

Лесоведение, 2022, № 6, стр. 713-726
Видовое и ценотическое разнообразие сообществ пойменных лесов на Северо-Востоке России
В. Ю. Нешатаев a, *, В. Ю. Нешатаева b, Н. В. Синельникова c, К. И. Скворцов b
a Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
194021 Санкт-Петербург, Институтский пер., д. 5, Россия
b Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Профессора Попова, д. 2, Россия
c Институт биологических проблем Севера ДВО РАН
685000 Магадан, ул. Портовая, д. 18, Россия
* E-mail: vneshatayeva@binran.ru
Поступила в редакцию 01.04.2022
После доработки 30.05.2022
Принята к публикации 07.06.2022
- EDN: HEALIL
- DOI: 10.31857/S0024114822060079
Аннотация
Для исследования видового разнообразия сообществ пойменных лесов Северо-Востока РФ использовано 177 геоботанических описаний, выполненных на севере Корякского округа, северо-востоке Магаданской области и юге Чукотского АО. Цель работы – анализ видового и ценотического разнообразия пойменных лесов в зависимости от условий местообитания и положения в сукцессионных рядах. При геоботанической характеристике сообществ чозениевых (чозения толокнянколистная (Chosenia arbutifolia)), тополевых (тополь душистый (Populus suaveolens)), ивовых (ива удская (Salix udensis), ива Шверина (S. schwerinii)) и ольховых (ольха пушистая (Alnus hirsuta)) лесов использована эколого-фитоценотическая классификация. Ценотическое разнообразие пойменных лесов представлено 19 ассоциациями, объединенными в 5 серий. Приведена характеристика флористического состава и структуры сообществ пойменных лесов, рассчитаны индексы Шеннона и Пиелу, средние показатели видового богатства, проективного покрытия видов и сомкнутости ярусов. По сравнению с лесами п-ова Камчатка, пойменные леса Северо-Востока РФ характеризуются невысоким синтаксономическим разнообразием и представлены флористически обедненными сообществами. Обсуждается структура и динамика сообществ пойменных лесов. Вычислены показатели видового разнообразия и выравненности для каждого из изученных сообществ. Наименьшим значением индекса Шеннона характеризуется пионерный чозенник редкотравный, встречающийся на молодых галечниках; а наибольшим значением – тополевник травяно-зеленомошный, занимающий наиболее высокие уровни поймы. Остальные ассоциации показали промежуточные значения индекса Шеннона. Выявлена тенденция к увеличению видового разнообразия в ходе аллювиальной сукцессии. Полученные данные позволяют предположить, что в ходе сукцессионных смен, связанных с уменьшением поемности и аллювиальности местообитаний, наблюдается увеличение видового разнообразия сообществ.
Пойменные леса Северо-Востока имеют большое природоохранное, берегозащитное, водоохранное и нерестовое значение. Для местного населения они являются важным источником деловой и дровяной древесины, а также сопутствующих растительных ресурсов. Распространение пойменных лесов в северотаежных и лесотундровых районах Северо-Востока РФ обусловлено более теплым микроклиматом в пределах пойм и надпойменных террас, а также существованием подрусловых таликов в долинах непромерзающих рек, благодаря которым создаются особо благоприятные, “оазисные” условия, способствующие развитию древесной растительности (Клюкин, 1970). На Северо-Востоке тополевые (тополь душистый) и чозениевые (чозения толокнянколистная) леса распространены по подрусловым таликам пойм в зоне многолетней мерзлоты. В поймах горных рек Охотии и Колымского нагорья пойменные леса занимают узкие полосы вдоль русел. В различных районах Восточной Сибири и Дальнего Востока описаны различные типы пойменных лесов: тополевник хвощово-свидиновый, тополевник хвощовый (Тюлина, 1959), Populus suaveolens forests (Krestov, 2003), чозенник с тополем разнотравно-хвощовый (Москалюк, 1988), кустарниково-разнотравно-злаковые лиственнично-чозениевые леса (Полежаев, 2005), чозениево-тополевые леса с примесью лиственницы и белой березы вейниково-грушанково-хвощовые (Гаращенко, 1993), кустарниково-разнотравно-злаковые лиственнично-чозениевые леса (Котляров, 1971), тополевник свидиново-грушанковый (Осипов, 2002). Лесоводственные характеристики тополевых и чозениевых насаждений, их состав и условия местопроизрастания достаточно хорошо изучены на юге Дальнего Востока (Зархина, 1969а, б, 1986).
