

Лесоведение, 2019, № 6, стр. 573-579
Семядоли проростков и зародышей хвойных
М. Г. Романовский a, *, Г. П. Морозов a
a Институт лесоведения РАН
143030 с. Успенское, Одинцовского района Московской области, ул. Советская, 21, Россия
* E-mail: michrom@ilan.ras.ru
Поступила в редакцию 20.08.2014
После доработки 20.02.2019
Принята к публикации 05.06.2019
Аннотация
Возможность использования числа семядолей при ранней диагностике быстроты роста неоднократно дискутировалась в отечественной лесоводственной литературе в связи с наследуемостью признака, декстральной асимметрией всходов, четностью числа семядолей и т.д. Исследования эмбрионов сосны кедровой сибирской заставляют по-новому взглянуть на генезис этого признака. Семядоли закладываются эпидермальными слоями эмбриона. Как правило, два слоя покровов, несущих семядоли, имеют билатеральную симметрию. Апекс проростка, контролирующий высоту, активируется позже, при прорастании. Рассмотрены результаты мониторинга семядолей у проростков и сеянцев сосны обыкновенной. Обсуждается роль наследственной изменчивости в определении числа семядолей.
При разработке политенной модели дифференциации тканей у растений была предложена гипотетическая модель развития апекса и появления семядолей (СД) у четырехсемядольного зародыша (Романовский, 2014; Романовский, Щекалев, 2014). Ожидалось, что при взгляде на апекс дифференцированного зародыша сверху можно будет увидеть радиально-симметричную картину. К каждой СД от апекса будет тянуться радиальная цепочка клеток, возможно, заметная через бинокулярный микроскоп, а между цепочками клетки расположатся без особой системы.
ОБЪЕКТЫ И МЕТОДИКА
Эмбриологический признак, описывающий фундаментальные свойства зародышей, будет присущ всем представителям семейства сосновых. Поэтому выбор конкретного вида определялся только соображениями удобства исследования. В качестве наиболее удобного объекта для массового анализа изменчивости числа семядолей (СД) у зародышей и проростков сосны были выбраны лущеные кедровые орешки. Зародыши сосны кедровой сибирской легко извлекаются из эндосперма, достаточно прочны, имеют длину до 1 см (100 мкм) и более. Под бинокулярной лупой МБС-10 с приближением 0.05 мм (50 мкм) можно подсчитать число семядолей и измерить параметры зародышей и эндоспермов. Использована партия семян Pinus sibirica Du Tour. из Новосибирской области.
При обработке обучающей выборки зародышей были зарисованы апексы каждого их них (всего 201 апекс). Измерены длины зародышей и размеры эндоспермов (максимальная ширина и длина).
Проанализированы также результаты мониторинга посевов сосны обыкновенной единичными семенами с подсчетом числа СД у проростков в питомнике Серебряноборского опытного лесничества Института лесоведения РАН. Посев семян проводили по сетке 5 × 5 см2 на грядках шириной 1 м подеревными площадками 1 × 1 м2. Перед посевом определяли массу 1000 полных отсортированных семян (удалены пустые) и проращивали партии семян, по 100 шт. на каждую семью. Высеяны 52 полусибсовых семьи из Московской области (урожай 1984 г.) и 90 семей из Киевской области (урожай 1986 г.). После расширения МКАД посевы и лесной питомник в 1991 г. были вынуждено ликвидированы.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
При взгляде на апекс зародыша сверху обнаружилась совершенно неожиданная картина. Во-первых, вопреки устоявшемуся стереотипу, апикальные зоны были не округлые, а в подавляющем большинстве эллиптические, с соотношением осей ~1.0 : 1.5 (от 1 : 1 до 1 : 2). Во-вторых, вдоль длинной оси эллипса были, как правило, обозначены две радиальные щели, разделявшие семядоли на 2 группы. На рис. 1 наблюдаемые картины деления семядолей щелями представлены схематически; эллиптические области условно заменены окружностями. Варианты деления апекса, приведенные в табл. 1, пронумерованы в соответствии с числом радиусов, делящих эллипс (кольцо) СД на дуги.
