

Лесоведение, 2022, № 4, стр. 351-363
Пространственно-временная динамика биологической активности почв в фитогенном поле сосны обыкновенной в средней тайге Карелии
Е. В. Мошкина a, *, О. Н. Бахмет b, М. В. Медведева a, А. Ю. Карпечко a, А. В. Мамай a
a Институт леса КарНЦ РАН
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, д. 11, Россия
b Федеральный исследовательский центр “Карельский научный центр Российской академии наук”
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, д. 11, Россия
* E-mail: lena_moshkina@mail.ru
Поступила в редакцию 09.11.2021
После доработки 24.01.2022
Принята к публикации 06.04.2022
- EDN: WBHMHI
- DOI: 10.31857/S0024114822040076
Аннотация
В 2017–2020 гг. в среднетаежной подзоне Карелии в сосняке брусничном исследована биологическая активность подзола иллювиально-железистого песчаного в различных зонах фитогенного поля дерева (ФПД) и факторов, на нее влияющих. В ФПД выделяли три функциональные зоны: “ствол”, “крона”, “окно”. Запасы лесной подстилки (ЗЛП) в приствольной зоне и под кроной деревьев варьировали от 27.69 до 78.34 т/га, в межкроновом пространстве в среднем составляли 27.69 т/га. Варьирование ЗЛП, обусловленное влиянием ФПД, составляло 15–37%. Годовое поступление опада было максимальным в подкроновом пространстве (2.47–2.49 т/га), а минимальным – в “окне” (2.37 т/га). Время оборачиваемости лесной подстилки составило 15–16 лет. Установлено, что крона нивелирует суточный и сезонный температурный режим почвы, снижая максимальные и увеличивая минимальные значения температуры верхнего слоя почвы. Сезонная динамика эмиссии СО2 из почвы на 41–56% определялась температурой почвы на глубине 10 см. Относительная влажность почвы (Wп) была максимальной в осенне-весенний период и составляла на глубине 0–5 см 65–77%, на глубине 5–15 см – 20–24%. В летние месяцы Wп снижалась до 50–65% на глубине 0–5 см, до 10–15% на – 5–15 см. В зависимости от сочетания погодных условий и сезонной динамики потоков СО2 из почвы в сосняке брусничном годовые эмиссии в разных зонах ФПД отличались в 1.5 раза и составляли в окне – 5.6 ± 0.7, у ствола – 8.5 ± 0.9, под кроной – 7.7 ± 0.9 т С/га в год. Вклад дыхания корней в общее дыхание почв составил 24–45%, при этом условия зон ФПД значимо не влияли на плотность размещения микориз на корнях сосны обыкновенной (Pinus sylvestris). Тенденция более высоких показателей микробиологической активности почв прослеживается в приствольной и подкроновой зоне дерева. Наибольшую численность имеют микроорганизмы, использующие органические формы азота. Численность комплекса целлюлозоразрушающих микроорганизмов низкая, что определяет заторможенность процессов деструкции органического вещества и формирование лесных подстилок. Основным экологическим фактором, влияющим на скорость деструкционных процессов и величину почвенных потоков СО2 в сосняке брусничном, является зона ФПД. Сезонная динамика почвенной эмиссии СО2 детерминируется температурой воздуха и почвы. В результате кластерного анализа, проведенного на основе изучаемых свойств, были получены два кластера. Один из них включал функциональную зону “окно”, другой – “ствол” и “крона”.
Древесные растения, являясь эдификаторами лесных экосистем, напрямую и опосредованно воздействуют на различные компоненты биогеоценоза, в том числе определяют свойства почвы, включая ее микробиологические характеристики (Gersper, Holowaychuk, 1970; Boettcher, Kalisz, 1990; Liski, 1995). Кроме того, колебательные движения деревьев создают вихревые потоки, которые являются одним из условий формирования фитогенного поля. В пределах фитогенного поля можно выделить три функциональные зоны: “ствол”–приствольное повышение, “крона”–подкроновое пространство, “окно”–межкроновое пространство, которые хорошо индицируются по мощности лесной подстилки и количеству еe запасов, а также по показателям кислотности, плотности почв, содержания гумуса в верхней части почвенного профиля. Микробиологические показатели также могут отражать условия функционирования различных зон фитогенного поля дерева. Кроны деревьев по-разному влияют на распределение атмосферных осадков, а также поступающих из выпадений элементов минерального питания и загрязнителей на территории леса, что также сказывается на свойствах почв (Карпачевский и др., 1998; Арчегова, Кузнецова, 2011, Карпечко и др., 2015; Карпечко и др., 2018). От приствольных возвышений к межкроновым пространствам увеличивается разнообразие растений живого напочвенного покрова, возрастает общее число видов и доля участия неморальных, светолюбивых, а также требовательных к плодородию почв видов растений (Семенюк и др., 2020). Растительный опад и отпад также существенно влияют на физико-химические и микробиологические свойства почвы, определяя их кислотность, запасы подстилки, толщину гумусового слоя (Hokkanen et al., 1995, Lawrence et al., 2012). Количество и качество растительного опада в различных зонах фитогенного поля дерева изменяется в широких пределах, его трансформация зависит от вида древесной растительности и растений напочвенного покрова. Таким образом, мозаичность почвенных условий во многом связана с формированием фитогенного поля дерева (Карпачевский, Просвирина, 1989), термином, введенным А.А. Урановым (Уранов, 1965). Значительное количество работ посвящено изучению свойств почв в ФПД, при этом наибольшее внимание заслуживает цикл работ В.В. Никонова и Н.В. Лукиной, которые были частью комплексных исследований пространственного варьирования свойств почв естественных и антропогенно нарушенных лесных экосистем Северной Фенноскандии (Лукина и др., 2003; Никонов, Лукина, 2000). Для почв Карелии первые исследования по данной проблеме проведены Л.М. Загуральской (Загуральская, 2003). Было показано, что изменения микробиологических свойств почв зависят от живого напочвенного покрова и расстояния от дерева (Медведева и др., 2020). Особенности трансформации биофильных элементов в почве различных зон ФПД во многом определяются функционированием микроорганизмов, которые обладают широкими возможностями метаболизма. Помимо надземной части древесных растений фитогенное поле дерева формирует его подземная часть. Корневая система, корневые выделения древесных растений могут контролировать большую часть окружающей их среды – регулировать состав почвенной микробиоты, притягивать и концентрировать питательные вещества, менять химические и физические характеристики участка почвы, на котором они растут (Калинин, 1991). Для всех лесообразующих пород в бореальных лесах характерно наличие эктомикоризы (Шубин, 1973; Taylor et al., 2000), таким образом, большая часть элементов минерального питания, поступающая в растение из почвы, проходит через интерфейс “гриб-корень” (Ostonen et al., 2011).
