

Лесоведение, 2021, № 3, стр. 265-277
Особенности пыльцы внутривидовых форм лиственницы сибирской в контрастных экотопах Южной Сибири
Т. С. Седельникова a, *, А. С. Аверьянов a, А. В. Пименов a
a Институт леса им. В.Н. Сукачева СО РАН, Федеральный исследовательский центр
“Красноярский научный центр СО РАН”
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: tss@ksc.krasn.ru
Поступила в редакцию 29.01.2020
После доработки 30.03.2020
Принята к публикации 03.02.2021
Аннотация
В естественных популяциях и искусственных насаждениях лиственницы сибирской (Larix sibirica), произрастающих в экологически контрастных условиях Южной Сибири, проведено исследование морфологических признаков пыльцы и аномалий ее развития у внутривидовых форм и морфотипов деревьев, дифференцированных по окраске и размерам женских шишек, с учетом типа половой конституции, возраста, наличия поврежденности лиственничной почковой галлицей, по сравнению с типичными особями. Выявлено, что у красношишечных форм деревьев (f. rubriflora, f. rosea) формируются достоверно более крупные пыльцевые зерна и образуется меньшее количество тератоморфной пыльцы, чем у зеленошишечной формы (f. viridiflora). Специфика крупношишечных форм деревьев по сравнению с мелкошишечными проявляется в продуцировании достоверно более крупной пыльцы. У крупношишечных и мелкошишечных форм деревьев пыльцевые зерна с аномалиями развития встречаются чаще, чем у типичных особей. Молодые 30-летние деревья характеризуются достоверно более мелкой пыльцой, чем особи старших возрастов, не отличаясь от них по количеству тератоморфных пыльцевых зерен. Образование более мелких пыльцевых зерен у молодых деревьев может быть связано не только с возрастом, но и с мужским типом их сексуализации. Заселение деревьев лиственничной почковой галлицей не оказывает влияния на размер пыльцевых зерен и число аномалий их развития, но влияет на изменение соотношения классов формы пыльцы. Морфометрические показатели пыльцы изменяются в соответствии с контрастными различиями эколого-климатических условий произрастания деревьев в подтаежном, предгорном, болотном и степном происхождениях. В условиях неблагоприятного температурного режима корнеобитаемого слоя торфяных криоземов на болоте и засушливого климата со значительными колебаниями температур в степи деревья продуцируют пыльцу более мелких размеров с высоким числом аномалий ее развития.
Лиственница сибирская (Larix sibirica Ledeb.) – вид хвойных растений из рода лиственница (Larix Mill.) семейства сосновые (Pinaceae (Spreng. Ex F. Rudolphi) – имеет обширный ареал, простирающийся с севера на юг от тундры (71° с.ш.) до горных районов Алтая и Саян (46° с.ш.) и с запада на восток – от Урала (62° в.д.) до Забайкалья (123° в.д.). В пределах области своего распространения лиственница сибирская образует широкий спектр морфологической изменчивости, проявляющийся в наличии внутривидовых форм и морфотипов деревьев, адаптированных к разнообразным условиям произрастания (Биоразнообразие …, 2010).
Половая репродукция L. sibirica тесно связана с процессом микроспорогенеза, который определяет формирование и развитие мужского гаметофита. Вопросам пыльцевой продуктивности и фенологии пыления, цитоэмбриологии, биохимии, физиологии и морфологии пыльцевого зерна у лиственницы сибирской и близкородственных видов – лиственницы европейской (Larix decidua Mill.), лиственницы Сукачева (Larix sukaczewii Dylis) и гибридного комплекса – лиственницы Чекановского (L. sibirica × L. gmelinii = = Larix czekanowskii Szaf.) посвящен ряд работ (Круклис, 1974; Круклис, Милютин, 1977; Некрасова, 1983; Тренин, 1986; Slobodnik, 2002; Романова, Третьякова, 2005; Сурсо и др., 2012; Vasilevskaya, Domakhina, 2018). Однако сведения об особенностях морфологии пыльцы у различных форм деревьев, являющихся собственно компонентами биоразнообразия природных популяций и искусственных насаждений лиственницы сибирской, отсутствуют. Между тем, данные о размерах и форме пыльцевых зерен успешно используются для диагностики и экологической дифференциации видов, межвидовых гибридов и внутривидовых форм древесных, в том числе хвойных, растений из родов береза (Betula L.), дуб (Quercus L.), ель (Picea A. Dietr.), кипарис (Cupressus L.), сосна (Pinus L.) (Захаренко, 2006; Пименов и др., 2011; Горячкина, Седаева, 2012; Wrońska-Pilarek et al., 2016; Темботова и др., 2017; Depciuch et al., 2018).
Известно, что морфологические параметры пыльцы как интегральные критерии обладают высокой степенью информативности. Вариабельность размера пыльцы определяется влиянием естественного отбора в зависимости от стратегии ее распространения, в совокупности со многими другими факторами (Knight et al., 2010). У видов хвойных из родов ель, кедр (Cedrus Trew), кипарис, сосна размер пыльцы связан с некоторыми биологическими признаками – строением оболочки пыльцевого зерна, плоидностью клеток, уровнем гетерозиготности генотипа, размером генома (Chira, 1973; Danti et al., 2010; Макогон, Коршиков, 2012; Bell et al., 2018). Для хвойных, в том числе лиственницы сибирской, установлена связь размера пыльцы и встречаемости аномалий ее развития с температурными и почвенно-гидрологическими условиями местопроизрастаний, атмосферным загрязнением, заболеваниями деревьев (Некрасова, 1983; Седельникова и др., 2004; Третьякова, Носкова, 2004; Бажина, 2005; Романова, Третьякова, 2005; Varis et al., 2011; Сурсо и др., 2012; Vasilevskaya, Domakhina, 2018; Sedelnikova et al., 2018).
Настоящая работа посвящена исследованию морфологических признаков пыльцы и выявлению факторов их изменчивости у внутривидовых форм и морфотипов лиственницы сибирской в экологически контрастных условиях произрастания юга Сибири.