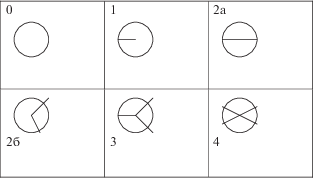
Таблица 1.
Варианты разделения радиальными щелями семядолей (СД) зародышей Pinus sibirica Du Tour
| Параметр | Виды вершины зародыша (см. рис.) | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 2а | 2б | 3 | 4 | |
| Число щелей, разделяющих эллипс (кольцо) СД, и разница чисел СД в дугах | Нет видимых щелей. СД сомкнуты, или разомкнутые, короткие, выстроенные в круг | Радиальная щель. Возможно не обнаружено диаметральное продолжение щели | Диаметральная щель. Числа СД в полуэллипсах равны или различаются на 1, реже 2–3 | Неравные сектора. Разница на 2–4 СД, очень редко – 5 СД | Числа СД различаются на 0–2, или равны только в двух секторах | Чаще всего секторы с попарно равным числом СД |
| Число зародышей | 22 | 24 | 104 | 18 | 19 | 14 |
| Частота, % | 11 | 12 | 52 | 9 | 9 | 7 |
| P(0.9)*, % | 4–28 | 4–21 | 43–56 | 2–35 | 2–34 | 1–38 |
Абсолютно преобладают (~61%) случаи деления СД зародыша щелью, состоящей из двух радиусов, на две примерно равные или неравные части (рис. 1, табл. 1, варианты 2а и 2б). СД группируются в двух половинах или секторах апикального эллипса. Варианты дробления СД на две дуги 2а и 2б принципиально ничем не отличаются.
Вариант 1 (рис. 1) объединяет случаи, когда семядоли образуют одну сплошную дугу, концы которой соединяются (12%). В вариант 1 могли отчасти войти случаи незафиксированного наблюдателем разделения семядолей на 2 группы, типа 2а и 2б.
Вариант 0 (рис. 1, табл. 1) объединил случаи отсутствия видимого разделения семядолей на группы (11%). Зародыши, включенные в него, неоднородны: это мелкие зародыши с короткими, отдельно стоящими по кругу семядолями (~3%), и “нормальные” зародыши с семядолями, компактно собранными вокруг апикальной точки (~8%). В первом случае зародышам как бы не хватило клеточного материала для завершения дифференциации. Длина большой и малой осей апикального эллипса близка (соотношение ~1 : 1), радиальные щели не обнаружены.
Редкие случаи разделения семядолей на 3 или 4 группы, суммарная частота которых не превышает 16%, представлены в последних графах табл. 1.
Интересен единичный случай сохраненной полиэмбрионии. Коррозионная камера эндосперма содержала 3 зародыша: один зародыш в халазальном конце камеры был очень короткий (~1 мм), с 10 зачатками СД, и завершал суспензор (подвеску); еще два зародыша с пятью длинными разомкнутыми клиновидными СД находились ближе к основанию суспензора.
В наиболее распространенном случае распределения СД по двум дугам их числа равны (Δ = 0; Р(∆) = 31%) или различаются на 1 (Δ = 1; Р(∆) = = 37%). Реже происходит деление СД-эллипса на 2 неравные дуги (Δ = 2; Р(∆) = 22%); различия на 3 и большее число СД в сумме встречаются с частотой ~10%. Общее число семядолей эмбриона чаще всего четное, P(∆ = 0, 2, 4) > P(∆ = 1, 3, 5) (табл. 2).
Таблица 2.
Различия в числах СД при делении радиальными щелями эллипса СД зародыша Pinus sibirica Du Tour. на 2 дуги и их частота встречаемости
| Параметр | Различия в числах СД (∆, шт.) | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | |
| Частота, Р(∆), шт.* | 42 | 50 | 30 | 9 | 1 | 2 |
| Частота, Р(∆), % | 31 | 37 | 22 | 7 | 1 | 1 |
| Интервал Р(0.9), %** | 21–44 | 30–49 | 12–37 | 1–35 | 0–35 | 0–35 |
Из наблюдений, выполненных авторами в питомнике Серебряноборского опытного лесничества Института лесоведения РАН, в связи с данной статьей наибольший интерес представляют оценки среднего числа семядолей в полусибсовых семьях сосны обыкновенной (подеревные потомства) и параллельные определения массы 1000 семян по семьям (М1000). Партии полусибсовых семян часто были немногочисленны, но их массу пересчитывали в М1000, выраженную в граммах (стандартный показатель, принятый в лесном семеноведении). Масса отдельного семени, тем более М1000, лишь косвенно связана с массой зародыша, от которой, как предполагалось ранее, и зависит число закладывающихся семядолей (Романовский, Морозов, 1992).