Чозениевые леса являются характерным компонентом растительного покрова региона, они широко распространены в поймах рек Дальнего Востока и Якутии (Шелудякова, 1943; Тюлина, 1959; Котляров, 1971; Miyawaki, 1988; Осипов, 2002; Krestov, 2003; Qian et al., 2003; Исаев, Кузнецова, 2010). Леса занимают острова низкого и среднего уровня (0.7–1.2 м), отмечены преимущественно на галечных аллювиях. В восточных районах Якутии чозениевые леса описаны на молодых галечно-песчаных отложениях поймы р. Неры (Бурцева, Дестякина, 1998). На востоке Якутии (Восточное Верхоянье, бассейн р. Хандыги) чозениевые и тополевые леса приурочены к укрытым от холодных ветров пойменным островам в верховьях и среднем течении горных рек (Куваев, 1956). Встречаются такие сообщества довольно редко, и некоторые исследователи (Яровой, 1939; Шелудякова, 1943) указывают на их реликтовый характер. Здесь отмечены чозениево-тополевые рощи на молодых песчаных и песчано-галечных аллювиях и сомкнутые чозениево-тополевые разнотравно-вейниковые леса с подлеском из ольхи кустарниковой (Alnus fruticosa). На западном побережье Камчатки тополевые и чозениевые леса имеют ограниченное распространение (Тюлина, 2001). Травостой пойменных лесов Центральной и Восточной Камчатки образован видами камчатского крупнотравья (лабазник камчатский (Filipendula camtchatica), крестовник коноплеволистный (Senecio cannabifolius), борщевик шерстистый (Heracleum dulce)) и крапива плосколистная (Urtica platyphylla); тополевники и ивняки вейниковые (вейник Лангсдорфа (Calamagrostis langsdorffii)) встречаются довольно редко (Нешатаева, 2009). Тополевники и ивняки крупнотравные распространены также в поймах горных рек Сахалина (Кабанов, 1940).
Леса из ивы Шверина многочисленны на Дальнем Востоке. На юге Дальнего Востока они формируют основу пойменной лесной растительности Амура (Ахтямов, 2001). Ивняки занимают острова низкого и среднего уровня (0.7–1.2 м), отмечены на суглинистых аллювиях в тыловых частях островов и по старичным понижениям. В Охотии леса из ивы Шверина менее распространены, в поймах крупных рек бассейна р. Тауй отмечены полидоминантные разнотравно-злаковые ивняки (Полежаев, 2005).
В пойменных лесах Магаданской обл. с использованием флористической классификации выделены ассоциации Salicetum schwerinii Sinelnikova 1995, Elymo-Chosenietum arbutifoliae Sinelnikova 1995 и Chosenio-Populetum suaveolentis Sinelnikova 1995, отнесенные к союзу Chosenion arbutifoliae Sinelnikova 1995 и порядку Populetalia laurifolia-suaveolentis Mirkin et al. 1986 класса Salicetea purpurea Moor 1958. Пойменные лиственничники отнесены к ассоциации Equiseto-Laricetum cajanderi Sinelnikova 1995 и союзу Roso acicularis-Laricion cajanderi Sinelnikova 2016 класса Vaccinio-Piceetea Br.-Bl. in Br.-Bl., Siss et Vlieger 1939. (Синельникова, 1995, 2016).
Показатели разнообразия растительности являются ее фундаментальными характеристиками. Различают α-разнообразие (разнообразие видов в пределах фитоценоза) и β-разнообразие (разнообразие растительных сообществ) (Розенберг, 2010). Мерой β-разнообразия является количество типов сообществ (низших единиц классификации растительности). В геоботанике наибольшее распространение для оценки α-разнообразия получили видовое богатство (число видов на ПП) и показатели выравненности: индекс Шеннона (H) и индекс Пиелу (Е) – отношение индекса Шеннона к его теоретически возможному максимальному значению (Pielou, 1975; Левич, 1980; Василевич, 2009, 2015, 2017, 2018; Розенберг, 2010; Василевич, Кессель, 2017).
Цель настоящей работы – проанализировать видовое и ценотическое разнообразие пойменных лесов Северной Корякии и сопредельных районов северо-востока Магаданской обл. и юга Чукотского АО, выявить их связь с условиями местообитания и положением сообществ в сукцессионных рядах. В задачи настоящего исследования входили геоботаническая характеристика пойменных лесов севера Корякского округа, определение количественных показателей видового и ценотического разнообразия и их анализ.
ОБЪЕКТЫ И МЕТОДИКА
Природные условия районов исследований. Исследования проведены в долинах рек Колымы, Омолон и Олой (Тенькинский и Среднеканский р-ны Магаданской обл. и Билибинский р-н Чукотского АО), в среднем течении р. Анадырь (Анадырский р-н Чукотского АО) и на севере Корякского округа – в долинах рек Пенжины, Белой, Ичигиннываям, Тыклаваям, Катальянайваям, Евъеинваям, Вывенки, Ветвей, Тылгаваям, Пылговаям (Пенжинский и Олюторский р-ны Камчатского края) (рис. 1).
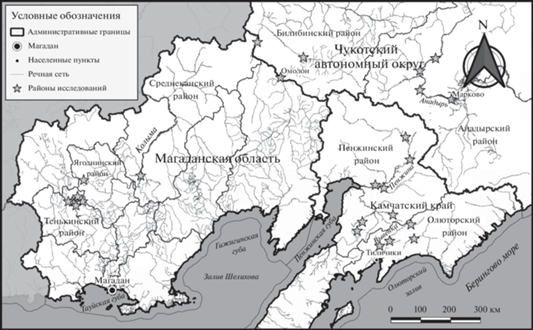
Для долины р. Колымы и ее притоков характерен континентальный климат с суровой продолжительной (220 дней) зимой; средняя температура января достигает –38.3°С, июля – +15.1°С. Среднее течение р. Анадырь находится в области умеренно-континентального климата; средняя температура января составляет –32°C, июля – +14°С. Годовая сумма осадков – 307 мм, продолжительность безморозного периода – 90–95 дней. Долина р. Пенжины изолирована от влияния Берингова моря; здесь климат континентальный, с продолжительной холодной зимой и коротким теплым летом. Средняя температура февраля составляет –25°С, июля – +13, +14°С. Вегетационный период – 100–110 дней. Сумма активных температур – 800–900°С. Годовая сумма осадков – 250 мм. В среднем течении р. Пенжины (пос. Аянка и Слаутное) климат более суровый, чем в ее низовьях (пос. Каменское и Манилы), находящихся под влиянием Охотского моря. Повсеместно распространена многолетняя мерзлота.
