

Лесоведение, 2021, № 1, стр. 11-27
Роль растительности в регулировании потоков тяжелых металлов в подтаежных лесных экосистемах центра Восточно-Европейской равнины
О. С. Железнова a, *, С. А. Тобратов a
a Рязанский государственный университет им. С.А. Есенина
390000 Рязань, ул. Свободы, 46, Россия
* E-mail: zheleznova_rzn@mail.ru
Поступила в редакцию 04.07.2018
После доработки 04.02.2019
Принята к публикации 06.06.2020
Аннотация
В методологии критических нагрузок древесная растительность рассматривается в качестве важнейшего источника буферности экосистем к потенциальному загрязнению, однако роль растений в регулировании потоков тяжелых металлов (ТМ) в лесных экосистемах изучена слабо. В настоящей работе на примере подтаежных экосистем Южной Мещеры (Рязанская область) анализируются факторы, влияющие на биогеохимическую устойчивость лесных экосистем к поступлению ТМ (Cu, Zn, Cd): закономерности транслокации и аккумуляции элементов в растительных органах; видовой состав растительности и геохимическая специализация видов; уровень биологической продуктивности древостоя; структура земель лесного фонда, в том числе доля антропогенно нарушенных земель. Результаты работы основываются на материалах почвенного и биогеохимического опробования 2013–2014 гг., таксационных данных, балансовых исследованиях. Показано, что накопление Cd в стволовой древесине может быть связано с его транспортом в ксилемном соке в свободной ионной форме, а накопление Cu – с акцепторной ролью древесины, лигнификация клеток которой невозможна без ряда Cu-содержащих ферментов. Построены картосхемы ежегодной иммобилизации ТМ в биомассе ствола и комля. Установлено, что пространственные закономерности аккумуляции ТМ в фитомассе существенно зависят от породного состава древостоя. Максимумы накопления Cu приурочены к участкам с преобладанием ариданитной сосны, а Zn и Cd – к участкам с преобладанием гумидокатных пород – березы и осины. Пространственные неоднородности биопродуктивности древостоев в существенной мере зависят от особенностей ландшафтной структуры территории – специфики погребенных морфоструктур и рельефа дневной поверхности. Результаты балансовых исследований свидетельствуют, что в фитомассе подтаежных экосистем в среднем может быть иммобилизовано до 46% атмосферных выпадений биофильных элементов – Cu и Zn – и менее 10% атмосферной поставки токсичного Cd. При этом Zn в Южной Мещере является слабодефицитным элементом, а Cu и особенно Cd – избыточными.
Антропогенное воздействие на лесные экосистемы приводит к изменению концентраций химических элементов в их компонентах, включению веществ антропогенного происхождения, в т. ч. тяжелых металлов (ТМ), в естественные циклы миграции (Лукина и др., 1994; Черненькова, 2004; Второва, Холопова, 2009). При этом биохимические механизмы миграции ТМ исследуются в настоящее время на достаточно ограниченном круге травянистых растений (Belleghem et al., 2007; Waters, Grusak, 2008; Sofo et al., 2012) или на молодых древесных растениях в вегетационных экспериментах (Cocozza et al., 2008; Di Baccio et al., 2009; Capuana, 2011; Vandecasteele et al., 2015), что недостаточно не только для региональных, но даже для экосистемных оценок химического загрязнения.
Тем не менее анализу влияния антропогенного загрязнения на состояние растительности лесов посвящено много работ (Никонов и др., 2004; Österås, 2004; Мартынюк, 2009; Huang et al., 2011; Сибиркина, 2014; и др.), однако роль самой растительности в регулировании потоков загрязняющих веществ в лесных экосистемах изучена в существенно меньшей степени. В условиях загрязнения древесная растительность выступает в качестве важнейшего источника естественной буферности лесных экосистем, обеспечивая долговременное изъятие элементов из миграции за счет их иммобилизации в тканях ствола и вероятное последующее их отчуждение при рубках (Manual …, 2004). В дальнейшем древесина, вовлекающаяся в хозяйственный оборот, превращается в отходы, однако они складируются на локальных участках, в основном в черте населенных пунктов. Тем самым отчуждение древесины при рубках оказывает положительный эффект на состояние экосистем, подвергающихся загрязнению через атмосферу. Оно, безусловно, не позволяет изъять ТМ из биосферы, но способствует существенной локализации их последующего участия в миграционных циклах.
Классические работы Н.И. Базилевич с соавт. (Базилевич, Родин, 1967; Базилевич, 1993), Н.П. Ремезова (1959) до сих пор считаются ключевым источником информации о биокруговоротах элементов в зональных экосистемах и рекомендуются к использованию при экологических оценках (Охрана …, 2004). Однако в настоящее время очевидны значительные трудности в адекватной экстраполяции этих данных и преимущества данных Государственного учета лесного фонда (ГУЛФ), обеспечивающие значительный территориальный охват и высокую корректность результатов (Усольцев, 2002).
Признанным на международном уровне механизмом оценки эколого-геохимической устойчивости экосистем является методология критических нагрузок. Критическая нагрузка соответствует максимальному уровню поступления поллютантов в экосистему, который не сопровождается необратимыми изменениями в функционировании биоты, структуре экосистемы и ее продуктивности в течение длительного периода времени (Башкин, Припутина, 2010). Согласно уравнению масс-баланса, используемому в методологии критических нагрузок при оценке геохимической устойчивости фоновых лесных экосистем, общее поступление металла должно уравновешиваться его накоплением в ежегодном приросте древесной растительности и выносом со стоком (Охрана …, 2004). Результаты наших предварительных исследований показывают, что долговременная иммобилизация ТМ в тканях ствола может составлять 22–76% от общей величины критической нагрузки (Кривцов и др., 2011).
Цель настоящей работы – рассмотреть факторы, определяющие регулирующую роль растительности лесных экосистем по отношению к потокам ТМ (на примере Cu, Zn, Cd), и оценить потенциал фитомассы по долговременной иммобилизации поллютантов. Cu и Zn – это биофильные элементы, активно участвующие в процессах метаболизма (фотосинтезе, дыхании, синтезе протеинов, защите от окислительного стресса, поддержании целостности мембран), но становящиеся токсичными для растений при избыточном поступлении в среду (Marschner’s …, 2012; Sinclair, Kramer, 2012; Singh et al., 2016). Cd является геохимическим аналогом Zn, однако метаболической потребности в данном элементе у растений нет (Cocozza et al., 2008). Со второй половины ХХ в. Cd по темпам и масштабам загрязнения стал одним из приоритетных поллютантов планеты (Волков, 2003).
ОБЪЕКТЫ И МЕТОДИКА
Территория исследования расположена на юго-западе Мещерской низины в центре Восточно-Европейской равнины (Солотчинский лесхоз Рязанской области) в зоне хвойно-широколиственных лесов (Природа …, 2008) и занимает площадь 389.6 км2. Характерная особенность Южной Мещеры – это малоконтрастный рельеф влажных, сырых и заболоченных песчаных равнин. Затрудненная гидродинамика и низкотрофные песчаные субстраты, являющиеся наследием четвертичных оледенений, определяют специфику почвенно-растительных условий района исследований. Дерново-подзолистые почвы плакоров в условиях ослабления дренажа сменяются болотно-подзолистыми и болотными почвами. К видам-эдификаторам растительных сообществ относятся сосна обыкновенная (Pinus sylvestris), мелколиственные породы – береза повислая (Betula pendula) и осина обыкновенная (Populus tremula), а в условиях повышенной трофности субстрата – также дуб черешчатый (Quercus robur), ель европейская (Picea abies) и ольха черная (Alnus glutinosa). Преобладающими древесными породами являются сосна и береза. Территория расположена на относительном удалении от крупных промышленных эмитентов (ближайший из них – южный промузел г. Рязани в 30 км к югу).