ОБЪЕКТЫ И МЕТОДИКА
Объектами исследования послужили естественные популяции и искусственные насаждения L. sibirica, произрастающие на ключевых участках (происхождениях) в Ширинском районе Республики Хакасия и районе Академгородка г. Красноярска (табл. 1). Сбор образцов пыльцы производился в 2017 г. в третьей декаде апреля в искусственных посадках L. sibirica, представляющих собой дендроценоз на урбанизированной территории в городской черте г. Красноярска, в происхождении “Академгородок”, территориально приуроченном к биоклиматическим условиям подтаежного пояса Приенисейской части Восточного Саяна. В первой – второй декадах мая 2017 г. пыльца данного вида отбиралась в естественных популяциях Июсско-Ширинской горностепной ландшафтной системы в 4 происхождениях: “Туим” – редкостойный лиственничник кустарниково-разнотравный на остепненных склонах юго-западной экспозиции; “Марчелгаш” – редкостойный лиственничник злаково-разнотравный на предгорной равнине восточного макросклона Кузнецкого Алатау; “Тунгужуль/суходол” – редкостойный лиственничник кустарниково-разнотравный на первой надпойменной террасе р. Тунгужуль в Бело-Июсской системе межгорных долин восточного макросклона Кузнецкого Алатау; “Тунгужуль/болото” – елово-лиственничное редколесье разнотравно-кустарничково-зеленомошное на криогенном торфянике в пойме р. Тунгужуль в Бело-Июсской системе межгорных долин восточного макросклона Кузнецкого Алатау. Данные ключевые участки (происхождения) отобраны в соответствии с их эколого-климатической контрастностью и подразделяются на подтаежный (“Академгородок”), степной (“Туим”), предгорные (“Тунгужуль/суходол”, “Марчелгаш”) и предгорный болотный (“Тунгужуль/болото”).
Таблица 1.
Характеристика объектов исследования и выборок деревьев
| Наименование ключевого участка/“происхождения” | Географические координаты | Выборки Larix sibirica | |
|---|---|---|---|
| внутривидовые формы и морфотипы | число деревьев, шт./ возраст деревьев, лет | ||
| Микрорайон Академгородок, г. Красноярск/“Академгородок” |
55°59′04″ с.ш., 92°45′09″ в.д. |
Зеленошишечная | 3/50 |
| Красношишечная | 5/50 | ||
| Розовошишечная | 1/50 | ||
| Окрестности с. Туим, Ширинский район, Республика Хакасия/“Туим” | 54°24′03″ с.ш., 89°58′15″ в.д. |
Типичная | 2/125 |
| Мелкошишечная, молодое дерево | 1/30 | ||
| Крупношишечная, мужской тип сексуализации | 1/95 | ||
| Фиолетовошишечная | 1/110 | ||
| Окрестности пос. Марчелгаш, Ширинский район, Республика Хакасия/“Марчелгаш” | 54°30′52″ с.ш., 89°47′02″ в.д. |
Типичная | 2/120 |
| Крупношишечная | 2/ 30 | ||
| Мужской тип сексуализации, молодые деревья | 5/30 | ||
| Первая надпойменная терраса в долине р. Тунгужуль, Ширинский район, Республика Хакасия/“Тунгужуль/суходол” | 54°19′48″ с.ш., 89°43′37″ в.д. |
Типичная | 6/80–90 |
| Мелкошишечная | 1/100 | ||
| Крупношишечная | 1/105 | ||
| Пораженная галлицей | 1/90 | ||
| Криогенный торфяник в долине р. Тунгужуль, Ширинский район, Республика Хакасия/“Тунгужуль/болото” | 54°16′30″ с.ш., 89°38′56″ в.д. |
Типичная | 10/90–110 |
При сборе образцов пыльцы L. sibirica были сформированы следующие выборки: деревья типичной формы; внутривидовые формы деревьев, дифференцированные по окраске кроющих чешуй женских шишек (макростробилов) – зеленошишечная (f. viridiflora Szaf.), красношишечная (f. rubriflora Szaf.) и ее вариации – розовошишечная (f. rosea Szaf.), а также обнаруженная нами уникальная фиолетовошишечная (f. purpura); формы, выделенные по размерам макростробилов – мелкошишечная, крупношишечная. В перечисленных выборках при выделении форм деревьев учитывались их возраст; тип сексуализации; наличие повреждений, вызванных лиственничной почковой галлицей (Dasyneura rozkovi Mam. et Nik. (Diptera, Cecidomyiidae)).
Отбор образцов пыльцы проводили в сухую солнечную погоду из естественно раскрывшихся микростробилов путем стряхивания в пакетики из кальки с последующим помещением их в эксикатор с CaCl2. Из каждой выборки пыльцы было сделано по 3 временных неокрашенных водных препарата, из популяционной выборки – 10 препаратов, в которых было исследовано по 10 полей зрения (n). Морфометрические параметры пыльцевых зерен измерялись при помощи окуляр-микрометра стереоскопического микроскопа МБС-10. Число измеренных в каждой выборке пыльцевых зерен приведено в табл. 2. Измерение параметров элементов пыльцы и определение размера и формы пыльцевого зерна проводились по общепринятым методам (Erdtman, 1945, 1952; Моносзон-Смолина, 1949; Halbritter et al., 2018).
Таблица 2.