По результатам проращивания семян, собранных в Украине, для выборок по 30 семей из трех популяций сосны (Лютеж, Сорокошичье, Добрянка) были рассчитаны коэффициенты корреляции r среднесемейных значений числа СД и М1000. Они изменялись в пределах 0.24–0.35. По совокупности 90 полусибсовых семей корреляция М1000 и числа СД у всходов r = 0.28 ± 0.05.
В табл. 3 представлено распределение всходов сосны обыкновенной по числу СД в этих семейных посевах. До 6 лет дожили сеянцы, обладавшие на стадии проростков 4–8 семядолями. Редкие проростки с тремя СД (0.13%), отмеченные при лабораторном тестировании семян, не взошли. Проростки с девятью СД (0.04%) взошли, но полностью выпали из посева к трехлетнему возрасту. Дальнейший рост сеянцев шестилеток был в среднем обратно пропорционален (r > –0.1, зависимость недостоверна) числу СД, зарегистрированному у них при прорастании в грунте (табл. 3).
Таблица 3.
Число семядолей, рост и сохранность моноподиально ветвящихся сеянцев в штучных посевах сосны обыкновенной, Московская область*
| Параметр | Число семядолей у всходов | |||||
|---|---|---|---|---|---|---|
| 4 | 5 | 6 | 7 | 8 | 9 | |
| Число всходов 1985 г. | 79 | 562 | 1043 | 325 | 35 | 2 |
| Число сеянцев 1990 г.** | 29 | 174 | 354 | 114 | 15 | 0 |
| Выживаемость 1990, % | 37 ± 5 | 31 ± 2 | 34 ± 1 | 35 ± 3 | 43 ± 8 | – |
| Высота 1990 г., см ± m | 56.5 ± 5.7 | 55.0 ± 2.2 | 56.1 ± 1.5 | 53.3 ± 2.7 | 53.6 ± 7.1 | – |
По среднему числу СД полусибсовые семьи, выращенные из семян урожаев 1984 г. (из Московской обл.) и 1986 г (из Киевской обл.), разделились на три кластера с равновероятными распределениями в каждом. Для сосновых семян из обоих мест заготовки сосновых семян выделяются 3 интервала средних чисел СД – 4.8–5.3, 5.4–6.0 и 6.1–6.6 (Украина) и 5.1–5.5, 5.6–6.1, 6.2–6.5 (Россия), внутри которых полусибсовые семьи (общее число семей – 142) распределены равномерно. Распределения, равновероятные на некоторых интервалах значений признака, свойственны гаплоидным объектам (Романовский, Морозов, 1992; Романовский, 2014; Романовский, Щекалев, 2014). По-видимому, масса семени отражает не столько массу зародыша, сколько массу гаплоидного женского гаметофита, служившего ему пищей, а трофический режим эмбриона лишь в некоторой степени (r ≈ 0.3) определяет его размеры (Романовский, Хромова, 2012).
После полной смены очередной хвои побегов хвоей брахибластов, происходящей в возрасте 3 лет или старше, рост сеянцев сосны обыкновенной в комфортных условиях определяется, прежде всего, числом квантов роста, создаваемых апексом в период преформирования побега (Романовский, 2014). Число СД, венчающих эпителиальные лепестки зародыша, лишь косвенным образом связано с быстротой роста.
В незначительной степени (r ≤ 0.3) число СД определяется размерами зародыша и варьирующей массой питающего макрогаметофита (эндосперма).