Источником первичной информации о продуктивности растительных сообществ послужили таксационные описания ГУЛФ для Солотчинского лесхоза (таксация 2002 г.). Всего были обработаны данные по более чем 300 лесным кварталам (свыше 3000 лесотаксационных выделов). Запасы отдельных фракций фитомассы рассчитывались на основе разработанных нами регрессионных зависимостей запасов от таксационных показателей древостоя. Запасы и продуктивность нижних ярусов оценивались по обширной базе данных “Продуктивность экосистем Северной Евразии” (2002–2006 гг.), созданной Н.И. Базилевич с соавт. Методика расчета запасов биомассы и продуктивности древостоя и нижних ярусов фитоценозов изложена в наших работах (Тобратов и др., 2016; Железнова, 2017).
На основе абсолютных значений запасов биомассы ствола рассчитывался производный параметр – коэффициент благоприятности состояния древостоя (Kб):
где Вфакт – запасы биомассы стволов с корой при фактической полноте древостоя (в абсолютно сухом состоянии, т га–1); ВЗН – зональный возрастной норматив запасов биомассы стволов с корой (в абсолютно сухом состоянии, т га–1) для подтаежных лесов центра Восточно-Европейской равнины в условиях современного климата, определяемый согласно (Усольцев, 2002). Kб, в отличие от абсолютных значений запасов, свободен от влияния онтогенетического фактора – возраста и поэтому может быть использован для оценки воздействия экологических факторов на рост и продуктивность древостоев. Значения Kб > 1 свидетельствуют о повышенных запасах биомассы стволов, что, как правило, говорит о благоприятности условий для функционирования сообществ; значения Kб < 1 характеризуют противоположную ситуацию. Kб позволяет оценить лесорастительный потенциал территории, реализованный в накопленных запасах фитомассы.Сезонный отбор образцов почв (горизонт А1) и различных структурных компонентов фитоценозов (фракции фитомассы надземных и подземных органов древостоя и растений нижних ярусов) осуществлялся в 2013–2014 гг. в соответствии с классическими методиками изучения биологического круговорота (Базилевич и др., 1978). Были отработаны 23 ключевых участка, различных по условиям произрастания и видовой структуре сообществ. Образцы фитомассы измельчались и высушивались до воздушно-сухого веса. Корни тщательно отмывались и высушивались.
Химические анализы образцов фитомассы и почв осуществлялись на базе лаборатории геохимии ландшафтов Рязанского государственного университета им. С.А. Есенина. Для определения концентраций ТМ в фитомассе проводилось кислотное разложение образцов в автоклавах с использованием микроволновой системы (объемное соотношение реактивов на 1 г фитомассы 24.3 : 6 : 2 : 1 для HNO3, Н2О2, Н2SO4, HF, соответственно) и последующее упаривание. Для почв определялись подвижные формы ТМ (экстракция ацетатно-аммонийным буферным раствором с рН 4.8) и их валовые несиликатные формы (экстракция царской водкой) (Методические …, 1992; МУК 4.1.985-00; ПНД Ф 16.1:2.2:2.3:3.36-2002). Определение концентраций ТМ осуществлялось атомно-абсорбционным методом на пламенном спектрометре “Спектр 5-4”; всего проанализировано свыше 400 образцов. Для Zn в торфяных почвах и для Cd в минеральных и торфяных почвах нами зафиксированы превышения экологических нормативов (верхних пределов накопления элементов в почве данного типа при отсутствии техногенного загрязнения) по подвижным формам (Железнова и др., 2018).
Для оценки потенциала растительности экосистем по иммобилизации ТМ осуществлялся синтез пространственных оценок продуктивности и запасов фитомассы сообществ с оценками содержания элементов в различных структурных компонентах фитоценозов. Расчет баланса ТМ произведен относительно приходной статьи баланса в соответствии с идеологией метода критических нагрузок (Manual …, 2004). Расчет выноса ТМ с поверхностными водами осуществлялся на основании гидрохимических (концентрация ТМ) и гидрометрических (расход воды) исследований в замыкающем створе бассейна р. Белой – основного водотока района исследований. Всего в течение годового цикла 2010–2011 гг. отобрано 15 проб поверхностных вод; пробоотбор был сопряжен с измерениями расхода воды в живом сечении водотока при помощи гидрометрической вертушки ГР-21 (Наставления …, 1972). Среднегодовой удельный водный вынос ТМ был определен по формуле:
где Mrunoff – искомый удельный вынос (г км–2 год), интегрально характеризующий интенсивность водной миграции элементов; Сср-взв – средневзвешенные концентрации ТМ в водах р. Белой (мкг л–1), определенные с учетом не только колебаний химического состава в течение года, но и динамики водности основных гидрологических фаз (половодья, летней и зимней межени и т.д.); Qгод – объем воды, проходящий через живое сечение р. Белой за годовой цикл (м3 год–1); F – площадь водосборного бассейна выше замыкающего створа (343 км2).Атмосферные выпадения ТМ оценивались на основании данных снегосъемки, осуществленной в марте 2010 г. Определение запасов зимних осадков, накопленных в период устойчивого существования снежного покрова (конец декабря–середина марта 2010 г.: всего 78 дней), и концентраций ТМ в образцах снеговой воды позволило оценить среднесуточную интенсивность атмосферного потока ТМ на земную поверхность. Каждый смешанный образец снежного покрова (1 образец на 35 км2) состоял из 6–10 индивидуальных проб, отбираемых при помощи поршневого пробоотборника на двух идентичных участках площадью 20 м2 каждый. Расположение мест взятия индивидуальных проб отвечало принципу типического пропорционального отбора (Дмитриев, 1972), отражая фактическое соотношение между площадями крон, межкроновых пространств и безлесных территорий (если они имелись); учитывалось также соотношение площадей, занимаемых кронами различных видов деревьев и кустарников. Полученные данные были экстраполированы на весь годовой цикл, в соответствии с имеющимися методическими подходами (Микроэлементы …, 1974; Учватов, 1994): считается, что отличия между концентрациями ТМ в осадках холодного и теплого сезонов и величинами плотности их потока из атмосферы, как правило, не превышают 30%.
Аккумуляция элементов в торфе рассчитана по данным радиоуглеродной датировки торфяника “Емельяново болото”, согласно которой торф на глубине 45.5 см имеет возраст 2085 + 51 лет (ИГАН-3877, калиброванные данные). Это свидетельствует о средних темпах накопления торфа в субатлантический период голоцена около 0.218 мм год–1. Указанную скорость торфонакопления следует признать низкой, но типичной для ландшафтов центра Русской равнины в ветровой тени Среднерусской и Смоленско-Московской возвышенностей. Аналогичные данные (в диапазоне 0.20–0.33 мм год–1) получены независимо друг от друга иными авторами для низинных и верховых болот севера Мещерской низменности (Абрамова, 1999; Новенко и др., 2016) и пойменных торфяников верховьев Дона в районе Куликова поля (Новенко, 2015). В менее континентальных регионах Русской равнины – Белоруссии, Прибалтике и даже Карелии – скорости вертикального роста торфяников в 1.5–10 раз выше (Козлов, 2011; Кузнецов, 2012; Инишева и др., 2013; Напреенко-Дорохова и др., 2016; Напреенко, Напреенко-Дорохова, 2018).