Морфометрическая характеристика пыльцевых зерен в изученных происхождениях Larix sibirica
| Внутривидовая форма и морфотип деревьев | Статистические показатели | |||||
|---|---|---|---|---|---|---|
| параметр пыльцевого зерна | n, шт. | Limit, µm | х ± Мx, µm | σ | C.V.,% | |
| “Академгородок” | ||||||
| Зеленошишечная | Высота | 120 | 57–107 | 73.83 ± 0.86 | 9.42 | 12.76 |
| Длина | 120 | 64–114 | 85.72 ± 0.73 | 8.01 | 9.34 | |
| Красношишечная | Высота | 120 | 57–100 | 78.33 ± 0.99 | 10.86 | 13.86 |
| Длина | 120 | 71–107 | 88.17 ± 0.86 | 9.44 | 10.71 | |
| Розовошишечная | Высота | 120 | 57–100 | 78.70 ± 0.99 | 10.86 | 13.80 |
| Длина | 120 | 57–114 | 88.89 ± 0.89 | 9.73 | 10.93 | |
| “Туим” | ||||||
| Типичная | Высота | 120 | 57–94 | 73.60 ± 0.77 | 8.43 | 11.45 |
| Длина | 120 | 57–100 | 85.17 ± 0.69 | 7.51 | 8.82 | |
| Мелкошишечная, молодое дерево | Высота | 120 | 57–100 | 70.37 ± 0.97 | 10.64 | 15.12 |
| Длина | 120 | 64–114 | 85.35 ± 0.92 | 10.13 | 11.87 | |
| Крупношишечная, мужской тип сексуализации | Высота | 120 | 57–100 | 71.38 ± 0.88 | 9.63 | 13.49 |
| Длина | 120 | 57–107 | 85.72 ± 0.83 | 9.05 | 10.56 | |
| Фиолетовошишечная | Высота | 120 | 57–100 | 77.22 ± 0.77 | 8.41 | 10.89 |
| Длина | 120 | 71–100 | 89.37 ± 0.54 | 5.95 | 6.66 | |
| “Марчелгаш” | ||||||
| Типичная | Высота | 240 | 57–100 | 75.46 ± 0.66 | 10.29 | 14.87 |
| Длина | 240 | 71–114 | 87.28 ± 0.53 | 8.19 | 13.56 | |
| Крупношишечная | Высота | 240 | 57–114 | 77.32 ± 0.74 | 11.5 | 10.20 |
| Длина | 240 | 64–121 | 89.93 ± 0.79 | 12.19 | 6.98 | |
| Мужской тип сексуализации, молодые деревья | Высота | 120 | 57–94 | 71.23 ± 0.66 | 7.27 | 10.20 |
| Длина | 120 | 71–100 | 84.17 ± 0.54 | 5.88 | 6.98 | |
| “Тунгужуль/суходол” | ||||||
| Типичная | Высота | 600 | 57–114 | 74.11 ± 0.39 | 9.46 | 12.77 |
| Длина | 600 | 64–114 | 85.82 ± 0.36 | 8.7 | 10.14 | |
| Мелкошишечная | Высота | 240 | 57–100 | 71.95 ± 0.57 | 8.88 | 12.34 |
| Длина | 240 | 71–114 | 83.13 ± 0.52 | 8.08 | 9.72 | |
| Крупношишечная | Высота | 240 | 57–100 | 73.57 ± 0.56 | 8.66 | 11.77 |
| Длина | 240 | 71–107 | 86.02 ± 0.48 | 7.43 | 8.63 | |
| Пораженная галлицей | Высота | 120 | 57–94 | 74.60 ± 0.86 | 9.44 | 12.65 |
| Длина | 120 | 71–100 | 84.39 ± 0.78 | 8.56 | 10.14 | |
| “Тунгужуль/болото” | ||||||
| Типичная | Высота | 120 | 57–100 | 70.88 ± 0.64 | 7.03 | 9.92 |
| Длина | 120 | 71–100 | 83.53 ± 0.56 | 6.10 | 7.30 | |
При измерении пыльцевого зерна наиболее широкая его часть идентифицировалась как длина, наиболее узкая – как высота. Для определения класса пыльцы по размеру использовался ее наибольший морфометрический параметр – длина (Erdtman, 1945). По классификации G. Erdtman (1945), в соответствии со значением длины диагностируются 6 классов размера пыльцевых зерен: <10 µm – очень мелкие (“very small grains”), 10–25 µm – мелкие (“small grains”), 25–50 µm – средние (“medium sized grains”), 50–100 – крупные (“large grains”), 100–200 µm – очень крупные (“very large grains”), >200 µm – гигантские (“gigantic grains”). Классы формы пыльцы определялись на основе соотношения высоты и длины, умноженного на 100 (Erdtman, 1952). По этому соотношению выделяются 9 классов формы пыльцы: <50 – сверх-сплюснутая (“per-oblate”), 50–75 – сплюснутая (“oblate”), 75–88 – почти-сплюснутая (“sub-oblate”), 88–99 – сплюснуто-сфероидальная (“oblate-spheroidal”), 100 – сфероидальная (“spheroidal”), 101–114 – вытянуто-сфероидальная (“prolate-spheroidal”), 114–133 – почти-вытянутая (“sub-prolate”), 133–200 – вытянутая (“prolate”), >200 – сверх-вытянутая (“per-prolate”) (Erdtman, 1952). Подсчет встречаемости и определение типов аномалий пыльцы проводились с использованием методики М.В. Сурсо с соавт. (2012).
Морфометрические параметры пыльцы анализировались с использованием общепринятых статистических методов. Для оценки достоверности различий размера пыльцевых зерен применялся t-критерий Стьюдента (tst) на 5%-м уровне точности по каждой из сравниваемых пар значений (tф). Для использованных в работе объемов выборок по всем сравниваемым парам критическое значение tst составляет 1.96.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Пыльца всех видов лиственницы является сложным объектом, поскольку имеет ограниченное число таксономически значимых признаков и высокий уровень индивидуальной изменчивости. Пыльца лиственницы на искусственных средах и при разных условиях инкубирования in vitro не прорастает (Сурсо и др., 2012), поэтому оценить ее качество можно только косвенными методами. Пыльцевые зерна видов рода Larix имеют следующие морфологические характеристики: единица рассеивания – монада, размер – крупные (51–100 мкм), класс – безапертурный, число апертур – 0, тип апертуры – апертура отсутствует, полярность – гетерополярная, контур в полярном положении (пыльца в гидратированном состоянии) – эллиптический, контур в полярном положении (сухая пыльца) – неправильный, форма (сухая пыльца) – неправильная, складки (сухая пыльца) – неравномерно сформированные, форма (пыльца в гидратированном состоянии) – от яйцевидной до сфероидальной (Halbritter, 2016).
Пыльца лиственницы не имеет воздушных мешков, спородерма состоит из зернистой эктэкзины, ламеллярной эндэкзины и интины. Исходным типом апертуры – части поверхности пыльцевого зерна, являющейся местом выхода пыльцевой трубки – у хвойных является дистальная борозда, которая в процессе эволюции сильно редуцировалась, а у видов рода Larix полностью исчезла. Сфероидальная форма пыльцевого зерна, сглаживание рисунка поверхности экзины, редукция апертур являются признаками специализации под действием анемофилии (Мейер-Меликян, Токарев, 2004; Токарев, 2004). Относительно тяжелая пыльца L. sibirica обладает невысокими аэродинамическими качествами, однако достаточно низкая удельная плотность обеспечивает ее способность к переносу на большие расстояния (Сурсо, 2009; Головко и др., 2011).