В межгорной депрессии Парапольский Дол, расположенной между Корякским нагорьем и Пенжинским хребтом, климат умеренно-континентальный. Средняя температура февраля равна –17, –20°С, июля – +11, +13°С. Сумма активных температур – 650–750°С. Годовая сумма осадков – 300 мм. В бассейне р. Вывенки климат континентальный, зима продолжительная (230 дней). Средняя температура января составляет –22°С; июля – +10, +12°С. Вегетационный период – около 100 дней. Годовая сумма осадков – 500–600 мм (Кондратюк, 1974).
По геоботаническому районированию (Нешатаева и др., 2020) районы исследований относятся к Корякской горной провинции Берингийской лесотундровой области и Колымской горной провинции подобласти Светлохвойных лесов Евразиатской таежной области. Зональная растительность представлена стланиковыми и кустарниковыми сообществами, образованными кедровым стлаником (Pinus pumila), ольховником кустарниковым (Alnus fruticosa) и березкой Миддендорфа (Betula middendorffii). Лесная растительность встречается в долинах крупных рек и представлена пойменными тополевниками (тополем душистым), чозенниками (чозенией толокнянколистной и ивняками (ивой удской, ивой Шверина), реже ольшаниками из ольхи пушистой. На юго-востоке Корякского нагорья в приморских районах до высот 100–250 м над ур. моря встречаются каменноберезовые рощи (береза Эрмана (Betula ermanii)). На высоких надпойменных террасах распространены ерниковые тундры, образованные березкой тощей (Betula exilis) и кустарничками. В переувлажненных депрессиях развиты осоково-пушицевые (осока траурная (Carex lugens), осока шаровидная (C. globularis L.), пушица влагалищная (Eriophorum vaginatum L.)), кочкарники с участием морошки (Rubus chamaemorus L.), подбела обыкновенного (Andromeda polifolia L.), клюквы мелкоплодной (Oxycoccus microcarpus) и мхи (сфагнум ленский (Sphagnum lenense), сфагнум Руссова (S. russowii), аулакомниум болотный (Aulacomnium palustre), дикранум удлиненный (Dicranum elongatum)). В горах до 400–500 м преобладают сообщества кедрового стланика в сочетании с кустарничковыми (голубикой обыкновенной (Vaccinium uliginosum L.), брусникой обыкновенной (V. vitis-idaea L.)), багульником стелющимся (Ledum decumbens), водяникой черной (Empetrum nigrum L.)) и ягельными (кладонией лесной (Cladonia arbuscula), кладонией оленьей (C. gangiferina L.), кладонией звездчатой (C. Stellaris) и др.) тундрами. На высотах 500 м и выше, на крутых склонах, вершинах и гребнях хребтов, преобладают каменистые осыпи и россыпи с разреженными группировками петрофитов и пятнами накипных эпилитных лишайников.
Методы полевых исследований. Использовали детально-маршрутные методы с закладкой пробных площадей (ПП) размерами 20 × 20 м. В поймах рек на севере Корякского округа было заложено 30 ПП, на северо-востоке Магаданской обл. и юге Чукотского АО – 147 ПП. Таксацию древостоев проводили глазомерно-инструментальным методом. Применяли оптический высотомер, возрастной бур Пресслера и рулетку. На каждой ПП выявляли полный видовой состав сосудистых растений, мохообразных и лишайников; определяли проективное покрытие для каждого вида и яруса, сомкнутость древостоя, среднюю и максимальную высоту и диаметр деревьев по элементам леса, высоту и сомкнутость подроста и подлеска. На каждой ПП закладывали 5–10 почвенных прикопок глубиной 0.3 м и почвенный разрез глубиной 0.7–1.5 м, выполняли описание почвенного профиля. Определяли мощность и характер генетических почвенных горизонтов и их характеристики, в т.ч. мощность свежего наилка, являющегося показателем аллювиальности.
Поймы крупных рек включают несколько уровней: низкая пойма (высота 0.5–1.2 м над уровнем воды в межень), средняя пойма (1.2–2.5 м), высокая пойма (2.5–3.5 м). Для каждой ПП устанавливали превышение уровня пойменной террасы над уровнем воды в реке в межень (далее – уровень террасы). Этот показатель косвенно характеризует поемность – длительность затопления во время половодья. Косвенным показателем уменьшения поемности и аллювиальности также является развитие кустарникового и мохового ярусов.
