавказа: Дикорастущие и культурные деревья и кустарники. Angiospermae – покрытосеменные: Dicotyledoneae – двудольные. Тбилиси: Изд-во АН ГССР, 1961. Т. 2. 335 с.
http://irbis.bti.secna.ru/doc5/2015-23.pdf, (26 November 2018).
Древесные растения Главного ботанического сада АН СССР. М.: Наука, 1975. 547 с.
Иваненко Б.И. Фенология древесных и кустарниковых пород. М.: Сельхозиздат, 1962. 184 с.
Кавеленова Л.М., Малыхина Е.В., Розно С.А., Смирнов Ю.В. К методологии экофизиологических исследований листьев древесных растений // Поволжский экологический журн. 2008. № 3. С. 200–210.
Князева С.Г. Изменчивость морфологических признаков сибирских видов можжевельника // Лесоведение. 2007. № 1. С. 65–69
Магомедова Б.М., Анатов Д.М., Рамазанова З.Р. Особенности роста и развития ювенильных древесных растений урбанофлоры г. Махачкалы в условиях Горного Дагестана // Известия Дагестанского государственного педагогического университета. Естественные и точные науки. 2012. № 2 (19). С. 22–28.
Магомедова Б.М., Мингажова М.М., Шаманова Ф.Х. Изменчивость признаков побега Salsola daghestanica (Turcz.) Turcz. (Chenopodiaceae) в Дагестане // Юг России: экология, развитие. 2016. Т. 11. № 4. С. 194–200.
Мигалина С.В., Иванова Л.А., Махнев А.К. Размеры листа березы как индикатор ее продуктивности вдали от климатического оптимума // Физиология растений. 2009. Т. 56. № 6. С. 858–862.
Некрасов В.И. Актуальные вопросы развития теории акклиматизации растений. М.: Наука, 1980. 101 с.
Николаева М.Г., Разумова М.В., Гладкова В.Н. Справочник по проращиванию покоящихся семян. Л.: Наука, 1985. 348 с.
Романова Н.Г., Ковригина Л.Н. Влияние погодных условий на строение очередных метамеров вегетативных побегов Sorbus sibirica Hebl. // Матер. Всерос. конф. “Фундаментальные и прикладные проблемы ботаники в начале XXI века” (Петрозаводск, 22–27 сентября 2008 г). Петрозаводск: Изд-во КарНЦ РАН, 2008. Ч. 1. С. 220–223.
Сергеев Л.И. Выносливость растений. М.: Советская наука, 1953. 284 с.
Туманов И.И. Физиология закаливания и морозостойкости растений. М.: Наука, 1979. 350 с.
Хохряков А.П., Мазуренко М.Т. Бластоид – элементарный блок побеговых растений // Жизненные формы, онтогенез и структура. 1993. С. 118–122 (230 с.).
Шмидт В.М. Развитие представлений о корреляциях и корреляционной структуре биологических объектов // Исследование биологических систем математическими методами. Л.: Изд-во ЛГУ, 1985. С. 5–18 (171 с.).
Kozlowski T.T., Kramer P.J., Pallardy S.G. The physiological ecology of woody plants, San Diego – London: Academic Press, 1991. 657 p.
Дополнительные материалы отсутствуют.