Сведения о размере пыльцы лиственницы сибирской крайне немногочисленны и приводятся для популяционных выборок деревьев или отдельных особей без учета особенностей их габитуса и морфологии. В связи с представлениями о том, что пыльца L. sibirica имеет близкую к сфероидальной форму, при исследованиях определялись значения только одного показателя – длины пыльцевого зерна. По имеющимся данным, длина пыльцевого зерна L. sibirica в северной подзоне тайги (Архангельская область) составляет 83–86 µm, в Карелии и Мурманской области – 77–79 µm, в районах Восточной Сибири – 76–87 µm, в Южной Сибири (Республика Хакасия) –78–79 µm. Коэффициент вариации (C.V.) длины пыльцевого зерна лиственницы сибирской определяется как низкий – 4–8% (Круклис, Милютин, 1977; Тренин, 1986; Баранчиков и др., 2009; Сурсо и др., 2012).
На рисунке 1 приведены микрофотографии зрелых пыльцевых зерен лиственницы сибирской в образцах пыльцы, использованных нами для анализа. Для более объективной оценки изменчивости размера пыльцы L. sibirica измерялись параметры длины и высоты пыльцевого зерна. По данным, приведенным в табл. 2, в образцах пыльцы L. sibirica средние параметры длины пыльцевых зерен варьируют от 83.1 ± 0.52 до 89.9 ± 0.79 µm, высоты – от 70.4 ± 0.97 до 78.7 ± 0.99 µm. В образцах пыльцы из всех происхождений высота пыльцевых зерен оказалась более вариабельным параметром по сравнению с их длиной, при этом значения коэффициентов вариации соответствуют низкому и среднему уровням изменчивости (табл. 2). По классификации G. Erdtman (1945), в образцах L. sibirica в соответствии со значениями длины идентифицируется пыльца двух классов размера: крупные пыльцевые зерна (“large grains”), длина которых варьирует в диапазоне 50–100 µm, и очень крупные пыльцевые зерна (“very large grains”), длина которых составляет 100–200 µm. Во всех образцах лиственницы преобладает крупная пыльца, однако в некоторых из них встречаются очень крупные пыльцевые зерна (табл. 3).
Рис. 1.
Зрелые пыльцевые зерна L. sibirica: a – происхождение “Академгородок”, зеленошишечная форма; б – происхождение “Туим”, типичная форма; в – происхождение “Тунгужуль/суходол”, крупношишечная форма. Стрелками указаны проталлиальные клетки.
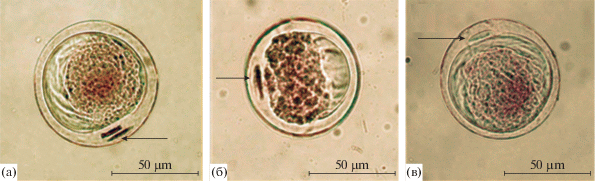
Таблица 3.
Встречаемость пыльцы различного класса размера в изученных происхождениях Larix sibirica, %
| Внутривидовая форма и морфотип деревьев | Класс размера пыльцевых зерен | |
|---|---|---|
| крупные | очень крупные | |
| “Академгородок” | ||
| Зеленошишечная | 98.3 | 1.7 |
| Красношишечная | 95.0 | 5.0 |
| Розовошишечная | 94.2 | 5.8 |
| “Туим” | ||
| Типичная | 100.0 | 0.0 |
| Мелкошишечная, молодое дерево | 97.5 | 2.5 |
| Крупношишечная, мужской тип сексуализации | 95.0 | 5.0 |
| Фиолетовошишечная | 100.0 | 0.0 |
| “Марчелгаш” | ||
| Типичная | 99.6 | 0.4 |
| Крупношишечная | 87.5 | 12.5 |
| Мужской тип сексуализации, молодые деревья | 100.0 | 0.0 |
| “Тунгужуль/суходол” | ||
| Типичная | 98.2 | 1.8 |
| Мелкошишечная | 99.6 | 0.4 |
| Крупношишечная | 99.6 | 0.4 |
| Пораженная галлицей | 100.0 | 0.0 |
| “Тунгужуль/болото” | ||
| Типичная | 100.0 | 0.0 |
Примечание. Число анализированных пыльцевых зерен (n) по каждой из выборок деревьев представлено в табл. 2.
Пыльца красношишечных форм деревьев (f. rubriflora, f. rosea, f. purpura) из происхождений “Академгородок” и “Туим” достоверно крупнее по сравнению с зеленошишечной формой (f. viridiflora) по параметрам длины (tф = 3.07, 3.90, 5.67, соответственно) и высоты (tф = 4.85, 5.25, 4.16, соответственно) (табл. 2). Возможно, это обусловливается наличием большого количества пыльцы очень крупного размера, длина которой варьирует от 100 до 114 µm у форм f. rubriflora и f. rosea (5.0 и 5.8% соответственно), хотя у формы f. purpura имеется только крупная пыльца и отсутствует очень крупная. Для отдельных видов древесных растений, например, из рода эвкалипт (Eucalyptus L’Hér), показано, что более крупные пыльцевые зерна имеют преимущественные шансы на успех в процессе оплодотворения, так как их размер определяет скорость роста пыльцевых трубок (Gore et al., 1990). Поскольку в сибирских популяциях L. sibirica зеленошишечные особи встречаются редко, а преобладают красношишечные деревья, встречаемость которых по мере продвижения на северо-восток в более холодные районы возрастает до 90% и выше (Круклис, Милютин, 1977; Биоразнообразие …, 2010), можно предположить, что одной из вероятных причин их устойчивости в суровых условиях произрастания является продуцирование более крупной пыльцы.