Сложная комплементарность и динамичность отдельных биотических блоков в функционировании фитогенного поля определила актуальность проводимых исследований. Целью работы было установление особенностей пространственно-временной динамики биологической активности почв в фитогенном поле сосны обыкновенной на примере сосняка брусничного среднетаежной подзоны Карелии. Данная цель предусматривала решение следующих основных задач:
1) установить варьирование свойств лесных подстилок в зависимости от зоны фитогенного поля;
2) проследить сезонную динамику гидротермических свойств почв, сформировавшихся в различных функциональных зонах фитогенного поля дерева;
3) определить показатели активности биотической компоненты почв в зависимости от зоны фитогенного поля дерева.
Оценка пространственного варьирования свойств почвы ненарушенных лесных биогеоценозов, обусловленных влиянием фитогенного поля дерева, важна для понимания и сохранения биоразнообразия в таежных лесах и может быть эталоном качества природной среды в современных условиях увеличения антропогенной нагрузки, глобального изменения климата (Лукина и др., 2010).
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в период вегетации растений на протяжении 2017–2020 гг. в среднетаежной подзоне республики Карелии. Комплексные работы выполняли в сосняке брусничном заповедника “Кивач”. Почва – подзол иллювиально-железистый песчаный, сформированный на озерно-ледниковых отложениях (Классификация и диагностика …, 2004), отдельные химические характеристики которого приведены в табл. 1.
Таблица 1.
Свойства подзола иллювиально-железистого песчаного в различных функциональных зонах фитогенного поля дерева
| Зона фитогенного поля | Горизонт почв |
Мощность, см |
рНKCl | Нг | V | С |
|---|---|---|---|---|---|---|
| мг-экв/100 | % | % | ||||
| Ствол | O | 4.6 ± 0.5 | 3.2 ± 0.1 | 90 ± 10 | 4 ± 1 | 44.32 ± 1.42 |
| E | 2.2 ± 0.4 | 3.3 ± 0.1 | 6 ± 1 | 4.7 ± 0.7 | 0.86 ± 0.06 | |
| BF | 18.3 ± 0.7 | 4.3 ± 0.1 | 4.4 ± 0.6 | 4.9 ± 0.5 | 0.64 ± 0.06 | |
| Крона | O | 5.1 ± 0.6 | 3.2 ± 0.1 | 100 ± 10 | 7 ± 1 | 44.00 ± 2.64 |
| E | 2.0 ± 0.3 | 3.5 ± 0.2 | 5.1 ± 0.7 | 5.2 ± 0.7 | 0.90 ± 0.09 | |
| BF | 17.9 ± 0.7 | 4.4 ± 0.1 | 3.9 ± 0.4 | 8 ± 3 | 0.67 ± 0.06 | |
| Окно | O | 4.1 ± 0.4 | 3.3 ± 0.1 | 99 ± 5 | 6 ± 1 | 44.98 ± 1.49 |
| E | 2.2 ± 0.4 | 3.4 ± 0.1 | 5 ± 2 | 6 ± 1 | 0.84 ± 0.07 | |
| BF | 18 ± 1 | 4.4 ± 0.1 | 3.2 ± 0.2 | 10 ± 3 | 0.65 ± 0.04 |
В зоне средней тайги в связи с медленным разложением органического вещества, поступающего с опадом, происходит активное формирование мортмассы – лесной подстилки. Запас лесной подстилки в разных зонах фитогенного поля дерева определяли однократно рамкой 25 × 25 см в июле 2018 г. в 12-кратной повторности. Лесная подстилка состояла из трех подгоризонтов разной степени разложения – O(L), O(F), О(Н), каждый из которых учитывали отдельно (Богатырев, Фомина, 1991; Орлова и др., 2015). Определение сухой массы подстилки проводили путем сложения массы всех подгоризонтов с учетом их влажности. Годовое поступление опада определяли суммированием поступающей растительной массы в опадоуловители (диаметром 70 см), которые были установлены в 6–9 кратной повторности (Руководство …, 2013). Учет массы поступающего опада проводили ежемесячно.
Интенсивность круговорота органического вещества в лесном биогеоценозе можно охарактеризовать с помощью коэффициента разложения опада (k), представляющего собой отношение годового количества опада (т/га) к массе подстилки (т/га). Величина, обратная этому коэффициенту, представляет собой среднее время оборачиваемости подстилки (1/k), выраженное в годах (Olson, 1963). Также одним из информативных показателей скорости деструктивных процессов стал опадо-подстилочный коэффициент, рассчитанный как отношение массы лесной подстилки к количеству ежегодного поступающего опада (Смольянинов, 1969).
С целью анализа температурного режима почв круглогодично фиксировали температуру воздуха и разных слоев почвы термологгерами (iButton DS1925L-F5). На основании первичных данных были получены среднесуточные значения температуры воздуха и почв на глубинах (0.5, 5, 10, 20 см) и рассчитаны соответствующие среднемесячные значения. Затем для генерализованной характеристики теплового режима почв была найдена сумма активных температур (>5°С и >10°С) на всех изучаемых глубинах.