Единственная известная нам работа, посвященная определению массы отдельных семян и числа СД у проростков, появившихся из этих же семян, выполнена П.П. Поповым (2013). Иных парных измерений индивидуальных параметров зародышей хвойных (масса семени – число СД или масса зародыша – число СД), по-видимому, не было. Масса семени и число СД у всхода, появившегося из данного семени ели, коррелируют: r = 0.32 ± 0.07 (Попов, 2013). Коэффициент корреляции средних масс семян и чисел СД, полученный нами в полусибсовых семьях сосны обыкновенной (r = 0.28 ± 0.05) полностью соответствует значению r, установленному для ели. Взаимосвязь средних размеров зародышей с числом СД в подеревных партиях семян косвенно подтверждает общность зависимости (Попов, 2013) для всех видов хвойных (Гаврись, 1938; Чтан Ши-цзюй, 1969; Попов, Жариков, 1974; Sorensen, Franklin, 1977; Голиков, 2007; Храмова 2009; Попов, 2013; и др.).
Число СД – исключительно устойчивый показатель. Даже экстремальные условия не способны сдвинуть его более чем на 1–2 СД (Шутяев, 1979; Котов, 1984; Zasada, 1988; Коба и др., 1996). Зародыш Pinaceae, СД которого участвуют в питании, заранее осваивает коррозионную камеру эндосперма (Романовский, Храмова, 2012). Размер камеры и, следовательно, максимальный размер зародыша предопределены еще перед оплодотворением (Zasada, 1988).
Колебания чисел СД в разных насаждениях одного лесорастительного района не превышают 1–2 СД (Гаврись, 1935, 1938). Мала и географическая изменчивость чисел СД, несмотря на необъятность ареалов многих хвойных видов Евразии. Так, колебания числа СД в пределах объединенного ареала елей сибирской и европейской ≤2.5 (Попов, 2013), 3.2 СД (Шутяев, 1979). Еще меньше (~2 СД) амплитуда колебаний числа СД у сосны обыкновенной (Попов, Жариков, 1974; Караваева, Попов, 1976; Голиков, Толстопятенко, 1978; Толстопятенко, 1978; Шутяев, 1979; Третьякова, Барсукова, 2012).
Ежегодно дерево восстанавливает варьирующее число СД, но при этом средние подеревные значения сохраняют индивидуальность. Рассчитанный П.П. Поповым (2013) коэффициент корреляции средних чисел СД у одних и тех же деревьев в урожаях разных лет составил 0.96 ± 0.08, что, по мнению автора, говорит о высоком постоянстве индивидуальности числа СД. Оптимальный объект для исследования вопроса о наследовании числа СД – соматические зародыши в культуре тканей (Третьякова, Барсукова, 2012; и др.).
В.М. Жариковым и В.Я. Поповым (1974) проведена уникальная работа по самоопылению разносемядольных деревьев, показавшая, что вклад наследственных особенностей материнских деревьев в изменчивость числа СД невелик. Такие же результаты были получены нами в опытах по самоопылению сосны обыкновенной, где были рассмотрены 3 дерева (данные не опубликованы).
СД имеют значение для поддержания начального роста всходов. У ели корреляция высоты однолеток и числа СД равнялась в среднем 0.51 (Голиков, Кирюхина, 1984). Для двухлеток ели коэффициент корреляции высоты и числа СД равнялся 0.35 ± 0.02 (Попов, 2013). В значительной мере от числа и длины СД зависели сохранность и рост сеянцев кедра сибирского (Храмова, 2009). Высота шестилеток сосны обыкновенной оказалась вообще не связана с числом СД, просматривается даже недостоверная отрицательная зависимость (табл. 3). С возрастом посевов влияние числа и длины СД на скорость роста сеянцев перестает читаться (Караваева, Попов, 1976; Романовский, Щекалев, 2014). Это заставляет усомниться в рекомендациях использования числа СД в качестве важного селекционного признака (Гаврись, 1935, 1938; Голиков, Толстопятенко, 1978; Толстопятенко, 1978, 1991; Голиков, Кирюхина, 1984; Коба и др., 1996; Голиков, 2007; Храмова, 2009; Попов, 2013; и др.).