Статистическая обработка данных выполнена в программных пакетах Statistica и Excel, построение картосхем – в среде ГИС “Карта 2005” (производство ОКБ “Панорама”).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты проведенного исследования свидетельствуют, что регулирующая роль растительности по отношению к потокам ТМ и в целом биогеохимическая устойчивость лесных экосистем к поступлению поллютантов определяются следующими основными факторами:
1) закономерностями транслокации и аккумуляции ТМ в растительных органах;
2) видовым составом растительности и геохимической специализацией видов;
3) уровнем биологической продуктивности древостоя;
4) структурой земель лесного фонда, в том числе долей антропогенно нарушенных земель.
Первый фактор – закономерности транслокации и аккумуляции ТМ в растительных органах – определяет потенциальные пулы элементов, ассоциированные с различными фракциями фитомассы. Различия в продолжительности жизни фракций и их дальнейшей судьбе после вырубки древостоя определяют емкость и интенсивность круговоротов элементов. Так, пул элементов, ассоциированный с периодически обновляющимися тканями и органами (листьями и хвоей, тонкими корнями), определяет масштабы рецикличной миграции, а элементы, иммобилизованные в стволовой древесине, надолго изымаются из миграции и могут отчуждаться из экосистемы в результате рубок. Наиболее характерные закономерности транслокации и аккумуляции ТМ растительностью подтаежных лесных экосистем установлены нами ранее (Железнова и др., 2017) и могут быть проиллюстрированы материалами табл. 1 и 2, в которых приведены значения запасов и ежегодной фиксации ТМ в структурных компонентах фитоценозов Южной Мещеры.
Таблица 1.
Запасы органического вещества и тяжелых металлов в лесных экосистемах Южной Мещеры
| Ярус, фракция фитомассы | Фитомасса | Тяжелые металлы в фитомассе | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cu | Zn | Cd | |||||||
| т га–1 | % | г га–1 | % | г га–1 | % | г га–1 | % | ||
| Древесный ярус |
Надземная часть | 84.24 | 77.4 | 199.27 | 67.8 | 1863.37 | 68.1 | 18.39 | 61.2 |
| в т.ч. в древесине ствола | 63.91 | 58.7 | 116.80 | 39.7 | 617.69 | 22.6 | 11.00 | 36.6 | |
| в т.ч. в коре ствола | 6.72 | 6.2 | 20.54 | 7.0 | 502.76 | 18.4 | 2.48 | 8.3 | |
| в т.ч. в тонких ветвях* | 3.87 | 3.6 | 24.24 | 8.2 | 225.04 | 8.2 | 1.59 | 5.3 | |
| в т.ч. в хвое и листьях | 3.70 | 3.4 | 26.43 | 9.0 | 337.68 | 12.3 | 1.26 | 4.2 | |
| Подземная часть в т.ч. в тонких корнях** |
19.05 | 17.5 | 59.12 | 20.1 | 601.74 | 22.0 | 8.78 | 29.2 | |
| 4.24 | 3.9 | 21.00 | 7.1 | 234.07 | 8.6 | 2.69 | 8.4 | ||
| Подлесок, травяно-кустарничковый и мохово-лишайниковый ярусы | 5.52 | 5.1 | 35.69 | 12.1 | 269.18 | 9.9 | 2.89 | 9.6 | |
| Итого по фитоценозу | 108.81 | 100 | 294.08 | 100 | 2734.29 | 100 | 30.06 | 100 | |
Примечание. В табл. 1 и 2: * диаметр тонких ветвей ≤5 мм; ** диаметр тонких корней ≤5 мм. Данные представительны для земель лесного фонда; земли населенных пунктов и сельхозназначения не учитывались.
Таблица 2.
Биологическая продуктивность лесных экосистем Южной Мещеры и вовлечение тяжелых металлов в продукционные процессы
| Ярус, фракция фитомассы | Прирост | Тяжелые металлы в продукции | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cu | Zn | Cd | |||||||
| т га–1 год | % | г га–1 год | % | г га–1 год | % | г га–1 год | % | ||
| Древесный ярус |
Надземная часть в т.ч. в тканях ствола в т.ч. в хвое и листьях |
3.74 | 44.9 | 18.38 | 40.4 | 208.08 | 46.5 | 1.15 | 29.6 |
| 1.45 | 17.5 | 2.76 | 6.1 | 24.92 | 5.5 | 0.29 | 7.4 | ||
| 1.61 | 19.3 | 11.77 | 25.9 | 165.93 | 37.0 | 0.59 | 15.2 | ||
| Подземная часть в т.ч. в тонких корнях |
2.37 | 28.5 | 11.33 | 24.7 | 121.66 | 27.2 | 1.41 | 36.3 | |
| 2.15 | 25.9 | 10.68 | 23.5 | 116.48 | 26.0 | 1.35 | 34.7 | ||
| Подлесок, травяно-кустарничковый и мохово-лишайниковый ярусы | 2.09 | 25.1 | 15.29 | 33.7 | 115.35 | 25.7 | 1.28 | 32.9 | |
| Средняя аккумуляция в ежегодно нарастающем слое торфа (0.218 мм) | 0.12 | 1.5 | 0.56 | 1.2 | 2.81 | 0.6 | 0.05 | 1.2 | |
| Итого по фитоценозу | 8.32 | 100 | 45.56 | 100 | 447.90 | 100 | 3.89 | 100 | |
Примечание. См. подписи к табл. 1.
Как следует из табл. 1, на долю стволовой древесины приходится около 59% биомассы фитоценоза, однако с данной фракцией фитомассы ассоциировано менее половины запасов ТМ (23–40% от общего количества запасов ТМ в растительности экосистем) в связи с низкой зольностью древесины. При этом минимальное накопление в стволовой древесине характерно для Zn (22.6%), что является вполне закономерным. Zn, существенная доля которого в ксилемном соке мигрирует в комплексе с хелаторами (Sinclair, Kramer, 2012; Bouain et al., 2014), поступает в ветви и листья деревьев (транспорт в комплексе с хелаторами ограничивает связывание металла с отрицательными зарядами клеточных стенок и увеличивает его транслокацию в побеги). Его геохимический аналог Cd, мигрирующий в ксилемном соке в основном в свободной ионной форме Cd2+ (Conn, Gilliham, 2010; Hazama et al., 2015), накапливается в ксилеме (36.6%), притягиваясь отрицательными зарядами полигалактуроновых кислот клеточных стенок.
Еще большее накопление в древесине (39.7%) характерно для Cu. Данный факт нельзя объяснить высоким сродством ионов Cu2+ с материалом клеточных стенок, поскольку ксилемный транспорт Cu, в отличие от Cd, осуществляется преимущественно в комплексе с хелаторами (Burkhead et al., 2009). По нашему мнению, накопление Cu в древесине связано с тем, что данная фракция фитомассы, наряду с фотосинтезирующими органами, является одним из ключевых акцепторов комплексов “Cu–лиганд”. Известно, что Cu оказывает сильное влияние на формирование и химический состав клеточных стенок, при этом влияние Cu на лигнификацию особенно выражено в клетках склеренхимы стеблевых тканей (Marschner’s …, 2012). Недостаточная лигнификация сосудов ксилемы в условиях тяжелого дефицита Cu связана с участием в биосинтезе лигнина по меньшей мере двух Cu-содержащих ферментов: полифенолоксидазы и диаминоксидазы. Дефицит Cu в высших растениях и связанные с ним нарушения биосинтеза лигнина вызывают изгибание и скручивание стеблей и ветвей, а также летнюю суховершинность у деревьев. Последний признак – характерное проявление нарушения водного транспорта вследствие недостаточной лигнификации сосудов ксилемы или структурной ослабленности клеточных стенок. Таким образом, молодая древесина, выступая одним из важнейших акцепторов Cu, транспортируемой в комплексе с хелаторами, способна аккумулировать и структурно связывать существенные количества данного металла.