Отбор проб для микробиологических анализов проводился по традиционной методике (Мишустин и др., 1968; Методы почвенной микробиологии …, 1991). Исследовали верхний слой (0–10 см) почвы, включающий органогенный горизонт – лесную подстилку (О) и подзолистый горизонт (Е). Учет численности почвенных гетеротрофных микроорганизмов, участвующих в трансформации азот- и углеродсодержащих соединений, проводили методом посева на плотные элективные питательные среды (Методы почвенной микробиологии …, 1991). Количество бактерий, использующих органические формы азота, учитывали на мясо-пептонном агаре (МПА), ассимилирующих минеральный азот – на крахмало-аммиачном агаре (КАА), бацилл – на среде МПА + сусло-агар, олигонитрофилов – на среде Эшби, олиготрофных микроорганизмов – на почвенном агаре (ПА). Численность актиномицетов определяли на КАА. Комплекс целлюлозоразрушающих микроорганизмов оценивали на среде Гетчинсона. Микроскопические грибы подсчитывали на сусло-агаре с лимонной кислотой. Для характеристики биологической активности почв в пространственно-временном аспекте устанавливали целлюлозолитическую способность почв в модельном полевом опыте in situ (Методы почвенной микробиологии …, 1991).
Определение интенсивности дыхания в полевых условиях проводили камерным методом в сезонной динамике (2 раза в месяц) с мая по ноябрь включительно. Использовали серию измерительных камер, представляющих собой врезанные в почву без удаления подстилки ПВХ трубы диаметром 10 см. Оценку вклада дыхания корней в общую эмиссию диоксида углерода с поверхности почв проводили методом удаления корней (Кузяков, Ларионова, 2006). Измерения проводили с мая 2017 г. по ноябрь 2020 г. Одновременно с измерением эмиссии СО2 с поверхности почв контролировали температуру (Тп) почвы на глубине 0–5 см, 5–15 см, температуру воздуха (Твозд) припочвенного слоя 1.5 см и на высоте 1.5 м, а также влажность лесной подстилки и минеральной части почвы на глубине 5–15 см.
Для характеристики температурной чувствительности почвенного дыхания использовали температурный коэффициент Q10 (коэффициент Вант-Гоффа), который рассчитывали исходя из уравнения экспоненты: Q10 = exp(10k).
Содержание углерода микробной биомассы (Cмик) определяли методом субстрат-индуцированного дыхания (СИД), которое оценивали по скорости начального максимального дыхания микроорганизмов после обогащения почвы глюкозой и инкубации в течение 1.5–2 ч. при температуре 22°С (Anderson, Domsch, 1978; Евдокимов, 2018). Изменение концентрации СО2 регистрировали газоанализатором (на основе NDIR-сенсора фирмы SenseAir, Швеция).
Величину СИД рассчитывали по формуле: СИД = dС × 12 × Vфлак × 1000/m × 22.4 × t × 100, где СИД – субстрат-индуцированное дыхание, мкг С/г субстрата/ч; dC – изменение концентрации СО2 c учетом нулевого значения, объемные %; Vфлак – объем флакона, мл; t – время инкубации, ч; m – масса абсолютно сухой почвы, г.
Углерод микробной биомассы рассчитывали по формуле: Cмик = СИД × 40.04 + 0.37 (Anderson, Domsch, 1978), где Cмик – углерод микробной биомассы, мкг С/г почвы; СИД – субстрат-индуцированное дыхание, мкг С–СО2/г почвы в час.
Для определения плотности размещения микориз были отобраны почвенные образцы в разных зонах фитогенного поля дерева (до 15 шт. в каждой зоне), из которых извлекали корни предпоследнего порядка общей длиной с каждого образца не менее 10 см. Затем образцы корневых систем фотографировали в высоком качестве и проводили анализ фотографий. Далее осуществляли пересчет числа сосущих окончаний на единицу длины несущего корня (Семенова, 1980).
Анализ полученных данных проводили с использованием классических статистических методов (программное обеспечение Statistica v.9.0) для расчета среднего, максимального и минимального значений, стандартного отклонения и коэффициента вариации. Кластерный анализ применяли для исследования сходства между различными свойствами с использованием определенной метрики, такой как евклидово расстояние (Berrueta, 2007). Результаты кластеризации были отображены с использованием древовидной диаграммы, показывающей структурную связь данных и выявляющей взаимосвязи различных признаков объектов и анализа их факторной структуры.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Поступление и трансформация растительного опада. Средние значения мощности лесных подстилок уменьшаются от приствольных возвышений к межкроновым пространствам. Запасы лесной подстилки в приствольной зоне и под кроной деревьев значительно не различаются и варьируют от 27.69 до 78.34 т/га. Данные учета массы лесной подстилки отражают характер накопления мортмассы вблизи стволов деревьев и под кроной, в межкроновом пространстве запасы подстилки значимо меньше (p = 0.0123) и составляют 27.69 ± 2.19 т/га. Коэффициент вариации запаса подстилки, обусловленный влиянием фитогенного поля дерева, равен 15–37%, а в пределах сосняка брусничного вариабельность запаса подстилки достигает 32%. Суммарное годовое поступление опада было максимальным в подкроновом пространстве (2.47–2.49 т/га), а минимальным – в “окне” (2.37 т/га). Ежегодное поступление опада, по полученным нами данным, в среднем составляет 2.44 т/га, что характерно для сосновых древостоев таежной зоны (Базилевич, Титлянова, 2008). Таким образом, наибольшие запасы лесной подстилки сконцентрированы под кронами деревьев, что напрямую связано с количеством поступающего опада (p = 0.00239). Наши расчеты показали, что время оборачиваемости лесной подстилки одинаково в приствольном повышении и в межкроновом пространстве и составляет 16 лет, в зоне под кроной и на краю кроны – 15 лет (табл. 2). Важная роль в биологическом круговороте наземных систем принадлежит деструктивным процессам, поскольку преобладающая часть биологической продукции трансформируется в форму детрита под действием различных агентов разложения (Krishna, Mohan, 2017). Рассчитанный опадо-подстилочный коэффициент варьирует от 6.35 до 24. Полученные данные свидетельствуют о медленных процессах минерализации поступающего опада.
Таблица 2.