Хвойные деревья с красной окраской кроющих чешуй женских и мужских репродуктивных органов содержат антоцианы, активация биосинтеза которых происходит в стрессовых условиях произрастания. У представителей семейства Pinaceae выявлено 4 антоцианидина, один из которых (3-глюкозид пеонидин) обнаружен в женских шишках лиственницы (Anderson, 1992; Griesbach, Santamour, 2003). Показано, что формы деревьев, содержащие антоциан в мужских и женских шишках, более часто встречаются в условиях низких температур. Так, пыльца краснопыльниковой формы сосны обыкновенной (Pinus sylvestris L. f. (var.) erythrantera Sanio) при низких температурах лучше прорастает, характеризуется более крупными и менее изменчивыми размерами, а ее вылет происходит раньше по сравнению с желтопыльниковой формой (f. (var.) sulfuranthera Kozubov) (Некрасова, 1983; Пименов и др., 2011; Темботова и др., 2017). У красношишечных форм лиственницы сибирской выход семян из шишек выше, чем у зеленошишечных, что может свидетельствовать о более успешных процессах опыления у таких деревьев (Ковылина и др., 2008). Дифференциация форм деревьев по окраске шишек, связанная с параметрами пыльцы, вероятно, обеспечивает эффективность репродукции хвойных в широком диапазоне температурных условий.
По данным табл. 2, среди проанализированных образцов наиболее мелкие пыльцевые зерна сформировались у деревьев L. sibirica из происхождения “Тунгужуль/болото”, наиболее контрастного по условиям температурного режима почвы по отношению к остальным происхождениям. Различия между типичными деревьями из происхождения “Тунгужуль/болото” и тремя другими участками – “Тунгужуль/суходол”, “Туим” и “Марчелгаш” – достоверны как по длине пыльцы (tф = 2.64–4.15), так и по ее высоте (tф = = 3.55–4.47). Примечательно, что в выборке “Тунгужуль/болото” отсутствует пыльца очень крупного размера (табл. 3). Вероятно, уменьшение размера пыльцы у L. sibirica из происхождения “Тунгужуль/болото” связано с произрастанием деревьев в условиях экстремальной экологической обстановки при низкой температуре корнеобитаемого слоя торфяных криоземов.
Пыльцевые зерна L. sibirica крупношишечной формы из происхождений “Тунгужуль/суходол” и “Марчелгаш” достоверно крупнее по сравнению с мелкошишечной формой из происхождений “Туим” и “Тунгужуль/суходол” по высоте (tф = 2.84–8.14) и длине (tф = 3.55–10.15). Различия недостоверны (tф = 0.71) лишь по высоте пыльцы крупно- и мелкошишечной форм из происхождения “Тунгужуль/суходол” (табл. 2). Выявленные различия могут определяться большим количеством пыльцевых зерен очень крупного размера в рассматриваемых образцах пыльцы крупношишечных деревьев, но, вероятно, не только этим фактором. Так, по данным табл. 3, доля очень крупной пыльцы у трех крупношишечных деревьев из происхождений “Туим” и “Марчелгаш” довольно высокая (5.0 и 12.5% соответственно). В то же время, у крупношишечного дерева из происхождения “Тунгужуль/суходол” она незначительная (0.4%). У двух мелкошишечных деревьев из происхождений “Туим” и “Тунгужуль/суходол” встречаемость пыльцевых зерен очень крупного размера так же различна (2.5 и 0.4% соответственно) (табл. 3). Пыльца указанных крупношишечных деревьев из происхождений “Туим”, “Тунгужуль/суходол” и “Марчелгаш” несколько крупнее по длине по сравнению не только с мелкошишечными, но и обычными особями из этих происхождений, характеризующимися средними по размерам шишками (табл. 2). По некоторым данным, у крупношишечных форм лиственницы сибирской формируются семена с более высокой массой (Зеленяк и др., 2018).
Необходимо отметить, что в происхождении “Марчелгаш” молодые 30-летние деревья отличаются достоверно более мелкими размерами пыльцевых зерен относительно деревьев старших возрастов, произрастающих на этом участке, как по высоте (tф = 5.30; 4.03, соответственно), так и по длине (tф = 4.89; 3.70, соответственно) (табл. 2). Для хвойных, в частности, P. sylvestris, показано формирование более мелкой пыльцы у деревьев молодого возраста (Некрасова, 1983). Однако возможно, что продуцирование более мелкой пыльцы у молодых деревьев из происхождения “Марчелгаш” связано не только с возрастом этих особей, но и с их половой конституцией, представляющей мужской тип сексуализации. В происхождении “Марчелгаш” деревья мужского типа сексуализации характеризуются наличием многочисленных микростробилов в кроне и продуцированием большого количества пыльцы, что, по всей вероятности, не может обеспечить ее крупные размеры. В то же время дерево мужского типа сексуализации и молодое 30-летнее дерево из происхождения “Туим” достоверно не отличаются по размеру пыльцы от остальных деревьев, произрастающих на этом участке (табл. 2).
Пыльцевые зерна с дерева, пораженного лиственничной почковой галлицей, из образца “Тунгужуль/суходол” не отличаются по размерам от пыльцы остальных шести типичных деревьев из данного пункта сбора и происхождений “Туим” и “Марчелгаш” (табл. 2). Сходные результаты получены Ю.Н. Баранчиковым с соавт. (2009): по данным, приведенным в их работе, у лиственницы в Хакасии заражение деревьев галлицей не оказало влияния на размеры пыльцы и содержание в ней крахмала, жиров и пероксидазы.
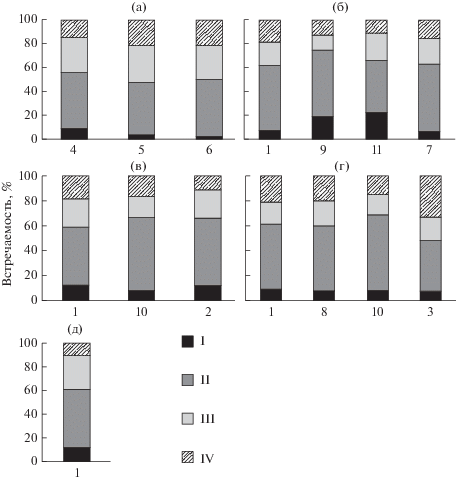
Разнообразие форм пыльцевых зерен в исследованных образцах пыльцы лиственницы сибирской, по классификации G. Erdtman (1952), соответствует 4 классам: сплюснутая (“oblate”), почти-сплюснутая (“sub-oblate”), сплюснуто-сфероидальная (“oblate-spheroidal”), сфероидальная (“spheroidal”). Данные, приведенные на рис. 2, дают представление о встречаемости пыльцевых зерен различных классов формы в конкретных выборках и происхождениях L. sibirica. Оказалось, что у деревьев из всех происхождений преобладает пыльца почти-сплюснутой формы, хотя встречаемость ее неодинакова и варьирует от 40.8 до 62.1%. Дерево, пораженное лиственничной почковой галлицей из происхождения “Тунгужуль/суходол”, отличается минимальной долей пыльцевых зерен почти-сплюснутой формы (40.8%) с одновременным возрастанием доли сфероидальной формы пыльцы (32.5%) в образце.