Эколого-фитоценотическая классификация пойменных лесов Северной Корякии и Камчатки разработана нами ранее (Нешатаева, 2009; Нешатаева и др., 2017, 2018). Принципы классификации лесной растительности В.Н. Сукачёва (1951) приняты в качестве национального стандарта РФ (ГОСТ 18486-87, Лесоустроительная …, 2018). Основной единицей классификации является тип лесорастительных условий (ЛРУ), который выделяется по почвенно-гидрологическим и высотно-поясным условиям, индикаторами которых являются доминаты и экологические группы растений. В системе фитоценологической номенклатуры типу ЛРУ соответствует цикл ассоциаций, а его подчиненным синтаксонам – серия ассоциаций (Нешатаев, 2001). Согласно В.Н. Сукачёву (1945) и его последователям (Гельтман, 1982 и др.), для каждого типа леса может быть выделена одна или несколько ассоциаций (или субассоциаций), отражающих варьирование фитоценозов в пределах типа леса, обусловленное возрастными изменениями сомкнутости и продуктивности древостоя, вариациями эдафических условий, освещенности и др. Классификацию ЛРУ проводили путем упорядочивания геоботанических описаний ПП по степени увлажнения, поемности и аллювиальности местообитаний. Затем описания группировали в фитоценотических таблицах по сходству ЛРУ и индицирующих их видов растений и выделяли серии ассоциаций. В пределах серий выделяли ассоциации с учетом преобладающих видов древесного яруса. Латинские названия синтаксонов даны в соответствии с Проектом Всероссийского кодекса фитоценологической номенклатуры (Нешатаев, 2001). Номенклатура видов сосудистых растений приведена по работе (Сосудистые …, 1985–1996), мохообразных – (Ignatov et al., 2006), лишайников – (Andreev et al., 1996). Методическое руководство “Классификация и диагностика почв России” (2004) послужило источником для номенклатуры почв. При характеристике синтаксонов использовали средние арифметические и их стандартные ошибки, равные корню квадратному из частного от деления дисперсии на количество ПП, отнесенных к данному синтаксону.
Методы оценки видового разнообразия. Разнообразие сообществ пойменных лесов оценивали по следующим показателям: 1) число видов, встреченных в каждом ярусе, в составе внеярусной растительности (эпифиты) и в целом для всего сообщества; 2) индекс Шеннона, рассчитанный для древостоя и подроста, подлеска, травяно-кустарничкового (ТКЯ) и мохово-лишайникового (МЛЯ) ярусов, сумма индексов Шеннона для всех ярусов;
$H = --\sum p_{i}^{*}{\kern 1pt} {{\log }_{2}}{\kern 1pt} {{p}_{i}}$, i = 1…N, где pi – вероятность i-ого события из полной группы событий, когда ∑pi = 1, в геоботанике соответствует проективному покрытию или сомкнутости крон по ярусам, выраженным в долях от 1;
3) индекс выравненности Пиелу (Pielou, 1975), рассчитанный для каждого яруса и в целом для всего сообщества. Индекс выравненности E рассчитан по формуле:
E = H/Нmax, где Нmax – теоретически возможное максимальное значение H-функции, наблюдаемое при одинаковом проективном покрытии всех N видов, встреченных в сообществе, и в общем проективном покрытии, равном 100%:
Следует отметить, что в одновидовых ярусах (N = 1) или при их отсутствии (N = 0) в формуле выравненности возникает деление на 0 (Log21 = 0) или на пустое множество (Log20 ∈ Ø). В этом случае индекс выравненности мы рассматриваем как предел, к которому стремится H/Нmax при условии, что он лежит в области значений от 0 до 1. Поэтому:
при N → 1; Log2N → 0; E → ∞ и 0 < E < 1 ⇒ ⇒ lim E = 1;
при N → 0; Log2N → -∞; E → –1 и 0 < E < 1 ⇒ ⇒ lim E = 0.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Всего в изученных сообществах пойменных лесов на 177 ПП отмечено 205 видов (не считая эпифитов), из них деревьев – 14, кустарников – 23, кустарничков – 3, трав – 118, эпигейных мохообразных – 36, эпигейных лишайников – 3. На пробной площади (400 м2) встречается от 4 до 43 видов, среднее число видов на ПП – 11. Распределение числа видов и индексов Шеннона и Пиелу по сериям ассоциаций приведены в таблице 1.
Анализ структуры древостоев и видового состава сообществ, отражающих условия местообитания, позволил выявить ценотическое разнообразие пойменных лесов районов исследований, представленное 6 формациями и 19 ассоциациями. Из них 15 ассоциаций были описаны ранее (Тихомиров, 1935; Воробьёв, 1937; Колесников, 1937; Биркенгоф, 1938; Васильев, 1956; Балмасова, Нешатаева, 1994; Нешатаева, 2009; Нешатаева и др., 2002, 2017, 2018), четыре ассоциации выделены по материалам исследований 2021 г. Ассоциации объединены в 5 серий, в зависимости от условий местообитания и динамического статуса сообществ.
Ниже приведена краткая характеристика серий ассоциаций, расположенных в порядке продвинутости сообществ в ходе аллювиальной сукцессии, обусловленной понижением базиса эрозии и, соответственно, уровня воды в реке и грунтовых вод по отношению к поверхности, на которой расположено сообщество. Сомкнутость ярусов, суммы проективных покрытий травяно-кустарничкового (ТКЯ) и мохово-лишайникового (МЛЯ) ярусов, число видов по ярусам и их сумма, а также индексы Шеннона и Пиелу для каждой серии ассоциаций приведены в таблице 1. При геоботанической характеристике серий ассоциаций константными мы считали виды, встреченные в сообществах данной серии на 50% и более ПП.