Коэффициент разложения опада в различных зонах фитогенного поля дерева в сосняке брусничном (среднее ± SE)
| Зона фитогенного поля | Запас лесной подстилки, т/га | Поступление опада за год, т/га | Опадо-подстилочный коэффициент | k, 1/год |
Оборачиваемость подстилки, лет |
|---|---|---|---|---|---|
| Ствол | 39.83 ± 4.20 | 2.47 ± 0.017 | 11.91 ± 0.67 | 0.062 | 16.1 |
| Крона | 37.61 ± 1.57 | 2.49 ± 0.017 | 13.21 ± 0.77 | 0.066 | 15.1 |
| Окно | 27.69 ± 2.19 | 2.37 ± 0.018 | 12.83 ± 1.67 | 0.062 | 16.1 |
Температурно-влажностный режим почв. Анализ сезонной динамики среднесуточной температуры почвы на глубине 0–5 см и 5–15 см в различных зонах фитогенного поля дерева сосняка брусничного показал их сходный характер с небольшими расхождениями в горизонте лесной подстилки, с глубиной эти различия нивелировались. Наибольшие различия температуры минеральных горизонтов почвы в разных зонах фитогенного поля дерева наблюдались в период с мая по июль, для лесной подстилки этот период был несколько короче (май–июнь). В мае в большей степени прогревалась почва в межкроновом пространстве. В июне температура почвы в зоне “окно” также была выше по сравнению с зоной приствольного повышения: на 2°С в органогенном горизонте и на 1°С в минеральном слое на глубине 5–15 см. В июле при общей тенденции прогрева воздуха и почвы значимых различий (p < 0.05) по температуре органогенного горизонта почв не было выявлено, однако в минеральных горизонтах температура почвы в межкроновом пространстве оставалась на полградуса выше, чем в приствольной зоне. С августа по октябрь температура почвы выравнивалась как в горизонтальном, так и вертикальном направлении.
Анализ суммы температур воздуха, превышающих +5 и +10°C, не выявил существенной разницы между участками различных зон фитогенного поля дерева (рис. 1). Почвы подкронового пространства прогревались в теплое время года несколько сильнее соседних участков, особенно на глубине 10 см. Установлено, что крона нивелирует температурный режим почвы, снижая максимальные и увеличивая минимальные значения температуры верхнего слоя почвы.
Рис. 1.
Сумма активных температур почвы Тп > +5°C (а) и Тп > +10°C (б) на различных глубинах в различных зонах фитогенного поля дерева (ствол, крона, окно) сосняка брусничного за вегетационный период.
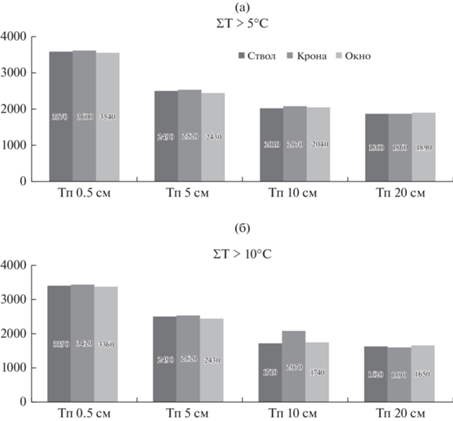
Высокое содержание органического вещества в верхнем горизонте почв определяет ее гидрофильные свойства: способность удерживать влагу, выполнять защитную функцию от нехватки воды в засушливый летний период. Относительная влажность почвы в осенне-весенний период была максимальной в органогенном горизонте и варьировала от 65 до 77%, в минеральных горизонтах она была ниже и составляла 20–24%. В летние месяцы относительная влажность составляла 50–65% для лесной подстилки, 10–15% для минеральной части корнеобитаемого слоя почвы.
СО2-эмиссионная активность почв. Интегральным показателем почвенной биоты является биологическая активность почв, которая оценивается по целому ряду показателей. Одним из информативных показателей, способных оценить активность микробиоты в пространственно-временном аспекте, служит дыхательная активность почв (Ананьева, 2003; Ananyeva et al., 2020) Эмиссия диоксида углерода из почвы является наиболее чувствительной составляющей углеродного цикла, которая быстро реагирует на любой абиотический и биотический стресс. Потоковая часть цикла углерода отражает современное состояние климата в регионе исследования и характеризует отклик углеродного баланса на изменения гидротермического режима (Larionova et al., 2010; Kurganova et al., 2011). Годовые потоки СО2 из почв лесных экосистем характеризуются высокой временной вариабельностью. В нашем исследовании установлены значимые сезонные различия величины эмиссии СО2 с поверхности почвы (p < 0.001). Зона фитогенного поля дерева также достоверно значимо оказывала влияние на эмиссионные потоки СО2 из почвы (p = 0.0002). Так, в зависимости от сочетания погодных условий и величины годовых потоков из почвы в сосняке брусничном потоки СО2 в разных зонах фитогенного поля дерева могли отличаться более чем в 1.5 раза, составляя в межкроновом пространстве 5.6 ± 0.7 т С/га в год, и 8.5 ± 0.9 т С/га в год у ствола, и 7.7 ± 0.9 под кроной (табл. 3 ), при этом влияние ФПД не перекрывает сезонные флуктуации значений почвенного дыхания. Различия величины корневого дыхания между участками разных зон фитогенного поля дерева прямо пропорционально связаны с корненасыщенностью почв данных участков (p = 0.046, R2 = 0.84). Условия зон фитогенного поля значимо (p < 0.05) не влияют на плотность размещения микориз на корне. Это, по-видимому, связано с тем, что мицелий гриба представлен во всех зонах фитогенного поля, что позволяет формировать микоризу, а изменения свойств почвы внутри изучаемых зон, в свою очередь, не оказывают существенного влияния на микоризообразование.
Таблица 3.
Вклад корневого дыхания в общую эмиссию СО2 с поверхности почв
| Зона фитогенного поля дерева | R микробное, г С-CO2/м2 сут | R корневое, г С-CO2/м2 сут | Вклад дыхания корней, % |
|---|---|---|---|
| Ствол | 1.97 ± 0.12a | 0.67 ± 0.10a | 24 ± 3a |
| Крона | 2.40 ± 0.24a | 1.44 ± 0.24b | 32 ± 4b |
| Окно | 1.44 ± 0.24b | 0.50 ± 0.17b | 45 ± 5b |
Динамика почвенного дыхания в течение вегетационного сезона была тесно связана с температурой приземного слоя воздуха (R2 = 0.35, p < 0.0001) и более существенно – с температурой почвы на глубине 10 см (R2 = 0.50, p < 0.0001). Эти зависимости хорошо аппроксимировались экспоненциальным уравнением 1-го порядка: R = expkT. Значение температуры почвы объясняло 41–56% изменчивости интенсивности почвенного дыхания в течение вегетационного периода.