Другой характерной особенностью распределения ТМ во фракциях фитомассы, как следует из табл. 1, является интенсивное накопление биофильных Cu и Zn в тонких ветвях, хвое и листьях. Накопление Cd в данных фракциях фитомассы снижено из-за существования биологического барьера (повышения избирательности мембранных транспортных систем в направлении от корней к фотосинтезирующим органам). Повышенная аккумуляция Cd в подземной фитомассе (почти 30% его суммарных запасов в растительности экосистем) связана с функционированием корневого биологического барьера (поясков Каспари), препятствующего интенсивной транслокации токсичных ТМ в надземные органы.
Как следует из табл. 2, долговременная иммобилизация ТМ в ежегодном приросте древесины и коры – важнейший источник буферности экосистем к потенциальному загрязнению – составляет 5.5–7.4% от величины суммарной ежегодной фиксации ТМ в приросте всех основных компонентов фитоценоза. В целом, на фоне рецикличной миграции, такая иммобилизация невелика: как следует из табл. 2, циклические потоки ТМ, связанные с обновлением листвы, хвои и тонких корней, в 6.8 (Cd)–11.3 (Zn) раза масштабнее. Минимальный масштаб рецикличной миграции характерен для токсичного Cd, что связано как с его ограниченной транслокацией в тонкие ветви и листья, так и с повышенной аккумуляцией в древесине стволов.
С нижними ярусами фитоценозов, как показано в табл. 1, ассоциировано около 10% запасов ТМ. При этом вклад растений нижних ярусов в продукционные процессы почти в 5 раз выше, чем в запасы вещества, в соответствии с этим возрастает и их роль в ежегодной фиксации ТМ в приросте биомассы (табл. 2).
Определенную прибавку к долговременной иммобилизации ТМ в тканях ствола может дать аккумуляция элементов в ежегодно нарастающем слое торфа, которая также представляет собой долговременное изъятие ТМ из миграции и, следовательно, фактор естественной буферности экосистем. Согласно нашим расчетам (табл. 2), аккумуляция ТМ в ежегодно нарастающем слое торфа в 5–10 раз меньше их иммобилизации в тканях ствола. Это связано, в первую очередь, с незначительной величиной ежегодных торфяных аккумуляций, которая в среднем в 12 раз меньше ежегодного прироста ствола.
Второй фактор, определяющий регулирующую роль растительности по отношению к потокам ТМ, – видовой состав и геохимическая специализация растений. Влияние биологических особенностей видов на их химический состав проявляется в существовании закрепленных наследственностью альтернативных стратегий устойчивости (толерантности) растений к ТМ: индикации, избегания (исключения) и истинной толерантности (гипераккумуляции) (Ernst, 2006; Серегин, 2009; Maestri et al., 2010; Sofo et al., 2012). Ярким примером реализации наследственного фактора в химической композиции растений является гумидокатность и ариданитность. Известно (Перельман, Касимов, 1999), что гумидокатные виды (в том числе произрастающие на территории исследования береза и осина), сформировавшиеся в гумидных ландшафтах, где преобладают кислые почвы, энергично накапливают подвижные в кислой среде катионогенные микроэлементы (в том числе Zn2+ и Cd2+). Ариданитные растения (дуб и сосна), возникшие в аридных ландшафтах, энергичнее накапливают подвижные в нейтральной и щелочной среде анионогенные микроэлементы. К числу последних может быть отнесен также Cu, поскольку известно (Ладонин, Марголина, 1997), что данный элемент тесно ассоциирован с органическим веществом почвы и мигрирует в основном в составе отрицательно заряженных органоминеральных комплексов.
Влияние видового состава растений на регулирование потоков ТМ в экосистемах Южной Мещеры рассмотрим совместно с еще одним фактором – биологической продуктивностью древостоя. При прочих равных условиях чем выше значение биопродуктивности, тем большее количество элемента будет вовлечено в рециркуляцию и иммобилизацию в многолетних тканях. В табл. 3 приведены значения коэффициентов корреляции между параметрами биогенной миграции ТМ и двумя факторами – породным составом древостоя и уровнем его биопродуктивности. Как следует из табл. 3, на накоплении рассматриваемых ТМ в фитомассе отражается ариданитность сосны и гумидокатность березы и осины. Чем больше доля сосны в древостое, тем больше масштаб иммобилизации Cu в тканях ствола и комля. Биогенная миграция Zn и Cd, напротив, отрицательно коррелирует с долей сосны и положительно – с долей березы и осины. Иммобилизация и рециркуляция всех ТМ положительно связана с уровнем биопродуктивности как сосны, так и березы. Исключением является лишь отсутствие значимой связи в системе параметров “иммобилизация Cu–продуктивность березы”, что обусловлено гумидокатностью данной древесной породы.
Таблица 3.
Корреляция параметров биогенной миграции тяжелых металлов – ежегодной иммобилизации в тканях ствола и комля и рецикличной миграции с породным составом древостоя и уровнем его биологической продуктивности
| Элемент | Параметр | |||
|---|---|---|---|---|
| корреляция между Мupt (Мcycl) и | ||||
| долей сосны, % | долей березы и осины, % | Кб сосны | Кб березы | |
| Cu | 0.49 (–0.01) | –0.47 (–0.01) | 0.51 (0.29) | –0.07 (0.15) |
| Zn | –0.63 (–0.32) | 0.69 (0.36) | 0.19 (0.24) | 0.46 (0.31) |
| Cd | –0.10 (–0.20) | 0.15 (0.22) | 0.45 (0.25) | 0.29 (0.24) |
Примечание. Мupt – ежегодная иммобилизация тяжелого металла в тканях ствола и комля; Мcycl – рецикличная миграция; Kб – коэффициент благоприятности состояния древостоя. При расчете Мcycl учтены периодически обновляющиеся ткани и органы растений древесного яруса (хвоя и листья, тонкие ветви (диаметр ≤5 мм), тонкие корни (диаметр ≤5 мм)), а также растения травяно-кустарничкового и мохово-лишайникового ярусов. Коэффициенты корреляции, выделенные полужирным шрифтом, значимы на высоком уровне (p < 0.05).
На рисунке 1 представлены картосхемы, иллюстрирующие пространственные закономерности масштабов ежегодной иммобилизации ТМ в биомассе ствола и комля. Ландшафтные местности, границы которых приведены на рис. 1, отражают объективное существование в пределах района исследований различных условий для функционирования сообществ. Различия между местностями обусловлены неодинаковой абсолютной высотой, набором морфоскульптурных форм, историей развития, спецификой погребенных морфоструктур и, следовательно, современной гидродинамикой (степенью дренированности). Эти факторы определяют пространственные неоднородности продуктивности древостоев района исследований (Железнова, Тобратов, 2017). Подчеркнем, что ландшафтную структуру территории, влияющую на породный состав древостоя и его биопродуктивность, можно выделить в качестве еще одного фактора, определяющего регулирующую роль растительности экосистем по отношению к потокам поллютантов.
Рис. 1.
Ежегодная иммобилизация тяжелых металлов (А – Cu; Б – Zn; В – Cd) в биомассе ствола и комля древостоев подтаежных экосистем Южной Мещеры. Ландшафтные местности: I – Солотчинская останцовая местность; II – Полковская депрессия; III – Пяткинская возвышенность; IV – Лопуховская наклонная равнина; V – Полянская наклонная равнина; VI – Ласковская поозерная равнина; VII – Борисковская равнина; VIII – Дорофеевская равнина; IX – Темновская равнина (бывшая Юго-Западная ложбина стока); X – Переделецкая равнина (бывшая Северная ложбина стока); XI – Красное болото.

Рис. 1.
Продолжение.