Серия ассоциаций Oligoherbosa – редкотравная (20 ПП). Включает три корреспондирующие ассоциации: Salicetum schwerinii oligoherbosum – ивняк из ивы Шверина редкотравный, Chosenietum oligoherbosum – чозенник редкотравный, Populetum suaveolentis – тополевник редкотравный. Сообщества описаны на участках со свежими песчано-галечными наносами, не успевающими зарастать в силу периодического поступления свежего аллювия. Средний уровень террасы – 1.6 ± 0.1 м (0.6–2.5). Почвы аллювиальные слоистые с фрагментами дерново-аллювиальных грунтово-глеевые сильно- и среднескелетные. Древостой разного возраста, высотой 10.8 ± 1.2 м (3–24). Здесь и далее для высоты древостоя и уровня террасы даны средние значения ± стандартная ошибка и размах варьирования. В подлеске константных видов нет. ТКЯ редкий, с покрытием 0.2–20%, константны злаки: вейник пурпурный (Calamagrostis purpurea) и пырейник длинноколосый (Elymus macrourus). Всего на 20 ПП отмечено 55 видов (здесь и далее – без учета стволовых эпифитов). В сообществах серии отмечено от 5 до 11 видов на ПП.
Серия ассоциаций Calamagrostidosa purpureae – вейниковая (95 ПП). Включает шесть ассоциаций: Alnetum hirsutae calamagrostidosum – ольшаник вейниковый, Salicetum udensis calamagrostidosum – удскоивняк вейниковый, Salicetum schwerinii calamagrostidosum – швериноивняк вейниковый, Laricetum cajanderi calamagrostidosum – лиственничник вейниковый, Populetum suaveolentis calamagrostidosum – тополевник вейниковый и Chosenietum calamagrostidosum – чозенник вейниковый. Сообщества вейниковой серии занимают средние и высокие уровни террас с превышением 2.1 ± 0.1 м (0.8–4.0 м). Почвы дерново-аллювиальные грунтово-глеевые сильно- и среднескелетные. Древостой разновозрастный, высотой 17.8 ± 0.6 м (5–27 м), в подлеске константна смородина печальная (Ribes triste). Сомкнутость подлеска менее 50%. ТКЯ высотой около 1 м, его общее проективное покрытие от 30 до 90%, доминирует вейник пурпурный. Мхи встречены на 35% ПП, их среднее покрытие менее 2%. На 95 ПП отмечено 155 видов растений. В сообществах серии зафиксировано от 4 до 40 видов на ПП, в среднем 11 видов. Наибольшее число видов (15–40) отмечено для чозенника вейникового, обогащенного видами высокотравья: недоспелкой копьевидной (Cacalia hastate L.), волжанкой двудомной (Aruncus dioicus). Наряду с этими эутрофными видами в сообществах встречаются также олиготрофные кустарнички: водяника черная (Empetrum nigrum L.), голубика обыкновенная, брусника обыкновенная, заселяющие отложения крупнопесчаного аллювия прошлых лет.
Серия ассоциаций Fruticoso-Herbosa – кустарниково-травяная (49 ПП). Включает пять ассоциаций: Salicetum udensis fruticoso-herbosum – удскоивняк кустарниково-травяной, Salicetum schwerinii fruticoso-herbosum – швериноивняк кустарниково-травяной, Laricetum cajanderi fruticoso-herbosum – лиственничник кустарниково-травяной, Populetum suaveolentis fruticoso-herbosum – тополевник кустарниково-травяной и Chosenietum fruticoso-herbosum – чозенник кустарниково-травяной. Уровень террасы – 1.9 ± 0.1 м (0.8–3.2). Почва дерново-аллювиальная песчано-галечная или супесчано-галечная грунтово-глеевая сильноскелетная. Возраст и состав древесного яруса (ДЯ) сильно варьирует, его высота – 14.3 ± 1.0 м (4–26 м). Подлесок (сомкнутость менее 50%) образован низкими кустарниками (высотой 0.6–1.2 м). Часто встречаются смородина печальная (Ribes triste), шиповник иглистый (Rosa acicularis). В полидоминантном ТКЯ единственный константный вид – вейник пурпурный, его среднее покрытие 12%, оно варьирует от 1 до 25%, но он не является доминантом в сообществах серии. С покрытием до 20–30% встречаются недоспелка копьевидная, иван-чай узколистный (Chamerion angustifolium L.), пырейник смешиваемый (Elymus confuses), пырейник длинноколосый, хвощ луговой (Equisetum pretense), мятлик луговой (Poa pratensis L.), грушанка круглолистная (Pyrola rotundifolia L.). Мхи произрастают на валеже и приствольных повышениях с покрытием менее 1%. Общее число видов, встреченных на 49 ПП, – 138. Число видов на ПП варьирует от 4 до 43, среднее – 12. Наибольшее число видов (39 и 43) отмечено в чозенниках с обильной недоспелкой копьевидной, индицирующей наиболее богатые почвы лесных пойм в районах исследований. Здесь, как и в чозеннике вейниковом с недоспелкой копьевидной, наряду с эутрофными видами встречаются олиготрофные кустарнички, заселяющие отложения крупнопесчаного аллювия прошлых лет.