Структуры микробного сообщества почв. Анализ эколого-трофической структуры микробного сообщества верхнего органогенного горизонта почв подтвердил влияние ФПД на вариабельность микробиологических свойств почв (табл. 4). Тенденция более высоких показателей микробиологической активности почв прослеживается в приствольной и подкроновой зоне дерева. Установили, что наибольшую численность имеют микроорганизмы, использующие органические формы азота. При этом отмечали высокие показатели численности спорообразующих форм бактерий на данном участке. Как известно, спорообразующие микроорганизмы не только переживают неблагоприятные факторы среды, но и являются участниками более глубоких превращений органического вещества в почве: выделяя экзоферменты, осуществляют гидролитическое расщепление полипептидов, которые могут использовать другие участники процесса (Звягинцев и др., 2005). Резкое изменение численности бактерий – олигонитрофилов также отмечали по мере удаления от ствола дерева. Это может быть обусловлено трофической приуроченностью к “рассеянным” соединениям азота, образуемым при работе микроорганизмов-гидролитиков: получая ничтожно малые порции азота, они осуществляют несимбиотическую фиксацию азота, задерживая его в экосистеме.
Таблица 4.
Диапазон изменения численности микроорганизмов и количества углерода микробной биомассы почв в разных зонах фитогенного поля дерева
| Зона фито-генного поля дерева | Гори-зонт | Бактерии, использующие | Олиго-нитро- филы |
Олиго-трофы | Микроскопи-ческие грибы | КЦМ | Смик | ||
|---|---|---|---|---|---|---|---|---|---|
| N-NH2 | N-NH4 | ||||||||
| общие | споровые | ||||||||
| тыс. КОЕ/г почвы | мкг С/г почвы | ||||||||
| Ствол | О | $\frac{{2566 - 9944}}{{6726}}$ | $\frac{{358 - 2427}}{{1486}}$ | $\frac{{1040 - 9915}}{{6250}}$ | $\frac{{1988 - 10{\kern 1pt} 930}}{{8143}}$ | $\frac{{1216 - 3019}}{{1874}}$ | $\frac{{313 - 421}}{{350}}$ | $\frac{{0.3 - 3.5}}{2}$ | $\frac{{3800 - 4800}}{{4300}}$ |
| Е | $\frac{{110 - 247}}{{161}}$ | $\frac{{48 - 82}}{{58}}$ | $\frac{{215 - 517}}{{332}}$ | $\frac{{481 - 941}}{{685}}$ | $\frac{{45 - 331}}{{175}}$ | $\frac{{24 - 58}}{{41}}$ | $\frac{{0 - 2.8}}{2}$ | $\frac{{120 - 160}}{{140}}$ | |
| Крона | О | $\frac{{2700 - 8007}}{{6129}}$ | $\frac{{396 - 2181}}{{1324}}$ | $\frac{{1825 - 10{\kern 1pt} 351}}{{6902}}$ | $\frac{{2112 - 8558}}{{6408}}$ | $\frac{{909 - 1836}}{{1310}}$ | $\frac{{317 - 364}}{{344}}$ | $\frac{{0.3 - 8}}{5}$ | $\frac{{4400 - 5800}}{{5100}}$ |
| Е | $\frac{{105 - 126}}{{146}}$ | $\frac{{41 - 111}}{{68}}$ | $\frac{{156 - 307}}{{232}}$ | $\frac{{331 - 658}}{{525}}$ | $\frac{{59 - 331}}{{166}}$ | $\frac{{21 - 45}}{{36}}$ | $\frac{{0 - 2}}{1}$ | $\frac{{100 - 140}}{{120}}$ | |
| Окно | О | $\frac{{2715 - 5668}}{{3803}}$ | $\frac{{474 - 1238}}{{815}}$ | $\frac{{1982 - 7439}}{{4459}}$ | $\frac{{1219 - 5959}}{{4158}}$ | $\frac{{1342 - 3656}}{{2260}}$ | $\frac{{161 - 367}}{{269}}$ | $\frac{{0.3 - 4}}{3}$ | $\frac{{4200 - 5200}}{{4700}}$ |
| Е | $\frac{{124 - 192}}{{152}}$ | $\frac{{41 - 69}}{{53}}$ | $\frac{{169 - 404}}{{257}}$ | $\frac{{659 - 828}}{{727}}$ | $\frac{{30 - 450}}{{257}}$ | $\frac{{44 - 58}}{{52}}$ | $\frac{{0.1 - 4}}{2}$ | $\frac{{110 - 150}}{{130}}$ | |
В микробном сообществе присутствовали группы, чувствительность которых к изменению микроусловий почвенной среды была относительной. Отмечали, что диапазон численности олиготрофов не претерпевал заметных изменений в зависимости от ЗФП. Возможно, что это связано с тем, что, несмотря на изменение объема хвойного опада, поступающего на почву в разных частях фитогенного поля дерева, его состав не претерпевал резких изменений. Можно отметить высокое содержание лигнифицированной массы, трудноминерализуемых азот- и углеродсодержащих соединений, его высокую кислотность (Колмогорова, Уфимцев, 2018). Данные соединения являются предшественниками прогумусовых соединений, которые остаются неизменными: преобладают фульвокислоты и продукты их синтеза (Чуков, 2001). Олиготрофы – это сборная в таксономическом отношении группа микроорганизмов, способная использовать для своих конструктивных целей периферические, легко отщепляемые цепи фульвокислот. Резкого изменения численности комплекса целлюлозоразрушающих микроорганизмов также не выявили, что обусловлено более медленным вовлечением их в процесс трансформации органического вещества, зависимости синтеза целлюлозолитических ферментов от физико-химических педоусловий.
