

Лесоведение, 2021, № 1, стр. 65-77
Свойства лесных подстилок на ранних этапах естественного лесовозобновления после сплошных рубок в средней тайге Карелии
В. А. Долгая a, *, О. Н. Бахмет a
a Отдел комплексных научных исследований КарНЦ РАН, ФИЦ “Карельский научный центр РАН”
185910 Петрозаводск, ул. Пушкинская, д. 11, Россия
* E-mail: dolgaya.veronika@gmail.com
Поступила в редакцию 20.03.2019
После доработки 20.01.2020
Принята к публикации 07.10.2020
Аннотация
Актуальность исследования обоснована необходимостью изучения основных закономерностей изменения свойств лесных подстилок альфегумусовых почв, формирующихся на различных почвообразующих породах, на ранних стадиях лесовозобновления после проведения сплошных рубок сосновых древостоев. Объектами исследования послужили два хронологических ряда вырубок (текущего года, 3- и 8-летней давности). Почвы пробных площадей представлены альфегумусовыми почвами – подбурами и подзолами. Установлено, что на подбурах, формирующихся на глинистых сланцах, лесовозобновление протекает через поселение лиственных пород, в то время как на подзолах, формирующихся на флювиогляциальных песках, возобновляется сосной. В живом напочвенном покрове увеличивается доля травянистых растений: на подбурах – по истечении 3-летнего периода, на подзолах – спустя 8 лет. В ходе исследования выявлено, что применение многооперационных машин способствует сохранению живого напочвенного покрова и лесных подстилок на значительной части территории вырубок. Воздействие лесозаготовительной техники в большей степени отразилось на свойствах органогенных горизонтов подбуров, мощность и запасы которых варьировали в более широких диапазонах относительно подстилок, формирующихся на подзолах. С появлением лиственных пород и травянистых растений в компонентном составе опада возросла доля быстро разлагающихся фракций, что отразилось на свойствах лесных подстилок. Наименьшие значения мощности и запасов органогенных горизонтов, содержания в них лигнина и целлюлозы, а также сужение отношения пассивных и активных фракций отмечены в подстилках подбуров на вырубке 3-летней давности и под производными древостоями 8-летнего возраста, произрастающих на подзолах. Данные показатели свидетельствуют о более интенсивной трансформации органического материала в лесных подстилках подбуров, чем в органогенных горизонтах подзолов. Таким образом, улучшение условий для трансформации органического материала наблюдается с появлением в напочвенном покрове лиственных пород и травянистых растений, жизнеспособность которых во многом зависит от обогащенности почв органическим веществом.
На протяжении последних десятилетий интенсивность освоения природных ресурсов значительно возросла. Под влиянием антропогенной нагрузки происходит деградация наземных экосистем и, как следствие – нарушение баланса углерода (Bonan, 2008; Pan et al., 2011). В контексте данной проблемы особый интерес вызывает исследование лесных экосистем бореальной зоны, поскольку в фитомассе и почвенном органическом веществе таежных лесов аккумулируется значительная часть общепланетарных запасов углерода (Kauppi, 2003; Lal, 2005; Scharlemann et al., 2014, Smyth et al., 2014). Связующим звеном между фитоценозом и почвой являются лесные подстилки, так как протекающие в них процессы гумификации и минерализации органического материала в значительной степени определяют направленность почвообразовательных процессов, плодородие почв и продуктивность древостоев (Карпачевский, 1988; Ильина, Сапожников, 2007). Лесные подстилки являются одним из важнейших индикаторов изменений внутри лесных биогеоценозов, поскольку их структура и свойства находятся в прямой зависимости от видового разнообразия и пространственной структуры экосистемы (Рыжова, 2008; Решетникова, 2011; Волков, 2015; Орлова и др., 2015).
Одним из наиболее существенных антропогенных факторов, оказывающих влияние на биоразнообразие и продуктивность лесных экосистем, является промышленная заготовка древесины. Степень нарушения живого напочвенного покрова, лесных подстилок и верхних почвенных горизонтов во многом зависит от выбора технологии проведения рубок и вида лесозаготовительной техники. Так, современные технологии лесозаготовительных работ предусматривают использование многооперационных машин – харвестера и форвардера, применение которых способствует сохранению лесных подстилок и верхних почвенных горизонтов на большей части территории вырубки (Григорьев и др., 2008; Ильинцев и др., 2017, 2018; Дымов, 2018).
Изучению структурно-функциональных связей между фитоценозами бореальных лесов и почвой после проведения рубок посвящено значительное количество исследовательских работ, как зарубежных, так и отечественных. Достаточно хорошо освещен вопрос изменения лесных подстилок после проведения сплошных рубок, где авторы приводят данные, согласно которым под воздействием лесозаготовительной техники происходит либо полное уничтожение органогенных горизонтов (Морозова, 1964; Егорова, Куликова, 1965; Бахмет, Медведева, 2013), либо частичное нарушение их структуры, уменьшение мощности и запасов (Ponge, 2003; Nave et al., 2010; Дымов и др., 2012; Piirainen et al., 2015; Вдовиченко и др., 2016; Вдовиченко, Бахмет, 2017; Смирнов и др., 2018). Авторы отмечают, что нарушение динамического равновесия между компонентами лесных экосистем, вызванного проведением сплошных рубок, приводит к дисбалансу между образованием почвенного органического вещества в результате разложения растительного опада и его минерализацией до неорганических соединений, доступных растениям (Федорец, 2004; Jandl et al., 2007; Дымов и др., 2012; Falsone et al., 2012; Бахмет, 2014; Лаптева и др., 2015; Clarke et al., 2015; Piirainen et al., 2015; Дымов, 2018). Так, данные, полученные в ходе исследований (Palviainen et al., 2004; Smolander, Heiskanen, 2007; Лаптева и др., 2015; Piirainen et al, 2015; Вдовиченко и др., 2016), свидетельствуют об увеличении скорости минерализации азота в лесных подстилках и верхних почвенных горизонтах на ранних этапах лесовозобновления за счет поступления на поверхность почвы большого количества легкоразлагаемого органического материала.