Серия ассоциаций Alnosa fruticosae – ольховниковая (8 ПП). Включает две ассоциации: Populetum suaveolentis calamagrostidoso-alnosum fruticosae – тополевник вейниково-ольховниковый и Chosenietum alnosum fruticosae – чозенник ольховниковый. Сообщества серии занимают более высокие уровни, чем сообщества серии Calamagrostidosa, они приурочены к пойменным террасам, расположенным на высоте 2.6 ± 0.01 м (2.5–2.7 м) над уровнем воды в межень. Почвы дерново-аллювиальные грунтово-глеевые сильно- и средне-скелетные. Высота ДЯ – 21 ± 0.7 м (20–24 м), возраст – 60–75 лет. Сомкнутость подлеска более 50%; в нем преобладает ольха кустарниковая (Alnus fruticosa) высотой 2.5–3.5 м; константны смородина печальная, шиповник иглистый. В ТКЯ доминирует вейник пурпурный, константна княженика обыкновенная (Rubus arcticus L.). Покрытие мхов менее 2%. Всего на 8 ПП встречено 37 видов. Число видов на ПП варьирует от 6 до 21, среднее – 12.
Серия ассоциаций Herboso-Hylocomiosa – травяно-зеленомошная (5 ПП). Представлена тремя ассоциациями: Salicetum schwerinii herboso-hylocomiosum – швериноивняк травяно-зеленомошный, Populetum suaveolentis herboso-hylocomiosum – тополевник травяно-зеленомошный, Chosenietum herboso-hylocomiosum – чозенник травяно-зеленомошный. Уровень террасы – 3.3 ± 0.3 м (3.2–3.6 м). Местообитания краткопоемные, слабо аллювиальные. Почва – подбур грубогумусированный на песчано-галечных аллювиальных отложениях, сильноскелетный. Описания выполнены в верховьях рек, на границе с горными тундрами и лиственничными редколесьями. Возраст древостоев – более 50 лет. Высота ивняков – 3–4 м, тополевников – до 15 м, чозенников – 7 м. В континентальных районах Колымского нагорья, верхнем и среднем течении р. Пенжины для сообществ серии характерно присутствие в древостое лиственницы Каяндера (Larix cajanderi). Подлесок редкий или средней сомкнутости (менее 20%), константны курильский чай (Potentilla fruticosa L.) и ива мохнатая (Salix lanata L.). ТКЯ злаково-разнотравный, константны вейник краснеющий (Calamagrostis purpurascens), мятлик арктический (Poa arctica), княженика обыкновенная, голубика обыкновенная; доминируют вейник пурпурный, волоснец аянский (Elymus ajanensis), грушанка мясо-красная (Pyrola incarnate). Характерен развитый МЛЯ, его среднее покрытие 43% (варьирует от 15 до 60%); константны бореальные и субарктические виды мхов: аулакомниум вздутый (Aulacomnium turgidum), плевроциум Шребера (Pleurozium schreberi), полия поникшая (Pohlia nutans), саниония крючковатая (Sanionia uncinata). В качестве доминанта отмечен плевроциум Шребера, иногда обилен гилокомиум блестящий (Hylocomium splendens). На 5 ПП отмечено 55 видов. Число видов на ПП варьирует от 10 до 26, среднее – 20 видов.
Серии ассоциаций пойменных лесов расположены в порядке уменьшения поемности и аллювиальности: 1 – редкотравная; 2 – вейниковая; 3 – кустарниково-травяная; 4 – ольховниковая; 5 – травяно-зеленомошная. В этом ряду наблюдается последовательное изменение структуры фитоценозов от редкотравных молодняков на свежих аллювиях к лесам вейниковой серии и далее к серии кустарниково-травяной и ольховниковой (рис. 2). В последнем звене этого ряда, по мере выхода сообществ из поемного режима, находятся наиболее сукцессионно-продвинутые леса травяно-зеленомошной серии с развитыми кустарниковым, травяным и моховым ярусами, нередко с участием лиственницы Каяндера или кедрового стланика (рис. 2). По составу и структуре они приближаются к северотаежным белоберезовым и лиственничным лесам Северо-Востока, а в Берингийской лесотундровой области – к ерниковым тундрам и кедровым стланикам.
Рис. 2.
Изменение ценотической структуры пойменных лесов в ряду уменьшения поемности и аллювиальности, отрезки с точкой – стандартная ошибка. Условные обозначения: ярусы: ДЯ – древесный, КЯ – кустарниковый, ТЯ – травяно-кустарничковый, МЯ – мохово-лишайниковый; серии ассоциаций: РДТ – редкотравная; В – вейниковая; КТР – кустарниково-травяная; ОЛ – ольховниковая; МХ – травяно-зеленомошная.