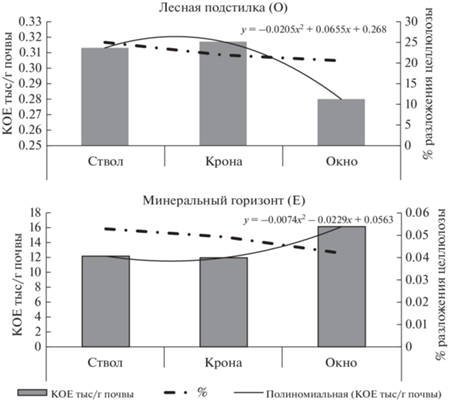
В минеральном горизонте почв резкого изменения эколого-трофической структуры микроорганизмов не отмечено. Изменение диапазона численности отмечали для бактерий, утилизирующих минеральные формы азота, группы олигонитрофилов. Столь согласованное изменение численности важнейших эколого-трофических групп микроорганизмов легко объясняется участием их на заключительных этапах превращения азотсодержащих веществ и меньшей зависимостью от содержания элементов минерального питания. Последние хорошо утилизируются мицелиальными прокариотами, численность которых была низкой во все сроки проведения наблюдений. Исследование показало, что целлюлозолитическая способность органогенного и верхнего минерального горизонтов почв, оцениваемая по скорости разложения целлюлозного полотна, изменяется однонаправленно, снижаясь в ряду ствол > крона > окно (рис. 2).
Рис. 2.
Изменение целлюлозолитической способности почв (%) и численности комплекса целлюлозоразрушающих микроорганизмов (КОЕ тыс./г почвы) в почвах различных зон ФПД сосняка брусничного.
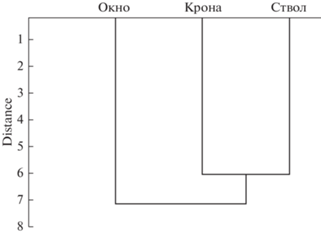
Влияние фитогенного поля дерева на свойства почв. Провели оценку влияния зон фитогенного поля на вариации микробиологических показателей органогенного горизонта почв (рис. 3). В иерархическом кластерном анализе исследуемые показатели были сгруппированы на основе расположения почв относительно функциональной зоны фитогенного поля. В результате кластерного анализа, проведенного на основе изучаемых свойств, были получены два кластера. Один из них включал функциональную зону “окно”, другой – “ствол” и “крона”. Подкроновая область фитогенного поля дерева была четко сгруппирована и удалена от межкроновой зоны. При этом при детальном анализе всей древовидной дендрограммы видна небольшая разница между свойствами почв различных функциональных зон, что обусловлено, возможно, комплексным влиянием факторов среды на свойства почв в каждой функциональной зоне.
Рис. 3.
Дендрограмма, полученная на основе иерархического кластерного анализа исследуемых свойств почв в различных функциональных зонах кроны дерева.
Сравнительный анализ характеристик биотической компоненты почв, проведенный на основе представленных в работе данных и результатов других исследователей, позволили заключить, что изучаемые компоненты биоты почв исследуемой территории имеют сходства с почвами таежной зоны Европейского Севера, а также свои особенности. Сходство заключается в высокой вариабельности микробиологических показателей в зависимости от климатических условий года (Chen et al., 2003; Оnwuka, Mang, 2018). Численность и активность биоты почв также зависит от состава опада, поступающего на почву (Ball, 1997; Krishna, Mohan, 2017). Можно утверждать, что сложная комплементарность между различными блоками биоты и растениями определяет и однонаправленность биохимических процессов, происходящих в почве (de Freitas Pereira, 2018). Это является генетически детерминированным процессом, что свидетельствует об общем плане функционирования биоты, формировании единой трофической системы, положительные связи которой обеспечивают выполнение экосистемных услуг (Rooney, Mc Cann, 2012). Сходство функционирования биоты таежной зоны определяется количеством поступающей в экосистему солнечной энергии, которая является “триггером”, пусковым механизмом метаболических реакций. В лесные экосистемы поступает разное количество солнечной энергии, следовательно, прогревание почв неодинаковое и скорость реакций также различна (Pfeifer et al., 2018).
Отличие функционирования биоты изучаемых нами участков определяется и спецификой почвообразующих пород, бедных по содержанию щелочных и щелочноземельных металлов (Морозова, 1991). Холодный гумидный климат тормозит биологическое выветривание минералов почвенной толщи, в этой связи в природном круговороте веществ циркулирует низкое содержание микроэлементов. Последнее, как известно, необходимо для нормального функционирования микробиоты, определяет ферментативную активность (Tabak et al., 2005). Трофические связи, которые могут формироваться между участниками процесса превращения органического вещества, эфемерные и короткие. В этой связи формируется гумус типа мор, структура которого более простая и, возможно, более неустойчивая к антропогенному воздействию (Ромашкевич, Герасимова, 1982). Низкая численность микроорганизмов целлюлозолитического комплекса связана наряду с гидротермическими условиями с невысоким содержанием микроэлементов в почве, что определяет заторможенность минерализации органического вещества, круговорот углерода.
В целом представленный перечень сходств и отличий биоты почв неполный, раскрывает лишь отдельные, наиболее главные, моменты. Однако они позволяют приблизить нас к познанию основ функционирования биоты почв ненарушенных лесов. Последнее важно для установления порога устойчивости наземных экосистем к различным видам воздействий: глобальному потеплению климата, аэротехногенному загрязнению и другим видам антропогенного воздействия (Herman, Clarholm, 1998).
ЗАКЛЮЧЕНИЕ
Анализ эколого-трофической структуры микробного сообщества почв сосняка брусничного среднетаежной подзоны Карелии показал, что наиболее высокой численностью и функциональной активностью отличаются верхние органогенные горизонты, в минеральных же горизонтах численность резко снижается. В микробном сообществе превалируют микроорганизмы, осуществляющие превращение азота, стратегия их выживания направлена на утилизацию рассеянных форм азота. Численность комплекса целлюлозоразрушающих микроорганизмов низкая, что определяет заторможенность процессов деструкции органического вещества, формирование лесных подстилок. По степени обогащения микроорганизмами исследуемую почву можно охарактеризовать как “бедную” и “среднюю”.