Проведенные ранее на территории Республики Карелии исследования (Морозова, 1964; Егорова, Куликова, 1965; Лазарева, Вуоримаа, 1986; Зябченко и др, 1988; Федорец, 2004; Бахмет, Медведева, 2013; Бахмет, 2014; Вдовиченко и др., 2016; Вдовиченко, Бахмет, 2017) также содержат информацию об изменении свойств лесных подстилок. Однако вышеперечисленные исследования затрагивают наиболее характерные для региона типы леса и почвы – сосновые и еловые насаждения, произрастающие на подзолах, формирующихся на флювиогляциальных отложениях. Изменение лесных подстилок, формирующихся на других почвообразующих породах, изучено слабо. Кроме того, не выявленным остается вопрос изменения свойств лесных подстилок после проведения рубок с использованием современных лесозаготовительных технологий.
В контексте всего вышеперечисленного, несомненно, актуальна задача выявления степени нарушения лесных подстилок в результате проведения лесозаготовительных работ и изменения их свойств при естественном восстановлении леса, поскольку полученные данные позволят оценить последствия антропогенного воздействия на бореальные биогеоценозы и дать прогноз о сроках восстановления лесных экосистем. В связи с этим цель исследования заключалась в определении морфологических и биохимических свойств лесных подстилок сосняков, произрастающих на почвах, формирующихся на различных почвообразующих породах, после проведения сплошных рубок с применением многооперационных машин.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в среднетаежной подзоне Республики Карелии на территории Кончезерского лесничества (рис. 1). Ввиду преобладания воздушных масс атлантического и арктического происхождения, близости к Балтийскому, Белому и Баренцеву морям, а также своеобразия местных природных условий (обилие озер и болот, пересеченный рельеф местности и др.) для региона характерны следующие климатические особенности: невысокая среднегодовая температура воздуха (3°С), продолжительная, но мягкая зима (число дней со снежным покровом 145–155), короткое прохладное лето (продолжительность периода со среднесуточной температурой воздуха выше 15°С – 40–45 дней), высокая относительная влажность воздуха (60–90%), значительное количество осадков (550–600 мм) (Назарова, 2003).
Для выявления закономерностей изменения лесных подстилок в процессе естественного лесовозобновления использовался подход, основанный на исследовании хронологических рядов вырубок. Объектами исследования послужили два хронологических ряда, представленные вырубками текущего года, а также производными древостоями, формирующимися на местах проведения рубок сосняков черничных 3- и 8-летней давности. В качестве контрольных участков были подобраны сосняки черничные 120-летнего возраста. Почвы первого ряда представлены подзолами иллювиально-железистыми (O–E–BF–C), формирующимися на флювиогляциальных песках, второго – подбурами грубогумусированными (O–BHF–C), формирующимися на глинистых сланцах. Характеристики почв представлены в табл. 1. Подбор пробных площадей осуществлялся с учетом однородности лесорастительных условий, включая особенности рельефа, почвообразующих пород, гидрологического режима почв и др. Фоновые участи примыкали к границе вырубок текущего года, остальные располагались в пределах одного лесного квартала.
Таблица 1.
Химические свойства исследуемых почв под ненарушенными сосновыми древостоями
| Тип почвы | Горизонт | рНсол | рНвод | V, % | P2O5, мг × 100–1 г | K2O, мг × 100–1 г | С, % | N, % | C : N |
|---|---|---|---|---|---|---|---|---|---|
| Подзол иллювиально-железистый |
O(L) | 3.1 | 4.0 | 45.6 | 54.9 | 21.3 | 40.0 | 1.20 | 33 |
| O(F+H) | 2.9 | 4.1 | 42.1 | 40.2 | 20.4 | 32.5 | 0.73 | 44 | |
| Е | 3.5 | 4.1 | 20.5 | 0.4 | 1.3 | 1.5 | 0.08 | 17 | |
| BF1 | 4.7 | 5.5 | 44.9 | 14.2 | 1.0 | 0.6 | 0.02 | 25 | |
| BF2 | 4.4 | 5.5 | 70.0 | 9.4 | 1.1 | 0.3 | 0.01 | 23 | |
| ВС | 4.3 | 5.5 | 87.7 | 9.8 | 1.2 | 0.1 | 0.01 | 11 | |
| С | 4.5 | 5.6 | 94.3 | 13.6 | 1.2 | 0.1 | 0.01 | 18 | |
| Подбур грубогумусированный | O(L) | 3.7 | 4.4 | 60.6 | 30.1 | 149.8 | 35.6 | 1.44 | 25 |
| O(F) | 3.4 | 4.2 | 58.3 | 26.3 | 125.5 | 43.3 | 1.41 | 31 | |
| O(H) | 3.2 | 4.1 | 52.1 | 20.0 | 26.8 | 12.4 | 0.38 | 32 | |
| BHF | 3.2 | 4.0 | 5.4 | 11.0 | 4.4 | 3.8 | 0.21 | 17 | |
| BF | 4.2 | 5.1 | 17.6 | 19.2 | 2.3 | 2.5 | 0.15 | 17 | |
| BC | 4.5 | 5.5 | 24.8 | 16.4 | 5.8 | 1.5 | 0.16 | 9 |
Описания древостоев и типов вырубок выполнены по общепринятым методикам научным сотрудником лаборатории динамики и продуктивности таежных лесов ФИЦ КарНЦ РАН В.А. Харитоновым, живого напочвенного покрова лесных фитоценозов – научным сотрудником лаборатории ландшафтной экологии и охраны лесных экосистем ФИЦ КарНЦ РАН В.В. Тимофеевой. Таксационные описания ненарушенных древостоев и вырубок представлены в табл. 2, 3.
Таблица 2.
Таксационная характеристика ненарушенных сосновых древостоев 120-летнего возраста
| Тип почвы | Состав древостоя | Порода | Густота, тыс. шт. га–1 | Высота, м | Диаметр, см | Запас, м3 га–1 | Полнота относительная |
Класс бонитета |
|---|---|---|---|---|---|---|---|---|
| Подзол иллювиально-железистый | 9С1Б | Сосна | 0.4 | 28.2 | 31.6 | 2966 | 0.6 | I |
| Береза | 0.03 | 20.2 | 16.8 | |||||
| Подбур грубогумусированный | 10С, ед. Б | Сосна | 1.3 | 19.7 | 18.1 | 322 | 0.9 | III |
| Береза | 0.1 | 7 | 7.1 |
Таблица 3.