Рис. 2.
Встречаемость форм пыльцы L. sibirica: Происхождения: а – “Академгородок”, б – “Туим”, в – “Марчелгаш”, г – “Тунгужуль/суходол”, д – “Тунгужуль/болото”. Форма пыльцевых зерен: I – сплюснутая, II – почти сплюснутая, III – сплюснуто-сфероидальная, IV – сфероидальная. Выборки L. sibirica: 1 – обычная форма; 2 – обычная форма, молодые деревья мужского типа сексуализации; 3 – обычная форма, пораженная галлицей; 4 – зеленошишечная; 5 – красношишечная; 6 – розовошишечная; 7 – фиолетовошишечная; 8 – мелкошишечная; 9 – мелкошишечная, молодое дерево; 10 – крупношишечная; 11 – крупношишечная, мужского типа сексуализации.
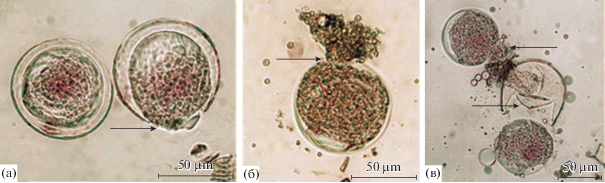
Во всех образцах пыльцы L. sibirica наряду с нормально сформированными пыльцевыми зернами обнаружена пыльца с аномалиями развития (тератоморфная). К категории нормальных относились пыльцевые зерна без видимых нарушений размера и формы (рис. 1). Данные, характеризующие соотношение нормальных и тератоморфных пыльцевых зерен в образцах и встречаемость типов аномалий пыльцы в различных происхождениях лиственницы сибирской, приводятся на рис. 3. Очевидно, что основную массу тератоморфной пыльцы составляют пыльцевые зерна с признаками плазмолиза цитоплазмы. В некоторых случаях наблюдались недоразвитие и деформация пыльцевых зерен, разрывы экзины (рис. 4). Единично отмечены двойные микрогаметофиты и аномально крупные, очевидно, диплоидные, пыльцевые зерна.
Рис. 3.
Встречаемость нормально развитой и тератоморфной пыльцы и спектр аномалий пыльцевых зерен L. sibirica: Происхождения: а – “Академгородок”, б – “Туим”, в – “Марчелгаш”, г – “Тунгужуль/суходол”, д – “Тунгужуль/болото”. Категория пальцевых зерен: I – плазмолиз, II – прочие аномалии, III – нормальные. Выборки L. sibirica: 1 – обычная форма; 2 – обычная форма, молодые деревья мужского типа сексуализации; 3 – обычная форма, пораженная галлицей; 4 – зеленошишечная; 5 – красношишечная; 6 – розовошишечная; 7 – фиолетовошишечная; 8 – мелкошишечная; 9 – мелкошишечная, молодое дерево; 10 – крупношишечная; 11 – крупношишечная, мужского типа сексуализации.
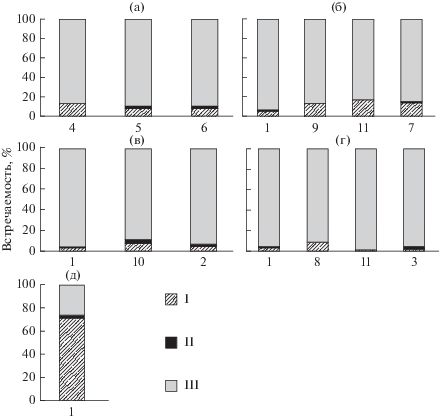
Рис. 4.
Аномалии пыльцевых зерен L. sibirica: а – разрыв экзины (указано стрелкой), происхождение “Марчелгаш”, крупношишечная форма; б – разрыв экзины с вытеканием цитоплазмы (указано стрелкой), происхождение “Тунгужуль/болото”; в – разрыв экзины с вытеканием цитоплазмы (вверху, указано стрелкой) и сбрасывание экзины (внизу, указано стрелкой), происхождение “Тунгужуль/суходол”, фиолетовошишечная форма.
По данным, представленным на рис. 3, в происхождении “Академгородок” красношишечные формы деревьев (f. rubriflora, f. rosea) содержат меньшее количество аномальной пыльцы, составляющее 7.8–8.3%, чем зеленошишечная форма (f. viridiflora), у которой доля тератоморфных пыльцевых зерен достигает 11.8%. Почти у всех особей, отличающихся морфологическим своеобразием женских шишек (за исключением одной крупношишечной формы из происхождения “Тунгужуль/суходол”), – двух мелкошишечных деревьев из происхождений “Туим” и “Тунгужуль/суходол”, фиолетовошишечной формы (f. purpura) из происхождения “Туим”, двух крупношишечных деревьев из происхождения “Марчелгаш” – тератоморфная пыльца встречается с высокой частотой (10.5–16.6%), в то время как у типичных деревьев из этих происхождений – с более низкой (2.2–6.6%). Установлено (Седельникова, Пименов, 2017), что в проростках семян с мелкошишечных и других отклоняющихся от типичной формы деревьев L. sibirica из Ширинско-Июсской степи выявлены нарушения числа хромосом и хромосомные перестройки, что может быть связано с наличием аномалий пыльцы, влияющих на качество семенного потомства, у таких особей. Зависимости между встречаемостью аномалий пыльцевых зерен и возрастом деревьев, их половой конституцией, а также наличием повреждений, вызванных лиственничной почковой галлицей, не выявлено (рис. 3).
Максимальным числом тератоморфных пыльцевых зерен (73.1%) отличаются деревья из происхождения “Тунгужуль/болото”, что, вероятно, связано с экстремальными температурными факторами их произрастания на мерзлотных почвах – торфяных криоземах (рис. 3). Высокая доля аномальной пыльцы (13–16%) отмечается у деревьев из ключевого участка “Туим”, характеризующегося наиболее засушливыми условиями произрастания и резкими перепадами суточных и сезонных температур среди всех исследованных происхождений.