Эдафические условия изменяются на фоне прижизненных выделений растений- эдификаторов. Наиболее выраженные изменения микробной трансформации органического вещества происходят в верхнем органогенном горизонте почв в зоне ствол-крона, по мере отделения от дерева активность снижается. В верхнем минеральном горизонте изменение эколого-трофической структуры микробного сообщества менее выражено, что обусловлено низким содержанием гумуса, элементов питания, формированием неблагоприятных условий для их развития. Установлено, что основным экологическим фактором, влияющим на скорость деструкционных процессов и величину почвенных потоков СО2 в сосняке брусничном является зона ФПД, при этом факторами, определяющими сезонную динамику почвенной эмиссии СО2, являются температура воздуха и почвы. Различия в микробиологических показателях почв обусловливают особенности круговорота элементов-биофилов, пространственную неоднородность эдафических условий. Последнее влияет на продукционную способность древостоя, его устойчивость к неблагоприятным природно-техногенным воздействиям.
Авторы выражают глубокую благодарность руководству Государственного природного заповедника “Кивач” за всестороннюю поддержку, а также сотрудникам института леса КарНЦ РАН Н.Н. Ивашовой и Ю.С. Кудиновой – аналитикам лаб. лесного почвоведения – за помощь в выполнении камеральных работ и сотрудникам ИФХ-иБПП РАН И.Н. Кургановой и В.О. Лопес де Гереню за научное консультирование в вопросах измерения эмиссии СО2 из почвы.
Список литературы
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М.: Наука, 2003. 223 с.
Арчегова И.Б., Кузнецова Е.Г. Влияние древесных растений на химический состав атмосферных осадков в процессе восстановления среднетаежных лесов // Лесоведение. 2011. № 3. С. 34–43.
Базилевич Н.И., Титлянова А.А. Биотический круговорот на пяти континентах: азот и зольные элементы в природных экосистемах. Новосибирск: Изд-во СО РАН, 2008. 376 с.
Богатырев Л.Г., Фомина Т.В. Характеристика подстилок сосняков Присурского лесного массива // Вестник МГУ. Серия 17. Почвоведение. 1991. № 3. С. 28–39.
Евдокимов И.В. Методы определения биомассы почвенных микроорганизмов // Russian J. Ecosystem Ecology. 2018. Т. 3. № 3. С. 1–20.
Загуральская Л.М. Микробиологические аспекты мониторинга лесных почв Карелии в условиях заповедного режима // Лесоведение. 2003. № 4. С. 42–46.
Звягинцев Д.Г., Бабьева И.П., Зенова Г.М. Биология почв. М.: Изд-во МГУ, 2005. 448 с.
Калинин М.И. Корневедение. М.: Экология, 1991. 173 с.
Карпачевский Л.О., Просвирина В.П. О динамике строения почвенного покрова в лесных биогеоценозах // Почвоведение. 1989. № 5. С. 94–103.
Карпачевский Л.О., Зубкова Т.А., Пройслер Т., Кеннел М., Гитл Г., Горчарук Н.Ю., Минаева Т.Ю. Воздействие полога ельника сложного на химический состав осадков // Лесоведение. 1998. № 1. С. 50–59.
Карпечко Ю.В., Лозовик П.А., Потапова И.Ю. Трансформация химического состава атмосферных осадков лесной растительностью // Фундаментальные проблемы воды и водных ресурсов: труды IV Всероссийской научной конференции. М.: ИВП РАН, 2015. С. 155–158.
Карпечко Ю.В., Лозовик П.А., Федорец Н.Г. Баланс химических элементов в сосняке и ельнике Карелии // Лесоведение. 2018. № 2. С. 107–118.
Колмогорова Е.Ю., Уфимцев В.И. Некоторые особенности химического состава опада сосны обыкновенной, произрастающей в условиях породного отвала // Успехи современного естествознания. 2018. № 11(2). С. 267–272.
Кузяков Я.В., Ларионова А.А. Вклад ризомикробного и корневого дыхания в эмиссию СО2 из почвы (обзор) // Почвоведение. 2006. № 7. С. 842–854.
Лукина Н.В., Горбачева Т.Т., Никонов В.В., Лукина М.А. Пространственная изменчивость кислотности почв в процессе техногенной сукцессии лесных биогеоценозов // Почвоведение. 2003. № 1. С. 32–35.
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва – растительность // Лесоведение. 2010. № 5. С. 45–56.
Медведева М.В., Бахмет О.Н., Мошкина Е.В., Солодовников А.Н., Зачиняева А.В. Пространственное варьирование биологической активности почв соснового древостоя среднетаежной подзоны Карелии // Проблемы окружающей среды и природных ресурсов. 2020. № 5. С. 35–47.
Методы почвенной микробиологии и биохимии / Под ред. Звягинцева Д.Г. М.: Изд-во МГУ, 1991. 303 с.
Мишустин Е.Н., Никитин Д.И., Востров И.С. Модернизация методов учета почвенного микронаселения и его активности // Микроорганизмы в сельском хозяйстве. Минск, 1968. С. 144–150.
Морозова Р.М. Лесные почвы Карелии. Л.: Наука, 1991. 184 с.
Никонов В.В., Лукина Н.В. Пространственно-временная изменчивость питательного режима Al–Fe-гумусовых подзолов бореальных лесов // Почвоведение. 2000. № 12. С. 1487–1495.
Орлова М.А., Лукина Н.В., Смирнов В.Э. Методические подходы к отбору образцов лесной подстилки с учетом мозаичности лесных биогеоценозов // Лесоведение. 2015. № 3. С. 214–221.
Ромашкевич А.И., Герасимова М.И. Микроморфология и диагностика почвообразования. М.: Наука, 1982. 127 с.
Руководство по комплексному мониторингу. Перевод с английского. М.: ФГБУ “ИГКЭ Росгидромета и РАН”, 2013. 153 с.