Таксационная характеристика вырубок
| Тип почвы | Возраст вырубки, лет | Тип вырубки/древостоя | Состав | Густота, тыс. га–1 | Встречаемость, % | Средняя высота, м |
|---|---|---|---|---|---|---|
| Подзол иллювиально-железистый | 0 | Кустарничково-зеленомошная | Сосна Береза Ива |
1.2 0.3 1.7 |
52 12 10 |
0.29 ± 0.02 0.30 ± 0.04 0.54 ± 0.08 |
| 3 | Кустарничково-зеленомошная | Сосна Береза Осина Ива Ольха |
5.3 3.7 0.8 1.1 0.5 |
77 83 53 30 7 |
0.49 ± 0.22 1.26 ± 0.16 0.98 ± 0.30 0.69 ± 0.18 0.91 ± 0.06 |
|
| 8 | Сосняк кустарничково-разнотравный (7С3Б+Ос) | Сосна Береза Осина Ива Ольха |
5.7 1.9 0.3 0.9 0.2 |
88 39 28 28 4 |
2.12 ± 0.06 3.05 ± 0.18 1.61 ± 0.40 2.53 ± 0.21 3.42 ± 0.06 |
|
| Подбур грубогумусированный | 0 | Потенциально Вейниково-луговиковая |
Сосна Береза Ива |
0.3 0.8 1.7 |
12 8 24 |
0.42 ± 0.22 0.34 ± 0.05 0.66 ± 0.20 |
| 3 | Вейниково-луговиковая | Сосна Береза Осина Ива |
3.0 1.0 1.6 1.3 |
35 15 25 15 |
0.22 ± 0.03 0.65 ± 0.31 0.49 ± 0.06 0.89 ± 0.38 |
|
| 8 | Осинник травяно-злаковый (5Ос2С2Б1Ол, ед. Е, Ив) | Сосна Ель Береза Осина Ива Ольха |
1.8 0.4 2.5 5.2 0.2 1 |
36 16 44 52 4 16 |
2.12 ± 0.40 0.77 ± 0.14 3.67 ± 0.43 3.34 ± 0.19 1.25 ± 0.25 3.20 ± 0.33 |
На типичных участках пробных площадей закладывали полнопрофильные разрезы почв, проведено их детальное морфологическое описание. Для оценки пределов варьирования морфологических и физико-химических свойств почв на пробных площадях закладывали по 5 полнопрофильных разреза почв. Почвы диагностировали согласно классификации почв России (Шишов, Тонконогов, 2004). Для исследования лесных подстилок на каждой пробной площади в случайном порядке закладывали прикопки в 20-кратной повторности, исключая приствольные участки крупных деревьев и лесные поляны. В прикопках с использованием линейки проводили определение мощности лесных подстилок, после чего отбирали образцы рамкой площадью 0.04 м2. Детальное исследование морфологических свойств, а также расчет запасов лесных подстилок проводили согласно общепринятой методике (Шумаков, Федорова, 1979). В лабораторных условиях при исследовании компонентного состава лесные подстилки сортировали по фракциям: древесина, ветви, кора, шишки, злаки, мхи, опад кустарничков, хвоя, листья и труха. Все фракции высушивали до абсолютно-сухого состояния, с последующим взвешиванием и вычислением процентного соотношения фракций от общей массы сухого вещества. Для определения химических свойств лесных подстилок образцы, отобранные ранее на каждой пробной площади в 20‑кратной повторности, усредняли путем смешивания. На научном оборудовании Центра коллективного пользования ФИЦ КарНЦ РАН выполнено определение общего содержания углерода и азота (2400 Series II CHNS/O Elemental Analyzer (Perkin Elmer, США)), содержания лигнина (методом Класона в модификации Комарова) и целлюлозы (методом Кюршнера–Хоффера). Статистическая обработка данных проведена с использованием пакетов программ Statistica 10, Microsoft Excel 2007. Для выявления достоверности различий использовали критерий Стьюдента.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сукцессионная смена растительности после проведения сплошных рубок
Фоновые участки представлены сосновыми древостоями (Pinus sylvestris L.) с примесью березы повислой (Betula pendula Roth.). Ярус подлеска развит слабо и состоит из единичных экземпляров ели европейской (Picea abies (L.) Karst), рябины обыкновенной (Sorbus aucuparia L.), осины обыкновенной (Populus tremula L.), ивы козьей (Salix caprea L.) и можжевельника обыкновенного (Junipirus communis L.). В живом напочвенном покрове доминантами выступают черника обыкновенная (Vacinium myrtilis L.), брусника обыкновенная (Vacinium vitis-ideae L.), плевроциум Шребера (Pleurozium schreberi (Brid.) Mitt.) и гилокиум блестящий (Hylocomium splendens (Hedw.) Schimp). В большом количестве присутствуют: вереск обыкновенный (Calluna vulgaris (L.) Hull), ожика волосистая (Lusula pilosa (L.) Willd.), седмичник европейский (Trientalis europaea L.), марьянник луговой (Melampyrum pratense L.) и дикранум метловидный (Dicranum scoparium Hedw.).
Несмотря на равные экологические условия, для фоновых участков характерны некоторые различия. В подлеске, формирующемся на подзоле, преобладает ель европейская и можжевельник обыкновенный, в то время как на подбуре – рябина обыкновенная и шиповник иглистый (Rosa acicularis Lindl). В живом напочвенном покрове первого экотопа присутствуют хвощ зимующий (Equisetum hyemale L.) и лесной (Equisetum sylvaticum L.), гудайера ползучая (Goodyera repens (L.) R. Br.), майник двулистный (Maianthemum bifolium (L.) F. W.), кладония оленья (Cladonia rangiferina (L.) F. H. Wigg.) и цетрария исландская (Cetraria islandica (L.) Ach.). Во втором экотопе в синузии трав встречаются достаточно редкие виды: короставник полевой (Knautia arvensis (L.) Coult.), линнея северная (Linnaea borealis L.), плаун годичный (Lycopodium annotinum L.).