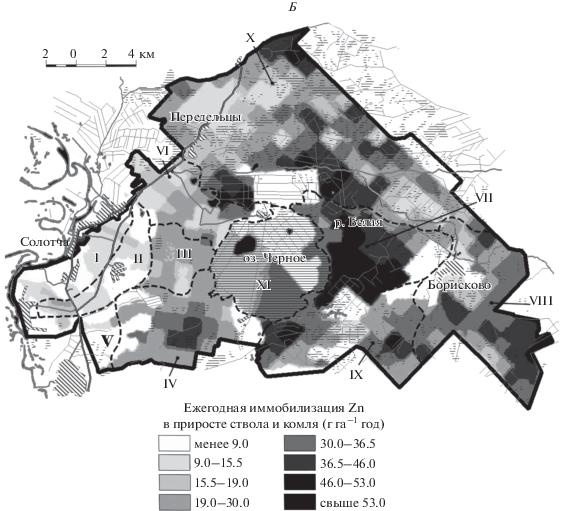
Рис. 1.
Окончание.
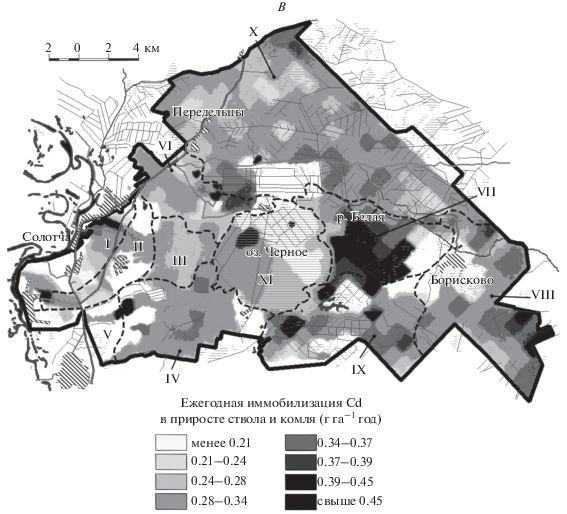
Как следует из рис. 1А, наиболее пространственно выраженные максимумы накопления Cu в фитомассе характерны для Солотчинской останцовой местности I и примыкающих территорий на западе района исследований. Особенностью местности I являются максимальные в пределах района исследований высотные отметки и вследствие этого – хорошая дренированность, господство старовозрастных, почти монодоминантных сосновых лесов, а также существенно превышающие зональный возрастной норматив запасы биомассы древостоя (табл. 4). Эти факторы обусловливают интенсивное накопление Cu в фитомассе экосистем Солотчинской местности I. Локальные максимумы биокруговоротов Cu вне местности I также соответствуют территориям с преобладанием высокопродуктивных сосновых древостоев. При этом аккумуляция Zn и Cd на этих участках и в самой Солотчинской местности близка к минимальной, что связано с ничтожным участием в породном составе древостоя гумидокатных березы и осины (рис. 1Б, В; табл. 4).
Таблица 4.
Породный состав и уровень биологической продуктивности древостоев Южной Мещеры
| Территория | Диапазон абсолютных высот, м | Породный состав древостоя, % | Кб сосны | Кб березы |
|---|---|---|---|---|
| Весь лесхоз | 103.5–130.2 | Сосна – 50.6; береза – 37.6; осина – 6.6; ольха – 3.1; дуб – 1.1; ель – 0.7; другие породы – 0.3 | 1.05 (0.37–1.79) |
0.80 (0.24–1.19) |
| Солотчинская останцовая местность I | 120.0–130.2 | Сосна – 92.4; береза – 3.1; дуб – 1.6; осина – 0.2; другие породы – 2.7 | 1.38 (1.01–1.74) |
0.96 |
| Борисковская сырая равнина VII | 108.9–116.8 | Береза – 55.5; сосна – 18.5; осина – 12.3; ольха – 9.4; дуб – 2.8; ель – 1.2; другие породы – 0.3 | 1.26 (0.96–1.79) |
0.89 (0.66–1.19) |
| Переделецкая заболоченная равнина X | 105.6–116.5 | Сосна – 54.3; береза – 40.2; осина – 2.6; ольха – 2.0; ель – 0.6; дуб – 0.2; другие породы – 0.1 | 0.92 (0.37–1.38) |
0.73 (0.43–0.96) |
Борисковская сырая равнина VII расположена на сниженной высотной ступени (табл. 4), что обеспечивает ее повышенное грунтовое увлажнение, однако она приурочена к характерному элементу погребенного рельефа – вершине доюрского известнякового эрозионного останца. Это частично компенсирует эффект переувлажнения и приводит к тому, что в пределах местности VII распространены сообщества с преобладанием гумидокатных влаголюбивых видов (влияние современного рельефа), но с максимальной для района исследований продуктивностью (фактор погребенного рельефа). В этой связи в данной местности масштабы ежегодной иммобилизации Cu в тканях ствола снижаются почти в 2 раза и соответствуют средним для территории значениям, в то время как биокруговороты Zn и Cd, напротив, максимальны (рис. 1). Максимумы биокруговоротов Zn и Cd обусловлены, таким образом, сочетанием двух факторов – активной аккумуляцией данных ТМ березой и осиной и высоким уровнем продуктивности древостоя.
Заболоченные территории (Переделецкая заболоченная равнина Х (табл. 4), обособленная в контурах крупной доплейстоценовой эрозионной ложбины, врезанной в кровлю регионального водоупора) с характерными для них сниженными запасами биомассы древостоя, в том числе и за счет его низкой полноты, маркируются сниженными масштабами иммобилизации ТМ в тканях стволов (рис. 1). При этом фактор породного состава так же, как и в случае с Борисковской местностью VII, играет приоритетную роль в пространственных закономерностях биокруговоротов ТМ. Локальные максимумы накопления Cu в тканях стволов в пределах Переделецкой местности Х приурочены к соснякам на сухих вереях, а Zn и Cd – к гидроморфным эдатопам с преобладанием березы.
Таким образом, биогеохимические особенности сосняков Солотчинской местности I определяют повышенную долговременную иммобилизацию Cu (в 1.8–2.0 раза выше, чем в местностях VII и Х), что является ее наиболее характерной особенностью. Гумидокатные породы Борисковской равнины, напротив, определяют приуроченность к ней максимумов биопоглощения Zn и Cd (соответственно в 1.8–3.6 и в 1.2–1.4 раза выше, чем в местностях Х и I). Несмотря на полуторакратное отставание в приросте, экосистемы Переделецкой гидроморфной равнины, тем не менее, в 2 раза опережают сосняки Солотчинской местности по ежегодной иммобилизации Zn в приросте древесины и коры ствола. Это наглядно свидетельствует, что при формировании иммобилизационного пула элементов в лесных экосистемах фактор породного состава более важен, чем фактор биопродуктивности.
Наконец, четвертый фактор, определяющий регулирующую роль растительности по отношению к потокам ТМ, – это особенности структуры земель лесного фонда. При этом особое значение имеет рост площади территорий, не покрытых лесом, где долговременная иммобилизация ТМ в приросте отсутствует, что приводит к снижению потенциала устойчивости экосистем к химическому загрязнению. Наличие территорий с нулевой или близкой к ней биогеохимической устойчивостью может быть обусловлено природными причинами: в условиях Мещеры – в основном развитием гидроморфных экосистем в термокарстовых понижениях рельефа (травяные болота – 1082.6 га, термокарстовые озера и речные русла – 240.8 га). Однако даже несмотря на высокую заболоченность Мещерской низменности, свыше 82% площадей безлесных территорий обусловлено антропогенным фактором: вырубки, квартальные просеки и дороги занимают в районе исследований 1456.1 га, антропогенные пустоши – 386.6 га, торфоразработки и заброшенные агроценозы – 4254.0 га. Заболачивание и уничтожение лесов способствуют снижению иммобилизационного потенциала лесных экосистем, определяемого масштабами накопления ТМ в тканях ствола, на 19.0% от теоретически возможного максимума, в том числе антропогенный фактор – на 15.7%. Следовательно, антропогенное вмешательство в лесные экосистемы центра Русской равнины приводит не только к снижению радиационного баланса территории на 11–12% (Дьяконов, 1991), но и к сопоставимому по масштабам снижению емкости биокруговоротов элементов.