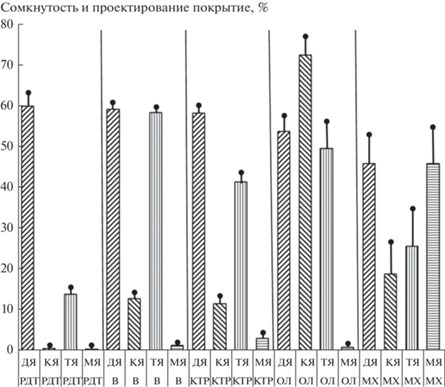
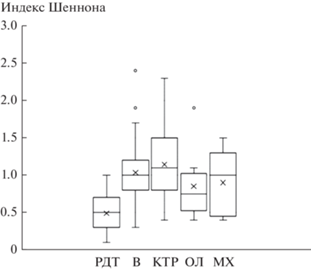
В рассматриваемом эколого-динамическом ряду сообществ происходит увеличение видового богатства и разнообразия от редкотравной серии к травяно-зеленомошной, что проявляется в увеличении числа видов во всех ярусах (рис. 3, табл. 1) и в увеличении значения индекса Шеннона (рис. 4, табл. 1). При этом максимальное разнообразие ТКЯ наблюдается в кустарниково-травяной серии; а в более продвинутых в сукцессионном ряду ольховниковой и травяно-зеленомошной сериях оно ниже (рис. 5). Таким образом, увеличение разнообразия идет за счет формирования мохового и кустарникового ярусов (рис. 2) и увеличения их видового богатства (табл. 1).
Рис. 3.
Изменение числа видов в сообществах пойменных лесов в ряду уменьшения поемности и аллювиальности; обозначения: крестик посередине прямоугольника – среднее арифметическое, линия чуть выше или ниже крестика – медиана, нижняя и верхняя грани прямоугольника соответствуют первому и третьему квартилю, расстояние между 1-м и 3-м квартилем – межквартильный размах, горизонтальные черточки на конце “усов” – максимальное и минимальное значения (без учета выбросов), отдельные точки – выбросы, т.е. значения, выходящие за пределы 1.5 межквартильных значений размаха от ближайшего квартиля; обозначения серий ассоциаций те же, что на рис. 2.
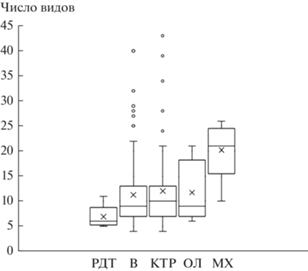
Таблица 1.
Показатели разнообразия пойменных лесов севера Корякского округа и северо-востока Магаданской области
| Показатели | Серия ассоциаций | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Oligo-herbosa | Calamagros-tidosa | Fruticoso-Herbosa | Calamagrostidoso-Alnosa | Herboso-Hylocomiosa | ||||||
| Количество ПП | ||||||||||
| 20 | 95 | 49 | 8 | 5 | ||||||
| M | SE | M | SE | M | SE | M | SE | M | SE | |
| Сомкнутость древесного яруса, % | 60 | 3.4 | 59.2 | 1.6 | 58 | 2 | 54 | 4 | 46 | 7.2 |
| Количество видов ДЯ | 2.9 | 0.2 | 2.5 | 0.1 | 2.5 | 0.1 | 2.5 | 0.3 | 2.2 | 0.4 |
| Индекс Шеннона ДЯ | 1.1 | 0.1 | 0.9 | 0.01 | 0.9 | 0.05 | 0.9 | 0.1 | 0.7 | 0.2 |
| Индекс Пилу ДЯ | 0.8 | 0.01 | 0.8 | 0.01 | 0.7 | 0.03 | 0.7 | 0.1 | 0.6 | 0.1 |
| Сумма сомкнутости крон подлеска, % | 0.7 | 0.5 | 12.9 | 1.4 | 12 | 2 | 73 | 4.4 | 19 | 8.2 |
| Количество видов подлеска | 0.7 | 0.2 | 2.2 | 0.2 | 2.2 | 0.3 | 3.5 | 0.4 | 3.8 | 0.9 |
| Индекс Шеннона для подлеска | 0 | 0.01 | 0.4 | 0.01 | 0.3 | 0.1 | 0.8 | 0.1 | 0.4 | 0.2 |
| Индекс Пилу для подлеска | 0.3 | 0.1 | 0.5 | 0.01 | 0.4 | 0.1 | 0.5 | 0.1 | 0.2 | 0.2 |
| Сумма покрытий видов ТЯ, % | 14 | 1.6 | 58.5 | 1.6 | 42 | 2.5 | 50 | 6.9 | 26 | 8.9 |
| Количество видов ТЯ | 3.2 | 0.3 | 5.4 | 0.4 | 6.2 | 0.8 | 4.5 | 1.3 | 8.6 | 2.4 |
| Индекс Шеннона ТЯ | 0.5 | 0.01 | 1 | 0.01 | 1.0 | 0.1 | 0.9 | 0.2 | 0.9 | 0.2 |
| Индекс Пилу ТЯ | 0.4 | 0.1 | 0.6 | 0.01 | 0.6 | 0.03 | 0.6 | 0.1 | 0.3 | 0.1 |
| Сумма покрытий видов МЯ, % | 0.5 | 0.5 | 1.4 | 0.5 | 3.1 | 1.2 | 0.9 | 0.5 | 46 | 7.1 |
| Количество видов МЯ | 0.2 | 0.1 | 1.2 | 0.2 | 1.2 | 0.3 | 1.3 | 0.6 | 5.6 | 1.1 |
| Индекс Шеннона МЯ | 0 | 0.01 | 0.1 | 0.01 | 0.1 | 0.04 | 0.1 | 0.01 | 1.2 | 0.2 |
| Индекс Пилу МЯ | 0 | 0.01 | 0.1 | 0.01 | 0.1 | 0.03 | 0 | 0.01 | 0.5 | 0.1 |
| Количество видов всех ярусов | 6.9 | 0.4 | 11.3 | 0.7 | 12 | 1.2 | 12 | 2.1 | 20 | 3.8 |
| Сумма индексов Шеннона | 1.6 | 0.1 | 2.4 | 0.1 | 2.5 | 0.1 | 2.6 | 0.2 | 3.2 | 0.4 |
| Средний индекс Пилу по всем ярусам | 0.4 | 0.01 | 0.5 | 0.01 | 0.5 | 0.02 | 0.5 | 0.1 | 0.4 | 0.1 |
| Уровень террасы | 1.6 | 0.1 | 2.1 | 0.1 | 1.9 | 0.01 | 2.6 | 0.01 | 3.3 | 0.3 |
Рис. 4.