Применение многооперационных машин при проведении лесозаготовительных работ способствует сохранению живого напочвенного покрова и органогенных горизонтов на значительной части территории вырубки. На свежей вырубке первого экотопа наблюдается большое число всходов сосны, подроста березы и ивы, а также сохранность кустарничново-зеленомошного покрова, в то время как численность видов мохово-лишайникового яруса значительно сокращается. По истечении 3-летнего периода после рубки древостоев происходит зарастание территории травяной растительностью, среди которой преобладает иван-чай узколистный (Chamaenerion angustifolium (L.) Scop.) и вейник наземный (Calamagrostis epigeios (L.) Roth). Несмотря на обилие трав сплошного задернения территории не происходит, и в напочвенном покрове по-прежнему преобладают светолюбивые кустарнички – черника, брусника, вереск. В мохово-лишайниковом покрове восстанавливается кладония и цетрария исландская. При достижении 8-летнего возраста древесный ярус представлен смешанным древостоем сосны и березы, осина встречается в единичных экземплярах. В живом напочвенном покрове преобладают вейник наземный и иван-чай узколистный, появляется большое количество трав: тысячелистник обыкновенный (Achillea millefolium L.), полевица тонкая (Agrostis tenuis Sibth.), дудник лесной (Angelica sylvestris L.), душистый колосок обыкновенный (Anthoxantum odoratum L.), купырь лесной (Anthriscus sylvestris (L.) Hoffm.), толокнянка обыкновенная (Arctostaphylos uva-ursi (L.) Spreng.), вероника лекарственная (Veronica officinalis L.). Значительно сокращается доля дикранума и кладонии, однако в большом количестве появляется новый вид – полия поникшая (Pоhlia nutans (Hedw.) Lindb.).
На подбурах восстановление в первые три года после проведения рубок происходит преимущественно лиственными древесными породами – березой и осиной, всходов сосны значительно меньше. В живом напочвенном покрове чаще других доминируют вейник тростникововидный, ландыш лесной (Convallaria majalis L.), также присутствуют луговик извилистый (Deschampsia flexuosa (L.) Trin) и иван-чай. Доля мохово-кустарничкового яруса сокращается значительно, из видов остаются только плевроциум Шребера и гилокиум блестящий. При достижении 8-летнего возраста древостоев на территории вырубки преобладающей породой становится осина, в значительно меньшей степени присутствуют сосна обыкновенная и береза. Сдерживающим фактором роста и развития осины является зоогенный фактор – ветви молодых деревьев объедают лоси. В живом напочвенном покрове господствует травяная растительность: тысячелистник обыкновенный, дудник лесной (Angelica sylvestris L.), луговик извилистый, марьянник луговой (Melampyrum pratense L.), вейник тростникововидный, ландыш майский. Моховой покров восстанавливается медленно, возобновление хвойных пород затруднено.
Таким образом, можно сделать вывод, что на относительно бедных органическим веществом подзолах возобновление древесной растительности происходит преимущественно сосной с примесью березы, в то время как на подбурах – лиственными породами.
Мощность и запас лесных подстилок
Сукцессионная смена растительности сопровождается изменением количественного и качественного состава поступающего на поверхность почвы растительного опада, и, как следствие, отражается на структуре и свойствах лесных подстилок Основными показателями, характеризующими лесную подстилку, являются мощность и запас, а также агрохимические свойства: реакция среды, содержание углерода, питательных элементов и т.д. (Карпачевский, 1977; Шумаков, 1979). Показатели мощности и запасов позволяют оценить скорость разложения органических остатков.
Под сосняками черничными органогенные горизонты представляют собой типичные для данного типа леса и климатических особенностей региона грубогумусные лесные подстилки, дифференцируемые на подгоризонты L (листовой), F (ферментации) и H (гумификации). Однако из-за различия экотопов формирование органогенных горизонтов на пробных площадях различается. Лесная подстилка, формирующаяся под сосняком, произрастающим на подзоле, стратифицируется только на 2 подгоризонта L и F + H, мощность которых составляет 1.4 ± 0.4 и 1.8 ± 0.5 см, соответственно. Органогенный горизонт сосняка, произрастающего на подбуре, имеет иную стратификацию – три подгоризонта L, F, H. Мощность подгоризонтов варьирует в диапазонах: 2.1 ± 0.7 см – для L; 3.9 ± 0.4 см – для F, 3.4 ± 0.6 см – для H. Аккумуляция органических остатков на поверхности подбуров связана с большей увлажненностью данных участков (в среднем 67% – в L, 53% – F, 46% – H), в отличие от органогенных горизонтов подзола (в среднем 26% – в L, 32% – F + H).
Проведение лесозаготовительных работ сопровождалось нарушением структуры лесных подстилок, на что указывает изменение мощности органогенных горизонтов (2.2 ± 1.2 см – на подзоле, 2.8 ± 1.9 см – на подбуре). Таким образом, степень нарушения лесных подстилок подзолов меньше, чем рыхлых органогенных горизонтов более каменистых подбуров, где под воздействием лесозаготовительной техники наблюдается сильное перемешивание и уплотнение органогенных горизонтов.
По истечении 3-летнего периода после проведения лесозаготовительных работ мощность лесных подстилок подзолов относительно свежей вырубки изменилась незначительно – до 2.3 ± 1.3 см, что обусловлено сохранностью зеленомошного напочвенного покрова и наличием слаборазложившихся компонентов древесного опада и порубочных остатков. В лесных подстилках подбуров напротив наблюдается снижение мощности до 2.3 ± ± 1.8 см. Данные изменения вызваны увеличением доли травянистой растительности в составе живого напочвенного покрова и улучшением условий для минерализации растительного материала.
К моменту смыкания древесного полога в возрасте 8 лет наблюдается уменьшение мощности лесных подстилок подзолов (2.1 ± 0.5 см) за счет изменения ботанического состава, в составе которого увеличилась доля травянистых растений. На подбурах сохраняются благоприятные условия для трансформации органического материала, что влечет за собой снижение показателей мощности органогенных горизонтов (2.2 ± 0.6 см).