По литературным данным (Тренин, 1986; Slobodník, 2002; Романова, Третьякова, 2005; Калашник, 2011; Сурсо и др., 2012; Vasilevskaya, Domakhina, 2018), у L. sibirica и близкородственных видов L. decidua и L. sukazcewii выявлены сходные типы аномалий пыльцы – пыльцевые зерна без содержимого, с редуцированным содержимым и признаками полной или частичной деградации ядра и цитоплазмы, нарушениями экзины, недоразвитые, мелкие, деформированные, гигантские, двойной микрогаметофит, полиспория. При этом отмечается, что тератологические изменения пыльцевых зерен и стерильность пыльцы L. sibirica могут быть вызваны нарушениями в процессе микроспорогенеза, неблагоприятными экологическими условиями, резкими изменениями погоды, а также техногенными выбросами различной природы.
ВЫВОДЫ
В естественных популяциях и искусственных насаждениях лиственницы сибирской, произрастающих в контрастных экологических условиях Южной Сибири, у внутривидовых форм и морфотипов деревьев, дифференцированных по окраске и размерам макростробилов с учетом их половой конституции, возраста, наличия пораженности лиственничной почковой галлицей, по сравнению с особями типичной формы, выявлен ряд особенностей морфологии пыльцы и аномалий ее развития:
1. У красношишечных форм деревьев L. sibirica (f. rubriflora, f. rosea) формируются достоверно более крупные пыльцевые зерна и образуется меньшее количество тератоморфной пыльцы, чем у зеленошишечной формы (f. viridiflora).
2. Специфика крупношишечных форм деревьев L. sibirica проявляется в продуцировании достоверно более крупной пыльцы по сравнению с особями с мелкими по размеру шишками. У крупношишечных и мелкошишечных форм деревьев пыльцевые зерна с аномалиями развития встречаются чаще, чем у типичных особей.
3. Молодые 30-летние деревья L. sibirica характеризуются достоверно более мелкой пыльцой, чем особи старших возрастов, не отличаясь от них по количеству тератоморфных пыльцевых зерен. Образование более мелких пыльцевых зерен у молодых деревьев может быть связано не только с возрастом, но и с мужским типом их сексуализации, обусловливающей наличие многочисленных микростробилов и продуцирование большого количества пыльцы, что не обеспечивает ее крупные размеры.
4. Заселение L. sibirica лиственничной почковой галлицей не оказывает влияния на размер пыльцевых зерен и число аномалий их развития, однако у пораженного дерева выявлено изменение соотношение классов формы пыльцы.
5. Морфометрические показатели пыльцы L. sibirica изменяются в соответствии с контрастными различиями эколого-климатических факторов подтаежного, предгорного, болотного и степного происхождений. Неблагоприятный температурный режим корнеобитаемого слоя торфяных криоземов в болотном экотопе, по всей вероятности, определяет уменьшение размера пыльцы лиственницы и возрастание числа аномалий ее развития. В степном экотопе условия засушливого климата и значительные колебания сезонных и суточных температур могут обусловливать увеличение количества тератоморфной пыльцы.
Список литературы
Бажина Е.В. Жизнеспособность пыльцы и изменчивость признаков побегов Abies sibirica, пораженных ржавчинным раком (Melampsorella cerastii) // Ботанический журн. 2005. Т. 90. № 5. С. 696–702.
Баранчиков Ю.Н., Третьякова И.Н., Буглова Л.В. Генеративный потенциал деревьев лиственницы сибирской, пораженных почковой галлицей // Вестник Московского государственного университета леса – Лесной вестник. 2009. Т. 68. № 5. С. 134–137.
Биоразнообразие лиственниц Азиатской России / Отв. ред. Ефремов С.П., Милютин Л.И. Новосибирск: Гео, 2010. 159 с.
Головко В.В., Куценогий К.П., Истомин В.Л. Использование конфокальной сканирующей микроскопии для определения объема и плотности пыльцевых зёрен сибирских растений // Труды VII международной выставки и Научного конгресса ГЕО-СИБИРЬ-2011. Дистанционные методы зондирования земли и фотограмметрия, мониторинг окружающей среды, геоэкология. Новосибирск: Изд-во СГГА, 2011. Т. 4. С. 249–253.
Горячкина О.В., Седаева М.И. Морфология и качество пыльцы у видов рода Picea (Pinaceae) из коллекции дендрария Института леса им. В.Н. Сукачева СО РАН // Растительный мир Азиатской России. 2012. Т. 10. № 2. С. 27–32.
Захаренко Г.С. Биологические основы интродукции и культуры видов рода кипарис (Cupressus L.). Киев: Аграрна наука, 2006. 256 с.
Зеленяк А.К., Морозова Е.В., Иозус А.П. Качество семян лиственницы сибирской на клоновой лесосеменной плантации // Успехи современного естествознания. 2018. № 1. С. 13–17.
Калашник Н.А. Аномалии пыльцы у лиственницы Сукачева в различных экологических условиях // Известия Самарского НЦ РАН. 2011. Т. 13. № 1(14). С. 835–838.
Ковылина О.П., Ковылин Н.В., Познахирко П.Ш. Изменчивость генеративных органов и посевные качества семян лиственницы сибирской в защитных насаждениях оз. Шира // Хвойные бореальной зоны. 2008. Т. XXV. № 3–4. С. 309–315.
Круклис М.В. Мейоз и формирование пыльцы у лиственницы Чекановского (Larix Czekanowskii Cz) // Изменчивость древесных растений Сибири. Красноярск: Институт леса и древесины Сибирского отделения Академии наук СССР, 1974. С. 20–34.
Круклис М.В., Милютин Л.И. Лиственница Чекановского. М.: Наука, 1977. 212 с.
Макогон И.В., Коршиков И.И. Качество пыльцы в связи с генетическими особенностями Picea abies (L.) Karst. в интродукционном насаждении // Бюллетень Государственного Никитского ботанического сада. 2012. Вып. 105. С. 107–112.
Мейер-Меликян Н.Р., Токарев П.И. Особенности строения пыльцевых зерен некоторых представителей Pinaceae по данным электронной микроскопии // Бюллетень Московского общества испытателей природы. Отдел биологический. 2004. Т. 109. Вып. 1. С. 19–21.
Моносзон-Смолина М.Х. К вопросу о морфологии пыльцы некоторых видов рода Pinus // Ботанический журнал. 1949. Т. 34. № 4. С. 352–380.