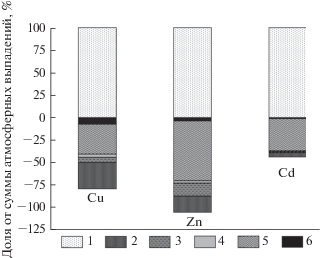
На рисунке 2 представлены результаты расчета балансов ТМ в экосистемах Южной Мещеры, которые позволяют дать интегральную оценку роли растительности в регулировании потоков ТМ. Как следует из рис. 2, биотический блок подтаежных экосистем способен иммобилизовать 39–46% атмосферных выпадений биофильных элементов – Cu и Zn – и менее 10% атмосферной поставки токсичного Cd (за счет аккумуляции в приросте древесины и коры ствола, тканей комля, а также накопления в ежегодно нарастающем слое торфа). При этом Zn в Южной Мещере является слабодефицитным элементом (невязка баланса равна –5.1%), а Cu и особенно Cd – избыточными (невязка балансов +20.4 и +55.7%, соответственно). Избыточность Cu обусловлена его сниженным водным выносом (слабая подвижность в природных растворах), а Cd – высоким уровнем антропогенного поступления по сравнению с санационными возможностями экосистем. Zn, будучи типоморфным элементом подтаежных ландшафтов, обладает одновременно и повышенным биопоглощением, и высокой гидрохимической подвижностью, которая и является главной причиной его дефицитности.
Рис. 2.
Баланс тяжелых металлов в лесных экосистемах Южной Мещеры. 1 – атмосферные выпадения; 2 – долговременная иммобилизация в древесине ствола; 3 – долговременная иммобилизация в коре ствола; 4 – долговременная иммобилизация в комле; 5 – вынос с поверхностными водами; 6 – накопление в ежегодно нарастающем слое торфа.
Топологическая (внутриландшафтная) дифференциация природы приводит к заметным колебаниям миграционных потоков ТМ от места к месту. При этом меняются как емкости их биокруговоротов, так и плотность потока металлов из атмосферы. В результате в разных местностях могут складываться различные соотношения входных и выходных потоков миграции ТМ, особенно с учетом того, что биогенная и абиогенная миграции управляются разными факторами с низкой вероятностью пространственных соответствий. Так, основная закономерность атмотехногенеза – повышенный уровень выпадений в “пойменном канале” воздушной миграции (Кривцов и др., 2011) и на “лесных”, пограничных территориях, непосредственно прилегающих к нему. Такое пограничное экотонное положение характерно для Солотчинской местности I, лесные массивы которой выступают в роли атмосферного фильтра для техногенных ТМ. К востоку от местности I атмосферные выпадения ТМ снижаются в 1.5–2 раза. Пространственные закономерности биогенной миграции значительно сложнее, но, как видно из табл. 5, масштаб внутриландшафтных различий, тем не менее, вполне сопоставим с таковым для абиогенных атмосферных выпадений. “Ядра типичности” наиболее показательных местностей I, VII и X различаются по интенсивности долговременной иммобилизации ТМ в 1.75–2.72 раза, и только типоморфный Zn является исключением – его биогенная миграция более контрастна, более зависима от местных условий, и различия между выбранными для анализа территориями достигают 5.8 раз.
Таблица 5.
Соотношение биогенных и абиогенных потоков тяжелых металлов в “ядрах типичности” некоторых местностей как характеристика масштаба пространственных ландшафтно-геохимических различий
| Элемент | Местность | Атмосферные выпадения, кг км–2 год |
Долговременная иммобилизация в фитомассе, кг км–2 год | Доля долговременной иммобилизации от атмосферных выпадений, % |
|---|---|---|---|---|
| Cu | I | 1.408 | 0.419 | 29.74 |
| VII | 0.982 | 0.311 | 31.69 | |
| X | 0.872 | 0.269 | 30.87 | |
| Zn | I | 12.169 | 0.930 | 7.64 |
| VII | 6.072 | 5.452 | 89.79 | |
| X | 6.684 | 2.531 | 37.87 | |
| Cd | I | 0.552 | 0.0271 | 4.91 |
| VII | 0.206 | 0.0455 | 22.10 | |
| X | 0.404 | 0.0295 | 7.30 |
Примечание. “Ядро типичности” – территория, прилегающая, как правило, к геометрическому центру местности, охватывающая наиболее типичный для нее набор фаций и урочищ, с наиболее характерным проявлением ландшафтно-геохимических свойств всей местности. При расчете долговременной иммобилизации в фитомассе учтена ежегодная аккумуляция элементов в тканях ствола и комля. Названия ландшафтных местностей I, VII, X см. табл. 4.
Биопоглощение ТМ определяется в основном геохимической специализацией и продуктивностью лесных экосистем. При этом монодоминантные ариданитные сосняки местности I специализируются на накоплении Cu, а на сниженных равнинах VII и X преобладают гумидокатные сообщества с преимущественным сродством к Zn и Cd. Однако местность Х обладает затрудненной гидродинамикой, и биогенная миграция элементов на ее территории снижена в связи с подавленностью продукционных процессов. К центру Борисковской равнины VII, наоборот, приурочен максимум биопродуктивности и параллельно с ним – один из минимумов атмосферной поставки ТМ. В результате экосистемы местности VII наиболее устойчивы к антропогенному поступлению Zn и Cd: доля долговременной иммобилизации от атмосферной поставки данных элементов превышает средние для Южной Мещеры значения в 2–2.5 раза (табл. 5 и рис. 2). В Солотчинской местности I, наоборот, иммобилизационные возможности древесного яруса по отношению к Zn и Cd в 1.5–6.0 раз ниже средних величин. Этому способствуют как незначительное участие гумидокатных видов в древостое, так и высокий уровень атмосферной поставки ТМ в местности I. Тем самым снижению устойчивости солотчинских сосняков к атмосферному поступлению Zn и Cd одновременно способствуют пространственные вариации и природных, и техногенных факторов. В то же время для Cu характерны противоположные закономерности: повышенный уровень атмосферной поставки в местности I компенсируется повышенным накоплением Cu, а в иных, менее экотонных местностях, удаленных от поймы р. Оки, биогенная иммобилизация и атмосферная поставка снижаются синхронно. Следствием этого является высокая пространственная стабильность буферных свойств экосистем к атмосферным выпадениям Cu, что следует признать примечательным, но случайным территориальным соответствием полей биогенной и абиогенной миграции.
ЗАКЛЮЧЕНИЕ
Таким образом, растительность может играть существенную роль в регулировании потоков загрязняющих веществ в подтаежных лесных экосистемах, иммобилизуя и выводя из миграции (за счет депонирования в тканях ствола, комля и последующего отчуждения при рубках) до 46% атмосферных выпадений ТМ. Наименьшая регулирующая роль растительности проявляется в отношении типичных токсикантов: например, долговременной иммобилизации Cd в фитомассе подвергается менее 10% его атмосферной поставки. При этом масштабы долговременной иммобилизации ТМ существенно варьируют в пределах района исследований: различия между ландшафтными местностями могут достигать 1.8–5.8 раз.