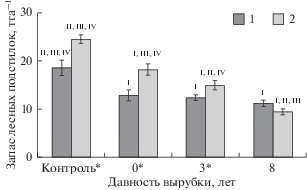
Изменение мощности подстилок в значительной степени отражается на запасах (рис. 2). По данным статистического анализа выявлено, что распределение запасов лесной подстилки в исследуемых образцах на 43% определено возрастом вырубки и на 5% – совместным действием двух факторов – типом почвы и возрастом вырубки. Статистически достоверные различия запасов лесных подстилок на подзолах и подбурах отмечены на контрольных участках и на вырубках текущего года и 3-летней давности. На подзолах статистически достоверные различия между стадиями восстановления отмечены только между контрольным участком и свежей вырубкой, в то время как на подбурах – между всеми стадиями восстановления. Исходя из полученных данных, можно сделать вывод, что интенсивность трансформации органических остатков в лесных подстилках подбуров протекает быстрее, чем в органогенных горизонтах подзолов.
Рис. 2.
Распределение запасов лесных подстилок, формирующихся на подзолах (1) и подбурах (2), на различных стадиях естественного лесовозобновления. На диаграмме указаны средние значения и их стандартные ошибки (N = 20). Статистически достоверные различия показателей значимы при P ≤ 0.05 – для разных типов почв (*) и на разных стадиях сукцессий (I – контроль, II – свежая вырубка, III – 3-летняя вырубка, IV – 8-летняяя вырубка).
Проведенные ранее исследования (Бахмет, Медведева, 2013) свидетельствуют, что при механизированных посадках лесных культур, наблюдаются значительные нарушения органогенных и верхних минеральных горизонтов подзолов иллювиально-железистых. Авторы отмечают, что с увеличением давности рубок постепенно возрастает мощность и запасы органогенных горизонтов, значения которых варьируют в более широких диапазонах относительно почв вырубок, находящихся в процессе естественного лесовозобновления.
Компонентный состав лесных подстилок
Для лесных подстилок фоновых участков характерно преобладание активных фракций над пассивными, что обусловлено преобладанием хорошо трансформированных растительных остатков в подгоризонтах F и H (F + H) над слабо трансформированным материалом в горизонте L. Однако в компонентном составе подстилок сосняка, произрастающего на подзоле (табл. 4), содержание пассивных фракций значительно больше, чем в органогенных горизонтах подбуров (табл. 5). Данные различия, вероятнее всего, обусловлены степенью увлажнения почв, поскольку в отсутствии достаточного увлажнения увеличивается опад древесных пород.
Таблица 4.
Содержание различных компонентов в лесных подстилках, формирующихся на подзолах (% от сухой массы)
| Фракция | Контроль | Свежая | 3 года | 8 лет |
|---|---|---|---|---|
| Пассивные фракции | ||||
| Древесина | 6.3 ± 2.7 | 11.5 ± 5.5* | 9.7 ± 5.5 | 7.6 ± 2.0 |
| Ветви | 6.5 ± 2.2 | 9.0 ± 3.0* | 7.6 ± 2.6 | 5.2 ± 1.8* |
| Кора | 4.5 ± 1.9 | 9.0 ± 2.4* | 8.1 ± 2.6 | 5.5 ± 1.5* |
| Шишки | 7.9 ± 2.4 | 10.3 ± 6.3 | 9.0 ± 2.4 | 4.2 ± 1.1* |
| Активные фракции | ||||
| Злаки | 1.1 ± 0.7 | 0.9 ± 0.4 | 3.8 ± 1.1* | 4.0 ± 0.9* |
| Мхи | 7.2 ± 2.0 | 4.5 ± 0.6* | 3.2 ± 1.7 | 3.0 ± 1.3* |
| Опад кустарничков | 3.6 ± 1.7 | 2.6 ± 0.9* | 3.4 ± 1.5 | 2.9 ± 1.1 |
| Хвоя | 6.7 ± 1.8 | 10.5 ± 2.9* | 6.6 ± 2.3* | 8.0 ± 1.6 |
| Листья | 1.1 ± 0.7 | 2.6 ± 0.9* | 2.5 ± 0.9 | 4.5 ± 0.9* |
| Труха | 54.9 ± 7.4 | 39.1 ± 19.4* | 46.2 ± 17.8 | 55.1 ± 14.0* |
| Отношение пассивных фракций к активным | 25.2/74.8 | 39.8*/60.2* | 34.3/65.7 | 22.5*/77.5* |
Таблица 5.
Содержание различных компонентов в лесных подстилках, формирующихся на подбурах (% от сухой массы)
| Фракция | Контроль | Свежая | 3 года | 8 лет |
|---|---|---|---|---|
| Пассивные фракции | ||||
| Древесина | 3.5 ± 1.1 | 10.1 ± 5.8* | 8.4 ± 3.4 | 2.0 ± 0.6* |
| Ветви | 4.3 ± 1.0 | 8.6 ± 5.3* | 5.5 ± 2.0 | 4.8 ± 1.3* |
| Кора | 5.1 ± 1.5 | 10.4 ± 7.6* | 9.0 ± 3.2 | 5.1 ± 2.0* |
| Шишки | 5.0 ± 1.9 | 6.2 ± 1.9 | 5.0 ± 1.5 | 1.0 ± 0.3* |
| Активные фракции | ||||
| Злаки | 2.3 ± 0.7 | 3.5 ± 1.2* | 5.5 ± 1.0* | 8.2 ± 4.1* |
| Мхи | 7.3 ± 1.8 | 3.0 ± 0.9* | 1.1 ± 0.4* | 5.8 ± 1.1* |
| Опад кустарничков | 2.5 ± 0.6 | 1.7 ± 0.6* | 1.5 ± 0.9 | 2.6 ± 0.7* |
| Хвоя | 6.7 ± 2.0 | 9.6 ± 2.9* | 5.5 ± 2.6* | 3.5 ± 1.0* |
| Листья | 3.0 ± 1.3 | 3.0 ± 0.9 | 2.8 ± 0.8 | 5.9 ± 1.6* |
| Труха | 60.5 ± 18.1 | 44.1 ± 14.5* | 55.9 ± 12.2* | 61.1 ± 12.3 |
| Отношение пассивных фракций к активным | 17.8/85.2 | 35.2*/64.8* | 26.8*/73.2* | 13.0*/87.0* |
В ходе проведения лесозаготовительных работ на поверхность почвы поступает значительное количество порубочных остатков (древесина > > кора > ветви > шишки), что влечет за собой увеличение доли пассивных фракций в компонентном составе лесных подстилок исследуемых почв. По истечении 3-летнего периода с момента проведения рубок изменение компонентного состава органогенных горизонтов подзолов не происходит, в то время как в составе лесных подстилок подбуров отмечается увеличение активных фракций, что обусловлено появлением в напочвенном покрове травянистых растений. Из порубочных и древесных остатков лучше разлагается хвоя и ветви, в то время как потери по массе коры и древесины минимальны. При достижении 8-летнего возраста древостоев в компонентном составе подстилок возрастает доля активных фракций, во-первых, вследствие увеличения фракции трухи, вызванной разложением древесных и порубочных остатков, во-вторых – в связи с поступлением свежего опада. В первом экотопе накопление происходит преимущественно за счет опада хвойных пород, в то время как во втором – за счет растительных остатков лиственных пород и злаков. В целом в условиях высокой каменистости и влажности трансформация органических остатков протекает быстрее. В лесных подстилках подбуров увеличение фракции трухи идет интенсивнее, чем в подзолах.