Выявленные у эмбрионов особенности локализации СД (рис. 1, табл. 1) заставляют по-новому взглянуть на многочисленные попытки использования числа СД как удобнейшего показателя в начале селекции (Гаврись, 1935, 1938; и др.). Апексы, контролирующие вертикальный прирост, и семядоли формируются разными гистогенами в разные периоды онтогенеза. Семядоли порождаются самым внешним эпидермальным слоем клеток зародыша. Наподобие двух ладоней, разделенных отсутствием симпластных плазмодесменных контактов, он охватывает внутренние слои клеток, в которых и покоится мелкоклеточная группа клеток будущего деятельного апекса. Будущий апекс всхода только при его прорастании проникнет в щель между ладонями, несущими СД, и раздвинет ее.
Возникают сомнения в правомерности применения к СД представлений хиральной (левое – правое) симметрии (Толстопятенко, 1978, 1991; Хохрин, 1981). У зародышей господствует билатеральная симметрия. Прямой связи между числом СД и левизной – правизной (декстральной симметрией (Хохрин, 1981)) побега, сформированного позже проростком, нет. Однако опосредованная связь может наблюдаться: проростки, отличающиеся замедленным ростом, чаще всего антидромны, одновременно им свойственно и минимальное число СД (Голиков, 1982).
Вопрос о четном и нечетном числе семядолей, неоднократно обсуждавшийся в литературе, посвященной СД, также снимается с повестки дня. Все определяется вероятностью завершения двух зародышевых полусфер равным или неравным числом СД “ресничек” (Голиков, Толстопятенко, 1978; Толстопятенко, 1978, 1991; Голиков, 1982; Голиков, Кирюхина, 1984).
Дробление апекса у Pinaceae стирает непреодолимость грани между двудольными и хвойными растениями. СД хвойных растений, так же как СД двудольных, формируются покровными тканями зародышей и отличаются билатеральной симметрией.
Выводы. 1. Семядоли формируются задолго до прорастания семени и начала апикального роста зародыша как выросты на его апикальных слоях (обычно двух), произведенных эпителиальными тканями гипокотиля. В покоящемся семени СД завершены, а апекс будущего проростка находится в теле зародыша в виде мелкоклеточной массы, не видимой снаружи и не проявляющей активности.
2. Компоненты системы “зародыш–эндосперм” развиваются практически независимо. Связь числа СД со средней массой семян или М1000 в полусибсовых семьях хвойных деревьев незначительна (r ≈ 0.3). Близкая величина коэффициента корреляции найдена П.П. Поповым (2013) для числа СД и массы отдельного семени.
3. Среднее число СД у зародышей одного дерева устойчиво. Дерево из года в год повторяет варьирующее число семядолей. Опыты самоопыления подтверждают малую значимость наследственного варьирования в определении числа СД.
4. Высота сеянца определяется работой апикальной меристемы проростка после прорастания семени. Число СД и высота сеянца напрямую не связаны. В то же время сеянцы с минимальным числом СД, как правило, низкорослы.
5. Число СД влияет на ранние стадии роста проростков, всходов, но на поздних этапах не сказывается. Отбор материнских деревьев по числу СД для повышения продуктивности древостоев не целесообразен.
6. Прямой связи между числом СД и склонностью сеянца к антидромному филлотаксису (хиральная симметрия побегов высшего порядка противоположна материнским побегам) нет. Но малосемядольные зародыши, отличаясь замедленным ростом (предпосылка антидромии), чаще демонстрируют антидромию при закладке побегов второго порядка.
Список литературы
Гаврись В.П. Изучение корреляционной зависимости у хвойных методом геноанализа. Рукопись. Отчет по теме №25. Пушкино: Моск. НИИ лесн. хоз-ва, 1935. 77 с.
Гаврись В.П. Многоформенность хвойных пород и практическое использование ценных форм сосны и ели // Лесное хоз-во. 1938. № 1. С. 78–88.
Голиков А.М. Явление диссимметрии хвои у Pinus sylvestris (Pinaceae) // Ботанический журн. 1982. Т. 67. № 2. С. 220–223.
Голиков А.М. Рост и формовая структура потомства ели европейской в зависимости от гетерозиготности деревьев и условий произрастания // Лесоведение. 2007. № 4. С. 51–58.
Голиков А.М., Кирюхина Н.А. Величина семян и рост однолетних тепличных сеянцев ели // Выращивание и формирование высокопродуктивных насаждений в южной подзоне тайги. Л.: Лен. НИИ лесн. хоз-ва, 1984. С. 16–21.