Основными факторами, определяющими емкость и интенсивность биокруговоротов ТМ и в целом биогеохимическую устойчивость лесных экосистем к поступлению поллютантов, являются закономерности транслокации и аккумуляции элементов в растительных органах, видовая специфика накопления ТМ, уровень биологической продуктивности древостоя, а также структура земель лесного фонда. Влияние этих факторов на масштабы иммобилизации ТМ в тканях ствола – важнейший, согласно методологии критических нагрузок, источник естественной буферности экосистем к загрязнению – рассмотрено нами на примере подтаежных лесных экосистем Южной Мещеры. Изучение пространственных закономерностей аккумуляции ТМ в ежегодном приросте тканей ствола показало, что приоритетная роль в определении масштабов иммобилизации ТМ в фитомассе принадлежит породному составу древостоя. Сообщества с преобладанием ариданитной породы – сосны – наиболее эффективно иммобилизуют Cu (сосняки на хорошо дренируемых почвах иммобилизуют Cu в 2 раза интенсивнее, чем в среднем для района исследований). При преобладании гумидокатных березы и осины активно – в среднем в 1.2–3.6 раза выше, чем в сосновых сообществах – иммобилизуются Zn и Cd. Поскольку уровень биопродуктивности древостоя и его породный состав являются функцией ландшафтной структуры территории, данный фактор также можно считать одним из определяющих при оценке регулирующей роли растительности по отношению к потокам ТМ. Установленные нами закономерности геохимических и биопродукционных процессов актуальны для большинства подтаежных ландшафтов полесского типа.
Список литературы
Абрамова Т.А. Результаты комплексного палеоботанического изучения эволюции ландшафтов Центральной Мещеры в голоцене // Ландшафтная школа Московского университета: традиции, достижения, перспективы / Под ред. Дьяконова К.Н., Мамай И.И. М.: РУСАКИ, 1999. С. 96–108.
Базилевич Н.И. Биологическая продуктивность экосистем Северной Евразии. М.: Наука, 1993. 293 с.
Базилевич Н.И., Родин Л.Е. Картосхемы продуктивности и биологического круговорота главнейших типов растительности суши // Известия Всесоюзного географ. общества. 1967. Т. 99. № 3. С. 190–194.
Базилевич Н.И., Титлянова А.А., Смирнов В.В. Методы изучения биологического круговорота в различных природных зонах. М.: Мысль, 1978. 183 с.
Башкин В.Н., Припутина И.В. Управление экологическими рисками при эмиссии поллютантов. М.: Газпром ВНИИГАЗ, 2010. 186 с.
Волков С.Н. Геохимическая эволюция кадмия в естественном и техногенном циклах миграции // Техногенез и биохимическая эволюция таксонов биосферы: Труды биогеохимической лаборатории. М.: Наука, 2003. Т. 24. С. 113–141.
Второва В.Н., Холопова Л.Б. Концентрации химических элементов в растениях и почве и оценка состояния лесных экосистем // Лесоведение. 2009. № 1. С. 11–17.
Дмитриев Е.А. Математическая статистика в почвоведении. М.: Изд-во МГУ, 1972. 272 с.
Дьяконов К.Н. Геофизика ландшафта. Биоэнергетика, модели, проблемы. М.: Изд-во МГУ, 1991. 96 с.
Железнова О.С. Комплексная оценка биогеохимической устойчивости экосистем в условиях техногенеза (на примере подтаежных лесов Южной Мещеры): Дис. … канд. биол. наук (спец. 03.02.08). М.: Российский университет дружбы народов, 2017. 297 с.
Железнова О.С., Тобратов С.А. Опыт ландшафтного анализа пространственных закономерностей продуктивности зональных экосистем Южной Мещеры // Известия РАН. Серия географ. 2017. № 6. С. 47–62.
Железнова О.С., Тобратов С.А., Кадыров А.С. Цинк и кадмий в подтаежных лесных экосистемах Восточно-Европейской равнины // Матер. Всеросс. науч. конф. “Химическое и биологическое загрязнение почв” (г. Пущино, 18–22 июня 2018 г.). Пущино: Товарищество научных изданий КМК, 2018. С. 18–20.
Железнова О.С., Черных Н.А., Тобратов С.А. Цинк и кадмий в фитомассе древесных растений лесных экосистем: закономерности транслокации, аккумуляции и барьерных механизмов // Вестник РУДН. Серия “Экология и безопасность жизнедеятельности”. 2017. Т. 25. № 2. С. 253–270.
Инишева Л.И., Кобак К.И., Турчинович И.Е. Развитие процесса заболачивания и аккумуляция углерода в болотных экосистемах России // География и природные ресурсы. 2013. № 3. С. 60–68.
Козлов Е.А. Географические особенности изменений скоростей накопления торфа // Вестник Брестского университета. Серия 5. 2011. № 1. С. 18–29.
Кривцов В.А., Тобратов С.А., Водорезов А.В., Комаров М.М., Железнова О.С., Соловьева Е.А. Природный потенциал ландшафтов Рязанской области. Рязань: Рязанский гос. университет им. С.А. Есенина, 2011. 768 с.
Кузнецов О.Л. Динамика растительности верховых болот // Известия Самарского НЦ РАН. 2012. Т. 14. № 1. С. 1288–1291.
Ладонин Д.В., Марголина С.Е. Взаимодействие гуминовых кислот с тяжелыми металлами // Почвоведение. 1997. № 7. С. 806–811.
Лукина Н.В., Никонов В.В., Райтио Х. Химический состав хвои сосны на Кольском полуострове // Лесоведение. 1994. № 6. С. 10–21.
Мартынюк А.А. Сосновые экосистемы в условиях аэротехногенного загрязнения, их сохранение и реабилитация: Дисс. … д-ра с.-х. наук (спец. 06.03.03). М.: Всерос. НИИ лесоводства и механизации лесного хозяйства, 2009. 380 с.
Методические указания по определению тяжелых металлов в почвах сельхозугодий и продукции растениеводства: Изд. 2-е, перераб. и доп. М.: Изд-во ЦИНАО, 1992. 61 с.
Микроэлементы в природных водах и атмосфере / Под ред. Жигаловской Т.Н., Малахова С.Г. М.: Гидрометеоиздат, 1974. 183 с.
МУК 4.1.985-00 Определение содержания токсичных элементов в пищевых продуктах и продовольственном сырье. Методика автоклавной пробоподготовки: Метод. указания. М.: Федеральный центр госсанэпиднадзора Минздрава России, 2000. 19 с.
Напреенко М.Г., Напреенко-Дорохова Т.В. История формирования болотной растительности в дельтовом ландшафте реки Неман в голоцене // Вестник Балтийского федерального университета им. И. Канта. Серия “Естественные и медицинские науки”. 2018. № 2. С. 62–72.
Напреенко-Дорохова Т.В., Напреенко М.Г., Субетто Д.А. История развития природных экосистем в центральной части Калининградской области в связи с изменениями общегеографической обстановки и деятельностью человека // Общество. Среда. Развитие. 2016. № 2. С. 101–109.
Наставления гидрометеорологическим станциям и постам. Вып. 6. Ч. II. Гидрологические наблюдения и работы на малых реках. Л.: Гидрометеоиздат, 1972. 266 с.
Никонов В.В., Лукина Н.В., Безель В.С. Рассеянные элементы в бореальных лесах. М.: Наука, 2004. 616 с.
Новенко Е.Ю. Растительность и климат Центральной и Восточной Европы в позднем плейстоцене и голоцене: Дис. … д-ра геогр. наук (спец. 25.00.25). М.: МГУ им. М.В. Ломоносова, 2015. 322 с.