Данные компонентного состава лесных подстилок подзолов фоновых участков хорошо сопоставимы с результатами исследований, проводимых ранее в Карелии (Морозова, Федорец, 1992; Бахмет, 1999), однако полученные нами данные имеют некоторые отличия. Так, по результатам нашего исследования доля хорошо разложившихся фракций в подгоризонтах подстилок оцениваются в 56.9%, в то время как у других авторов среднем составляет 65–70%. Кроме того, полученные ранее другими авторами данные по содержанию фракций хвои (4–6%), веток (3–6%) и шишек (4–5%) несколько ниже, чем полученные в ходе нашего исследования.
Биохимический состав лесных подстилок
Поступающий на поверхность почвы органический материал подвергается процессам минерализации и гумификации. О скорости минерализации можно судить по отношению общего углерода к азоту (C : N). Для лесных подстилок ненарушенных сосновых древостоев и ранних этапов возобновления отношение азота к углероду указывает на невысокую степень минерализации органического вещества. В целом показатели варьируют в пределах 30–45%, однако значения в первом сукцессионном ряду несколько выше, чем во втором. При достижении 8-летнего возраста наблюдается улучшение условий минерализации, отношение C : N в лесных подстилках подзолов сужается до 28, подбуров – до 20. Изменения происходят за счет увеличения микробиологической активности, которая сопровождается интенсивным потреблением азота.
Оценить степень гумификации органического вещества в лесных подстилках позволяет содержание целлюлозы и лигнина, поскольку данные компоненты обладают относительной устойчивостью к разложению и являются основным источником образования гумусовых кислот. В ходе исследования выявлено, что на контрольных участках в органогенных горизонтах подзолов доля целлюлозы и лигнина ниже, что свидетельствует о более интенсивной трансформации органических остатков в подстилках первого экотопа.
Как было отмечено выше, проведение лесозаготовительных работ в значительной степени повлияло на структуру лесных подстилок, что, в свою очередь, отразилось на содержании целлюлозы и лигнина, показатели которых несколько ниже данных контрольных участков (рис. 3, 4). По истечении 3-летнего периода в органогенных горизонтах подбуров наблюдается резкое снижение показателей, что указывает на более интенсивную трансформацию органических остатков по сравнению с показателями органогенных горизонтов подзолов. На данном этапе различия в скорости трансформации растительных остатков в лесных подстилках обусловлены увеличением в компонентном составе органогенных горизонтов опада живого напочвенного покрова и лиственных пород. По истечении 8-летнего периода различия показателей в составе лесных подстилок подзолов и подбуров нивелируются и скорость трансформации органического вещества уравнивается.
Рис. 3.
Содержание целлюлозы в лесных подстилках, формирующихся на подзолах (1) и подбурах (2), на различных стадиях естественного лесовозобновления. На диаграмме указаны средние значения и их стандартные ошибки (N = 20). Статистически достоверные различия показателей значимы при P ≤ 0.05 – для разных типов почв (*) и на разных стадиях сукцессий (I – контроль, II – свежая вырубка, III – 3‑летняя вырубка, IV – 8-летняяя вырубка).
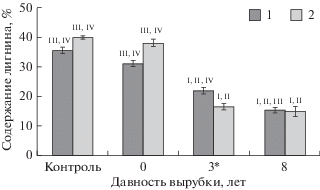
Рис. 4.
Содержание лигнина в лесных подстилках, формирующихся на подзолах (1) и подбурах (2), на различных стадиях естественного лесовозобновления. На диаграмме указаны средние значения и их стандартные ошибки (N = 20). Статистически достоверные различия показателей значимы при P ≤ 0.05 – для разных типов почв (*) и на разных стадиях сукцессий (I – контроль, II – свежая вырубка, III – 3‑летняя вырубка, IV – 8-летняяя вырубка).
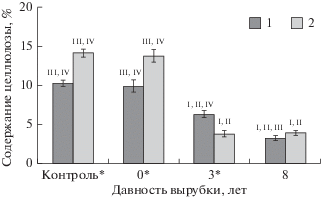
Таким образом, в лесных подстилках подбуров наилучшие условия гумификации отмечены на вырубке 3-летней давности, в органогенных горизонтах подзолов – под смешанным древостоем 8-летнего возраста. По данным статистического анализа установлено, что содержание лигнина в исследуемых образцах на 84% определено возрастом вырубки и на 5% – совместным действием двух факторов (тип почвы и давность вырубки), в то время как содержание целлюлозы – на 82 и 9%, соответственно.
Полученные ранее сведения об изменении содержания целлюлозы и лигнина в лесных подстилках подзолов в процессе искусственного лесовосстановления (Бахмет, Медведева, 2013) показывают, что после проведения лесозаготовительных работ процессы трансформации органического вещества заторможены. Установлено, что минимальные значения данных компонентов отмечены в органогенных горизонтах 7-летних культур. Можно сделать вывод, что темпы восстановления лесных подстилок на ранних этапах лесовозобновления при естественном и искусственном восстановлении совпадают.