Голиков А.М., Толстопятенко А.И. Некоторые особенности полиморфизма всходов сосны обыкновенной по числу семядолей // Восстановление леса на Северо-Западе РСФСР. Л.: Лен. НИИ лесн. хоз-ва, 1978. С. 144–146.
Жариков В.М., Попов В.Я. К вопросу о наследовании потомством сосны обыкновенной числа семядолей при направленном скрещивании различных биотипов // Состояние и перспективы развития лесной генетики, селекции, семеноводства и интродукции. Методы селекции древесных пород. Рига: Лат. НИИ лесохозяйственных проблем, 1974. С. 57–60.
Караваева А.А., Попов В.Я. О взаимосвязи некоторых анатомических и морфометрических признаков у разносемядольных всходов и сеянцев сосны обыкновенной // Вопросы лесовосстановления на Европейском Севере. Архангельск: Архангельск. институт леса и лесохимии, 1976. С. 68–74.
Коба В.П., Ругузов И.А., Молчанов Е.Ф. Изменчивость числа семядолей у всходов сосны крымской // Лесоведение. 1996. № 6. С. 67–70.
Котов М.М. Изучение наследственной изменчивости числа семядолей у сосны обыкновенной и модифицирующего влияния среды // Лесоводство, лесные культуры и почвоведение: рациональное использование и восстановление лесных ресурсов. Изд-во: ЛТА. 1984. С. 95–101.
Попов В.Я., Жариков В.М. Изменчивость потомства сосны обыкновенной по числу семядолей в разные репродуктивные годы // Вопросы лесокультурного дела на Европейском Севере. Архангельск: Архангельский институт леса и лесохимии 1974. С. 4–14.
Попов П.П. Популяционно-географическая изменчивость числа семядолей у всходов ели европейской и сибирской // Лесоведение. 2013. № 1. С. 9–15.
Романовский М.Г. Политения и гистогенез у лесных растений. Москва - Тула: Гриф и К, 2014. 123 с.
Романовский М.Г., Морозов Г.П. Эталонные характеристики двух форм сосны обыкновенной по наполненности шишек семенами // Лесоведение. 1992. № 3. С. 86–89.
Романовский М.Г., Хромова Л.В. Совместимость тканей в эмбриогенезе сосны // Лесоведение. 2012. № 2. С. 53–63.
Романовский М.Г., Щекалев Р.В. Система вида у лесных растений. М.: Товарищество научных изданий КМК, 2014. 212 с.
Толстопятенко А.И. Количество семядолей в связи с размером и цветом семян сосны обыкновенной // Восстановление леса на Северо-Западе РСФСР. Л.: Лен. НИИ лесн. хоз-ва, 1978. С. 149–150.
Толстопятенко А.И. Полиморфизм сосны и ели по числу семядолей в связи с изомерией шишек // Лесоведение. 1991. № 4. С. 71–79.
Третьякова И.Н., Барсукова А.В. Соматический эмбриогенез лиственниц и кедра сибирского в Сибири // Лесоведение. 2012. № 6. С. 63–70.
Хохрин А.В. Диссиметрическая изменчивость и стереобиология сосны обыкновенной // Экология. 1981. № 3. С. 58–65.
Храмова О.Ю. Репродуктивная способность и перспективы хозяйственного использование сосны кедровой сибирской (Pinus sibirica Du Tour) при интродукции в Поволжье (на примере Нижегородской области): Автореф … дисс. к.с-х.н. (спец. 06.03.01). М.: Моск. гос. университет леса, 2009. 22 с.
Чжан Ш.-Ц. Варьирование числа семядолей у всходов ели в связи с ее географическим происхождением и формовым разнообразием // Лесоведение. 1969. № 2. С. 79–81.
Шутяев А.М. Изменчивость числа семядолей у всходов сосны и ели // Лесоведение. 1979. № 3. С. 56–62.
Sorensen F.C., Franklin J.F. Influence of year of cone collection on seed weight and cotyledon number in Abies procera // Silvae Genetica. 1977. V. 26. № 1. P. 41–43.
Zasada J.C. Embryo growth in Alaskan white spruce seeds // Canadian J. Forest Research. 1988. V. 18. № 1. P. 64–67.
Дополнительные материалы отсутствуют.