Семенова Л.А. Морфология микориз сосны обыкновенной в спелых лесах // Микоризные грибы и микоризы лесообразующих пород севера. Петрозаводск: Карельский филиал АН СССР, 1980. С. 103–132.
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Кузнецова Я.Д. Оценка внутрибиогеоценозной изменчивости лесных подстилок и травяно-кустарничковой растительности в еловых насаждениях // Почвоведение. 2020. № 1. С. 31–43.
Смольянинов И.И. Биологический круговорот веществ и повышение продуктивности лесов. М.: Лесная промышленность, 1969. 192 с.
Уранов А.А. Фитогенное поле // Проблемы современной ботаники. Т. 1. М.–Л.: Наука, 1965. С. 251–254.
Чуков С.Н. Структурно-функциональные параметры органического вещества почв в условиях антропогенного воздействия. СПб.: Изд-во СПб. ун-та, 2001. 216 с.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Шубин В.И. Микотрофность древесных пород, ее значение при разведении леса в таежной зоне. Л.: Наука, 1973. 264 с.
Ananyeva N.D., Sushko S.V., Ivashchenko K.V., Vasenev V.I. Soil microbial respiration in subtaiga and forest-steppe ecosystems of European Russia: field and laboratory approaches // Eurasian Soil Science. 2020. V. 53. № 10. P. 1492–1501.
Anderson J., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biology & Biochemistry. 1978. V. 10. № 3. P. 215–221.
Ball A.S. Microbial decomposition at elevated CO2 levels: effect of litter quality // Global Change Biol. 1997. V. 3. P. 379–386.
Boettcher S.E., Kalisz P.J. Single-tree influence on soil properties in the mountains of eastern Kentuky // Ecology. 1990. V. 71. № 4. P. 1365–1372.
Berrueta L.A., Alonso-Salces R.M., Héberger K. Supervised pattern recognition in food analysis // J. Chromatography A. 2007. V. 1158. P. 196–214.
Chen Q.S., Li L.H., Han X.G., Yan Z.D., Wang Y.F., Yuan Z.Y. Influence of temperature and soil moisture on soil respiration of a degraded steppe community in the Xilin River basin of Inner Mongolia // Acta Phytoecol Sinica. 2003. V. 27. № 2. P. 202–209.
Gersper P.L., Holowaychuk N. Some effects of stem flow from forest canopy trees on chemical properties of soils // Ecology. 1970. V. 52. № 4. P. 691–702.
De Freitas Pereira M. Secretome analysis from the ectomycorrhizal ascomycete Cenococcum geophilum // Frontiers in Microbiology. 2018. V. P. 1–17.
Herman D., Clarholm V. Trophic relationships in the soil microfood-web: Predicting the responses to a changing global environment // Global Change Biology. 1998 V. 4. № 7. P. 713–727.
Hokkanen T.J., Järvinen E., Kuuluvainen T. Properties of top soil the relationship between soil and trees in a boreal Scots pine stands // Silva Fennica. 1995. V. 29. № 3. P. 189–203.
Kurganova I.N., Lopes de Gerenyu V.O., Petrov A.S., Myakshina T.N., Sapronov D.V., Ableeva V.A., Kudeyarov V.N. Effect of the observed climate changes and extreme weather phenomena on the emission component of the carbon cycle in different ecosystems of the southern taiga zone // Doklady Biological Sciences. 2011. V. 441. № 6. P. 412–416.
Krishna M.P., Mohan M. Litter decomposition in forest ecosystems: a review // Energy, Ecology and Environment. 2017. V. 2. № 4. P. 236–249.
Larionova A.A., Kurganova I.N., Lopes de Gerenyu V.O., Zolotareva B.N., Yevdokimov I.V., Kudeyarov V.N. Carbon dioxide emissions from agrogray soils under climate changes // Eurasian Soil Science. 2010. V. 43. № 2. P. 168–176.
Lawrence D.J., Luckai N., Meyer W.L., Shahi C., Fazekas A.J., Kesanakurti P., Newmaster S. Distribution of white spruce lateral fine roots as affected by the presence of trembling aspen: root mapping using simple sequence repeat DNA profiling // Canadian Journal of Forest Research. 2012. V. 42. P. 1566–1576.
Liski J. Variation in soil organic carbon and thickness of soil horizons within a boreal forest stand – effect of trees and implications for sampling // Silva Fennica. 1995. V. 29. № 4. P. 255–266.
Olson J.S. Energy storage and the balance of producers and decomposers in ecological systems // Ecology. 1963. V. 44. P. 322–331.
Onwuka B., Mang B. Effects of soil temperature on some soil properties and plant growth // Adv Plants Agric Res. 2018. V. 8. № 1. P. 34‒37.
Ostonen I., Helmisaari H.-S., Borken W., Tedersoo L., Kukumägi M., Bahram M., Lindroos A.-J., Nöjd P., Uri V., Merilä P., Asi E., Lõhmus K. Fine root foraging strategies in Norway spruce forests across a European climate gradient // Global Change Biology. 2011. V. 17. № 12. P. 3620–3632.
Pfeifer M., Gonsamo A., Woodgate W. et al. Tropical forest canopies and their relationships with climate and disturbance: results from a global dataset of consistent field-based measurements // Forest Ecosystem. 2018. V. 5. № 7. https://doi.org/10.1186/s40663-017-0118-7
Rooney N., McCann K.S. Integrating food web diversity, structure and stability // Trends in Ecology & Evolution/ 2012. V. 27. № 1. P. 40–46.
Tabak H.H., Lens P., van Hullebusch E.D., Dejonghe W. Developments in bioremediation of soils and sediments polluted with metals and radionuclides – 1. Microbial processes and mechanisms affecting bioremediation of metal contamination and influencing metal toxicity and transport // Reviews in Environmental Science and Bio/Technology. 2005. V. 4. № 3. P. 115–156.
Taylor A.F., Martin F., Read D.J. Fungal diversity in ectomycorrhizal communities of Norway Spruce (Picea abies (L.) Karst) and Beech (Fagus sylvatica L.) along north-south transects in Europe // Ecological Studies. 2000. V. 142. P. 343–365.
Дополнительные материалы отсутствуют.