ЗАКЛЮЧЕНИЕ
В результате проведенных исследований получены новые данные о свойствах лесных подстилок после проведения сплошных рубок древостоев с применением многооперационных машин и на различных стадиях естественного лесовозобновления. Установлено, что естественное возобновление древесных пород на альфегумусовых почвах, формирующихся на песчаных отложениях и глинистых сланцах, протекает различно. На подзолах иллювиально-железистых песчаных формируются смешанные сосновые древостои с большой примесью березы, в живом напочвенном покрове быстро восстанавливается покрытие мхов, в травяно-кустарничковом ярусе преобладают брусника, вереск и майник. На ранних этапах лесовозобновления на подбурах формируются преимущественно лиственные насаждения, представленных осиной и березой. В живом напочвенном покрове доминируют вейник лесной и малина, моховой покров восстанавливается медленно, возобновление хвойных пород затруднено.
Формирование лесных подстилок, их структура и свойства в значительной степени зависят от видового состава и продуктивности древостоев. Показатели лесных подстилок фоновых участков хорошо коррелируют с данными, полученными другими авторами ранее. После проведения лесозаготовительных работ радикального изменения морфологического строения органогенных горизонтов на большей части территории вырубки не происходит. При этом в местах воздействия лесозаготовительной техники мощность и запасы лесных подстилок подбура варьировали в более широких диапазонах относительно органогенных горизонтов подзола, что указывает на большую степень нарушения структуры лесных подстилок второго экотопа. Наименьшие значения вышеперечисленных показателей отмечены под производным древостоем 8-летнего возраста, произрастающего на подзоле, в то время как на подбуре – под вырубкой 3-летней давности.
В ходе проведения лесозаготовительных работ на поверхность почвы поступает значительное количество порубочных остатков, что влечет за собой увеличение пассивных фракций в компонентном составе органогенных горизонтов исследуемых почв. На всех этапах послерубочных сукцессий наблюдается сужение отношения пассивных и активных фракций в компонентном составе лесных подстилок, как подзолов, так и подбуров. Данные изменения вызваны, во-первых, увеличением количества легкоразлагаемых фракций лиственных пород и травянистых растений, во-вторых, – разложением древесных и порубочных остатков, сопровождающихся образованием трухи.
Трансформация органического материала в лесных подстилках сопровождается изменением биохимического состава органического вещества. После проведения лесозаготовительных работ содержание лигнина и целлюлозы в органогенных горизонтах свежих вырубок относительно контрольных участков не наблюдается. С увеличением периода после проведения рубок уменьшение содержания лигнина и целлюлозы в лесных подстилках подзолов протекает линейно, в то время как в органогенных горизонтах подбуров наблюдается резкая деградация компонентов в первые три года после проведения рубок. По истечении 8-летнего периода различия показателей в составе лесных подстилок подзолов и подбуров нивелируются, и скорость трансформации органического вещества уравнивается.
Таким образом, улучшение условий для трансформации органического материала наблюдается с появлением в напочвенном покрове лиственных пород и травянистых растений, жизнеспособность которых во многом зависит от обогащенности почв органическим веществом.
Сравнение морфологических и биохимических свойств лесных подстилок подзолов на ранних этапах естественного и искусственного возобновления свидетельствует, что темпы восстановления органогенных горизонтов совпадают. Так, наибольшая трансформация органического вещества лесных подстилок подзолов происходит примерно в один и тот же период – 7-летний возраст культур и 8-летний возраст производных древостоев, в то время как в лесных подстилках подбуров наибольшая интенсивность процессов гумификации и минерализации органического вещества наблюдается при достижении древостоями 3-летнего возраста.
Список литературы
Бахмет О.Н. Органопрофили лесных почв Карелии: Дис. … канд. биол. наук 03.00.27. М., 1999. 138 с.
Бахмет О.Н. Структурно-функциональная организация органопрофилей почв лесных экосистем Северо-Запада России: Дис. … д-ра. биол. наук 03.00.27. Петрозаводск, 2014. 350 с.
Бахмет О.Н., Медведева М.В. Изменение свойств почв Карелии в процессе искусственного лесовосстановления // Лесоведение. 2013. № 3. С. 38–45.
Вдовиченко В.А., Бахмет О.Н. Влияние сплошных рубок леса на трансформацию органического вещества почв Карелии // Известия Уфимского НЦ РАН. 2017. № 3(1). С. 41–44.
Вдовиченко В.А., Бахмет О.Н., Ткаченко Ю.Н. Начальные этапы восстановления почв на коренных горных породах после рубок сосновых древостоев // Ученые записки Петрозаводского государственного университета. 2016. № 6(159). С. 37–41.
Волков А.Г. Пространственная структура лесной подстилки в еловых экосистемах северной подзоны тайги: Дис. … канд. биол. наук 03.02.08, Архангельск, 2015. 130 с.
Григорьев И.В., Жукова А.И., Григорьева О.И., Иванов А.В. Средощадящие технологии разработки лесосек в условиях Северо-Западного региона Российской Федерации. Санкт-Петербург: Изд-во Лесотехнической академии, 2008. 176 с.
Дымов А.А. Почвы послерубочных, постпирогенных и постагрогенных лесных экосистем Северо-Востока Европейской части России: Дис. … д-ра. биол. наук 03.02.13, 03.02.08. М., 2018. 286 с.
Дымов А.А., Милановский Е.Ю., Лаптева Е.М. Изменение почв и почвенного органического вещества в процессе естественного лесовозобновления после рубки сосняка бруснично-зеленомошного // Вестник МГУЛ – Лесной вестник. 2012. № 2. С. 67–72.
Егорова Н.В., Куликова В.К. Влияние различных способов механизированных рубок леса на плодородие почв Южной Карелии // Плодородие почв Карелии. М.: Наука. 1965. С. 144–158.
Зябченко С.С., Загуральская Л.М., Лазарева И.П. Динамика экологических процессов на сплошных концентрированных вырубках северной Карелии // Лесоведение. 1988. №3. С. 3–10.
Ильина Т.М., Сапожников А.П. Лесные подстилки как компонент лесного биогеоценоза // Вестник КрасГАУ. 2007. № 5. С. 45–48.
Ильинцев А.С., Быков Ю.С., Солдатова Д.Н., Богданов А.П., Ершов Р.А. Воздействие современной лесозаготовительной техники на физические свойства почвы в северной тайге Архангельской области // Антропогенная трансформация природной среды. 2018. № 4. С. 153–155.