Новенко Е.Ю., Мироненко И.В., Волкова Е.М., Куприянов Д.А., Батанова А.К. Динамика ландшафтов Юго-восточной Мещеры в голоцене // Вестник Московского университета. Серия 5: География. 2016. № 2. С. 91–102.
Охрана природы. Городские экосистемы. Расчет величин критических нагрузок поллютантов на городские экосистемы: Методические рекомендации / разраб.: В.Н. Башкин, А.С. Курбатова, Д.С. Савин. М.–Смоленск: Маждента, 2004. 59 с.
Перельман А.И., Касимов Н.С. Геохимия ландшафта. М.: Астрея-2000, 1999. 768 с.
ПНД Ф 16.1:2.2:2.3:3.36-2002 Методика измерений валового содержания кадмия, кобальта, марганца, меди, никеля, свинца, хрома и цинка в почвах, донных отложениях, осадках сточных вод и отходах методом пламенной атомно-абсорбционной спектрометрии. М.: Изд-во ФГБУ “ФЦАО”, 2002. 22 с.
Природа Рязанской области / под ред. Кривцова В.А. Рязань: Рязанский гос. университет им. С.А. Есенина, 2008. 407 с.
Продуктивность экосистем Северной Евразии [Электронный ресурс] // База данных. 2002–2006. – URL: http://biodat.ru/db/prod/index.htm (дата обращения 29.09.2017).
Ремезов Н.П., Быкова Л.Н., Смирнова К.М. Потребление и круговорот азота и зольных элементов в лесах Европейской части СССР. М.: Изд-во МГУ, 1959. 284 с.
Серегин И.В. Распределение тяжелых металлов в растениях и их действие на рост: Дис. … д-ра биол. наук (спец. 03.00.12). М.: Институт физиологии растений им. К.А. Тимирязева РАН, 2009. 333 с.
Сибиркина А.Р. Биогеохимическая оценка содержания тяжелых металлов в сосновых борах Семипалатинского Прииртышья: Дис. … д-ра биол. наук (спец. 03.02.08). Омск: Челябинский гос. университет, 2014. 496 с.
Тобратов С.А., Железнова О.С., Водорезов А.В. Оценка пространственных закономерностей лесорастительного потенциала территории на основе “коэффициентов благоприятности” (на примере смешанных лесов Восточно-Европейской равнины) // Восточно-Европейский научный журн. 2016. Т. 8. № 5. С. 127–133.
Усольцев В.А. Фитомасса лесов Северной Евразии: нормативы и элементы географии. Екатеринбург: Изд-во УрО РАН, 2002. 762 с.
Учватов В.П. Природные и антропогенные потоки вещества в ландшафтах Русской равнины: Дис. … д-ра биол. наук (спец. 04.00.03). М.: МГУ им. М.В. Ломоносова, 1994. 471 с.
Черненькова Т.В. Закономерности аккумуляции тяжелых металлов сосной обыкновенной в фоновых и техногенных местообитаниях // Лесоведение. 2004. № 2. С. 25–35.
Belleghem F.V., Cuypers A., Semane B., Smeets K., Vangronsveld J., d’Haen J., Valcke R. Subcellular localization of cadmium in roots and leaves of Arabidopsis thaliana // New Phytologist. 2007. V. 173. № 3. P. 495–508.
Bouain N., Shahzad Z., Rouached A., Khan G.A., Berthomieu P., Abdelly C., Poirier Y., Rouached H. Phosphate and zinc transport and signalling in plants: toward a better understanding of their homeostasis interaction // J. Experimental Botany. 2014. V. 65. № 20. P. 5725–5741.
Burkhead J.L., Gogolin Reynolds K.A., Abdel-Ghany S.E., Cohu C.M., Pilon M. Copper homeostasis // New Phytologist. 2009. V. 182. № 4. P. 799–816.
Capuana M. Heavy metals and woody plants – biotechnologies for phytoremediation // iForest – Biogeosciences & Forestry. 2011. V. 4. № 1. P. 7–15.
Cocozza C., Minnocci A., Tognetti R., Iori V., Zacchini M., Scarascia-Mugnozza G. Distribution and concentration of cadmium in root tissue of Populus alba determined by scanning electron microscopy and energy-dispersive x-ray microanalysis // iForest – Biogeosciences & Forestry. 2008. V. 1. № 1. P. 96–103.
Conn S., Gilliham M. Comparative physiology of elemental distributions in plants // Annals of Botany. 2010. V. 105. № 7. P. 1081–1102.
Di Baccio D., Tognetti R., Minnocci A., Sebastiani L. Responses of the Populus × euramericana clone I-214 to excess zinc: Carbon assimilation, structural modifications, metal distribution and cellular localization // Environmental & Experimental Botany. 2009. V. 67. № 1. P. 153–163.
Ernst W.H.O. Evolution of metal tolerance in higher plants // Forest Snow & Landscape Research. 2006. V. 80. № 3. P. 251–274.
Hazama K., Nagata S., Fujimori T., Yanagisawa S., Yoeneyama T. Concentrations of metals and potential metal-binding compounds and speciation of Cd, Zn and Cu in phloem and xylem saps from castor bean plants (Ricinus communis) treated with four levels of cadmium // Physiologia Plantarum. 2015. V. 154. № 2. P. 243–255.
Huang J.-H., Ilgen G., Matzner E. Fluxes and budgets of Cd, Zn, Cu, Cr and Ni in a remote forested catchment in Germany // Biogeochemistry. 2011. V. 103. № 1–3. P. 59–70.
Maestri E., Marmiroli M., Visioli G., Marmiroli N. Metal tolerance and hyperaccumulation: Costs and trade-offs between traits and environment // Environmental & Experimental Botany. 2010. V. 68. № 1. P. 1–13.
Manual on methodologies and criteria for modeling and mapping critical loads & levels and air pollution effects, risks and trends [Электронный ресурс] / UNECE Convention on Long-Range Transboundary Air Pollution. 2004. http://www.icpmapping.org/ (дата обращения 25.09.2015).
Marschner’s mineral nutrition of higher plants: 3rd ed. / Ed. P. Marschner. Adelaide: School of Agriculture, Food and Wine, the University of Adelaide Australia, 2012. 672 p.
Österås A.H. Interactions between calcium and heavy metals in Norway spruce. Accumulation and binding of metals in wood and bark: dissertation. Stockholm: Department of Botany, Stockholm University, 2004. 52 p.
Sinclair S.A., Kramer U. The zinc homeostasis network of land plants // Biochimica et Biophysica Acta (BBA) – Molecular Cell Research. 2012. V. 1823. № 9. P. 1553–1567.
Singh S., Parihar P., Singh R., Singh V.P., Prasad S.M. Heavy metal tolerance in plants: role of transcriptomics, proteomics, metabolomics, and ionomics // Frontiers in Plant Science. 2016. V. 6. № 1143.
Sofo A., Scopa A., Remans T., Vangronsveld J., Cuypers A. Biochemical and functional responses of Arabidopsis thaliana exposed to cadmium, copper and zinc // The plant family Brassicaceae: contribution towards phytoremediation, 2012. V. 21. P. 239–263.
Vandecasteele B., Quataert P., Piesschaert F., Lettens S., De Vos B., Du Laing G. Translocation of Cd and Mn from bark to leaves in willows on contaminated sediments: Delayed budburst is related to high Mn concentrations // Land. 2015. V. 4. № 2. P. 255–280.
Waters B.M., Grusak M.A. Whole-plant mineral partitioning throughout the life cycle in Arabidopsis thaliana ecotypes Columbia, Landsberg erecta, Cape Verde Islands, and the mutant line ysl1ysl3 // New Phytologist. 2008. V. 177. № 2. P. 389–405.
Дополнительные материалы отсутствуют.