Некрасова Т.П. Пыльца и пыльцевой режим хвойных Сибири. Новосибирск: Наука, 1983. 169 с.
Пименов А.В., Седельникова Т.С., Ефремов С.П. Морфология и качество пыльцы желто- и краснопыльниковой форм Pinus sylvestris в болотных и суходольных условиях произрастания (Томская область) // Ботанический журн. 2011. Т. 96. № 3. С. 367–376.
Романова Л.И., Третьякова И.Н. Особенности микроспорогенеза у лиственницы сибирской, растущей в условиях техногенной нагрузки // Онтогенез. 2005. Т. 36. № 2. С. 128–133.
Седельникова Т.С., Пименов А.В. Числа хромосом форм Larix sibirica (Pinaceae) в Ширинской степи Республики Хакасия // Ботанический журнал. 2017. Т. 102. № 5. С. 693–697.
Седельникова Т.С., Пименов А.В., Ефремов С.П. Морфология пыльцы сосны обыкновенной на болотах и суходолах // Лесоведение. 2004. № 6. С. 1–5.
Сурсо М.В. Пыльцевой режим в хвойных насаждениях Севера // Лесной вестник. 2009. № 3. С. 33–37.
Сурсо М.В., Барабин А.И., Болотов И.Н., Филиппов Б.Ю. Весеннее развитие пыльцы у лиственницы сибирской (Larix sibirica Ledeb.) в северной подзоне тайги // Известия высших учебных заведений. Лесной журнал. 2012. № 6. С. 7–15.
Темботова Ф.А., Моллаева М.З., Пшегусов Р.Х. Изменчивость пыльцы желто- и краснопыльниковой форм сосны обыкновенной (Pinus sylvestris L.) на территории национального парка “Приэльбрусье” (Центральный Кавказ) // Известия Дагестанского государственного педагогического университета. Естественные и точные науки. 2017. Т. 11. С. 55–61.
Токарев П.И. Палинология древесных растений, произрастающих на территории России: Автореф. дис. … докт. биол. наук (03.00.05) М.: МГУ, 2004. 55 с.
Тренин В.В. Цитоэмбриология лиственницы. Л.: Наука, 1986. 88 с.
Третьякова И.Н., Носкова Н.Е. Пыльца сосны обыкновенной в условиях экологического стресса // Экология. 2004. № 1. С. 26–33.
Anderson Ø.M. Anthocyanins from reproductive structures in Pinaceae // Biochemical Systematics & Ecology. 1992. V. 20. P. 145–148.
Bell B.A., Bishop T.H., Fletcher W.J., Ryan P., Ilmen R. Cedrus atlantica pollen morphology and investigation of grain size variability using laser diffraction granulometry // Palynology. 2018. V. 42. N 3. P. 339–353.
Chira E. The development of pollen with more than the haploid chromosome number in Pinus sylvestris // Biologia. 1973. V. 28. № 7. P. 515–527.
Danti R., Della Rocca G., Mori B., Torraca G., Calamassi R., Mariotti Lippi M. Old world and new world Cupressus pollen: morphological and cytological remarks // Plant Systematics & Evolution. 2010. V. 287. № 3. P. 167–177.
Depciuch J., Kasprzyk I., Drzymała E., Parlinska-Wojtan M. Identification of birch pollen species using FTIR spectroscopy // Aerobiologia. 2018. V. 34. Iss. 4. P. 525–538.
Erdtman G. Pollen morphology and plant taxonomy. III. Morina L., with an addition on pollen-morphological terminology // Svensk Botanisk Tidskrift. 1945. V. 39. P. 279–285.
Erdtman G. Pollen morphology and plant taxonomy: Angiosperms. An introduction to palynology. Stockholm: Almqvist & Wiksell, 1952. 539 p.
Gore P.L., Potts B.M., Volker P.W., Megalos J. Unilateral cross incompatibility in Eucalyptus: the case of hybridisation between E. globulus and E. nitens // Australian Journal of Botany. 1990. V. 38. P. 383–394.
Griesbach R., Santamour F.S. Anthocyanins in cones of Abies, Picea, Pinus, Pseudotsuga and Tsuga (Pinaceae) // Biochemical Systematics & Ecology. 2003. V. 31. № 3. P. 261–268.
Halbritter H. Larix decidua // PalDat – A palynological database. 2016. https://www.paldat.org/pub/Larix_decidua/ 302356;jsessionid=0EB4C4A42EB60B4844E5E61032605C90; accessed 2018-12-19.
Halbritter H., Ulrich S., Grimsson F., Weber M., Zetter R., Hesse M., Buchner R., Svojtka M., Frosch-Radivo A. Illustrated pollen terminology. Vienna: Springer, 2018. 483 p.
Knight Ch.A., Clancy R.B., Gotzenberger L., Dann L., Beaulieu J.M. On the relationship between pollen size and genome size // Journal of Botany. 2010. Article ID 612017. P. 1–7.
Sedel’nikova T.S., Pimenov A.V., Efremov S.P., Muratova E.N. Peculiarities of the male generative sphere in Pinus sibirica from forest bog ecotopes of Western Siberia // Biology Bulletin. 2018. V. 45. № 1. P. 23–30.
Slobodník B. The early-spring development of male generative organs and abnormalities in pollen ontogenesis of European larch (Larix decidua Mill.) // Forest Genetics. 2002. V. 9. № 4. P. 309–314.
Varis S., Reiniharju J., Santanen A., Ranta H., Pulkkinen P. The size and germinability of Scots pine pollen in different temperatures in vitro // Grana. 2011. V. 50. № 2. P. 129–135.
Vasilevskaya N.V., Domakhina A.D. Teratomorphism of pollen of Larix sibirica Ledeb. (Pinaceae Lindl.) in the Arctic urbanized territory // Czech Polar Reports. 2018. V. 8. № 1. P. 24–36.
Wrońska-Pilarek D., Danielewicz W., Bocianowski J., Maliński T., Janyszek M. Comparative pollen morphological analysis and its systematic implications on three European oak (Quercus L., Fagaceae) species and their spontaneous hybrids // PLoS One. 2016. V. 11. № 8: e0161762. https://doi.org/10.1371/journal.pone.0161762
Дополнительные материалы отсутствуют.