Ильинцев А.С., Третьяков С.В., Наквасина Е.Н., Амосова И.Б., Алейников А.А., Богданов А.П. Влияние длительно-постепенных рубок в смешанных сосновых насаждениях на естественное лесовозобновление, живой напочвенный покров и некоторые свойства верхних горизонтов почвы // Лесотехнический журн. 2017. Т. 7. № 3(27). С. 85–99.
Карпачевский Л.О. Подстилка – особый биогоризонт лесного биогеоценоза // Роль подстилки в лесных биогеоценозах. Москва: Наука, 1988. С. 88–89.
Лазарева И.П., Вуоримаа Т.А. Влияние сплошных рубок леса на свойства песчаных подзолов Северной Карелии // Лесоводственные и экологические последствия рубок в лесах Карелии. Петрозаводск: КФ АН СССР, 1986. С. 61–79.
Лаптева Е.М., Втюрин Г.М., Бобкова К.С., Каверин Д.А., Дымов А.А., Симонов Г.А. Изменение почв и почвенного покрова еловых лесов после сплошнолесосечных рубок // Сибирский лесной журн. 2015. № 5. С. 64–76.
Морозова Р.М. Изменение процессов почвообразования под влиянием коцентрированных рубок леса // Возобновление леса на вырубках и выращивание сеянцев в питомниках. 1964а. С. 55–73.
Морозова Р.М., Федорец Н.Г. Современные процессы почвообразования в хвойных лесах Карелии. Петрозаводск: Изд-во Карельского НЦ РАН, 1992. 284 с.
Назарова Л.Е. Климат // Разнообразие биоты Карелии: условия формирования, сообщества, виды. Петрозаводск: Изд-во Карельского НЦ РАН, 2003. С. 6–8.
Орлова М.А., Лукина Н.В., Смирнов В.Э. Методические подходы к отбору образцов лесной подстилки с учетом мозаичности лесных биогеоценозов // Лесоведение. 2015. № 3. С. 214–222.
Решетникова Т.В. Лесные подстилки как депо биогенных элементов // Вестник КрасГАУ. 2011. № 12. С. 74–81.
Рыжова И.М., Подвезенная М.А. Пространственная вариабельность запасов органического углерода в почвах лесных и степных биогеоценозов // Почвоведение. 2008. № 12. С. 1429–1437.
Смирнов А. П., Смирнов А. А., Монгуш Б. А. Естественное лесовозобновление на вырубках Ленинградской области // Известия Санкт-Петербургской лесотехнической академии. 2018. С. 66–83.
Федорец Н.Г. Антропогенная трансформация подзолистых почв после вырубки сосновых древостоев // Антропогенная трансформация таежных экосистем Европы: экологические, ресурсные и хозяйственные аспекты: Матер. междунар. науч.-практ. конф. Петрозаводск: Изд-во Карельского НЦ РАН. 2004. С. 187–191.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Шумаков В.С., Фёдорова Е.Л. Методические рекомендации по определению запасов лесной подстилки и ее зольности при лесоводственных исследованиях. М.: Всесоюз. НИИ лесн. хоз-ва, 1979. 38 с.
Bonan G.B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests // Science. 2008. V. 320. № 5882. P. 1444–1449.
Clarke N., Gundersen P., Jönsson-Belyazid U., Kjønaas O.J., Persson T., Sigurdsson B.D., Stupak I., Vesterdal L. Influence of different tree-harvesting intensities on forest soil carbon stocks in boreal and northern temperate forest ecosystems // Forest Ecology & Management. 2015. V. 351. P. 9–19.
Falsone G., Celi L., Caimi A., Simonov G., Bonifacio E. The effect of clear cutting on podzolisation and soil carbon dynamics in boreal forests (Middle Taiga zone, Russia) // Geoderma. 2012. № 177–178. P. 27–38.
Jandl R., Lindner M., Vesterdal L., Bauwens B., Baritz R., Hagedorn F., Johnson D.W., Minkkinen K., Byrne K.A. How strongly can forest management influence soil carbon sequestration? // Geoderma. 2007. № 137. P. 253–268.
Kauppi P.E. New, low estimate for carbon stock in global forest vegetation based on inventory data // Silva Fennica. 2003. V. 37. № 4. P. 451–457.
Lal R. Forest soils and carbon sequestration // Forest Ecology & Management. 2005. V. 220. № 1–3. P. 242–258.
Nave L.E., Vance E.D., Swanston C.W., Curtis P.S. Harvest impact on soil carbon storage in temperate forest // Forest Ecology & Management. 2010. V. 259. P. 857–866.
Palviainen M., Finér L., Kurka A.-M., Mannerkoski H., Piirainen S., Starr M. Decomposition and nutrient release from logging residues after clear-cutting of mixed boreal forest // Plant Soil. 2004. V. 263. № 1. P. 53–67.
Pan Y., Birdsey R.A., Fang J., Houghton R., Kauppi P.E., Kurz W.A. A large and persistent carbon sink in the world’s forests // Science. 2011. V. 333. № 6045. P. 988–993.
Piirainen, S., Finer L., Starr M. Changes in forest floor and mineral soil carbon and nitrogen stocks in a boreal forest after clear-cutting and mechanical site preparation // European J. Soil Science 2015. V. 66. № 4. P. 735–743.
Ponge J.F. Humus forms in terrestrial ecosystems: framework to biodiversity // Soil Biology & Biochemistry. 2003. V. 35. № 7. P. 935–945.
Scharlemann J.P., Tanner E.V., Hiederer R., Kapos V. Global soil carbon: understanding and managing the largest terrestrial carbon pool // Carbon Managment. 2014. V. 5. № 1. P. 81–91.
Smolander A., Heiskanen J. Soil N and C transformations in two forest clear-cuts during three years after mounding and inverting // Canadian J. Soil Science. 2007. V. 87. P. 251–258.
Smyth C.E., Stinson G., Neilson E., Lempriere T.C., Hafer M., Rampley G.J., Kurz W.A. Quantifying the biophysical climate change mitigation potential of Canada’s forest sector // Biogeosciences. 2014. V. 11. P. 3515–3529.
Дополнительные материалы отсутствуют.