Изменение суммы индексов Шеннона по всем ярусам в ряду уменьшения поемности и аллювиальности; обозначения те же, что на рис. 2, 3.
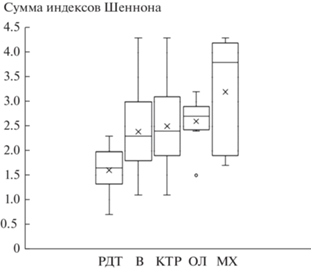
Рис. 5.
Изменение индекса Шеннона для ТКЯ в ряду уменьшения поемности и аллювиальности; обозначения серий ассоциаций те же, что на рис. 2, 3.
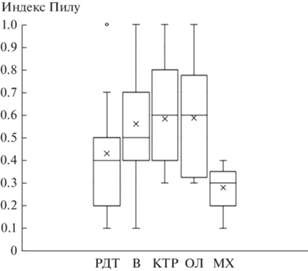
Индекс Пиелу (Е), рассчитанный как средний по ярусам, незначительно меняется в ходе аллювиальной сукцессии, принимая значения от 0.4 до 0.5 (табл. 1). В то же время неожиданным оказалось низкое значение этого индекса для ТКЯ травяно-моховой серии, по сравнению с индексами, рассчитанными для ТКЯ других серий (рис. 6). Это можно объяснить, во-первых, невысоким суммарным покрытием видов ТКЯ и, во-вторых, наличием в нем хорошо выраженных доминантов.
Рис. 6.
Изменение индекса Пиелу для ТКЯ пойменных лесов в ряду уменьшения поемности и аллювиальности; обозначения те же, что на рис. 2, 3.
ЗАКЛЮЧЕНИЕ
Пойменные леса севера Корякского округа, северо-востока Магаданской обл. и юга Чукотского АО находятся близ северного предела распространения. Несмотря на сравнительно невысокое видовое богатство, они увеличивают общее разнообразие растительного покрова Северо-Востока РФ. На основе табличного анализа 177 геоботанических описаний приведена геоботаническая характеристика пойменных лесов, образованных чозенией, тополем душистым, ольхой пушистой, ивами удской и Шверина, лиственницей Каяндера, распространенных в долинах рек севера Корякского округа, северо-востока Магаданской области и юга Чукотского АО. На пробных площадях всего отмечено 205 видов растений, в том числе сосудистых 166 видов, мохообразных – 36, лишайников – 3. Сообщества пойменных лесов отнесены к шести формациям и 19 ассоциациям, объединенным в 5 серий. Серии расположены в эколого-динамическом ряду по степени ослабления поемного и аллювиального режимов: редкотравная (3 ассоциции) → вейниковая (6 ассоциаций) → кустарников-травяная (5 ассоциаций) → ольховниковая (2 ассоциации) → → травяно-зеленомошная (3 ассоциации).
Ивняки из древовидных ив удской и Шверина, чозенники и тополевники встречаются в поймах рек на всей территории исследований. Ивняки образованы пионерными древесными породами, встречаются на низких участках прирусловой поймы, подвержены регулярному длительному затоплению во время паводков. Чозенники приурочены к молодым галечникам, перекрытым тонким слоем песчаного аллювия; почвы под ними примитивные, маломощные. Чозениевые древостои с участием тополя душистого (до 2 единиц) приурочены к участкам пойм, подверженным кратковременному затоплению. Сукцессионная динамика идет по пути постепенного отмирания чозении и увеличения доли тополя в древостое. Тополевники встречаются на участках высокой поймы, вышедших из-под влияния регулярного затопления. Для них характерны аллювиальные дерновые почвы и дерново-подбуры (мощность почвенного профиля 35–55 см) на галечниках или песчано-галечных аллювиальных отложениях. По мере выхода сообществ из поемного режима формируются леса травяно-зеленомошной серии с развитыми кустарниковым, травяным и моховым ярусами; в континентальных районах с участием лиственницы, в приморских – кедрового стланика. Выражена меридиональная дифференциация пойменных лесов: пойменные лиственничники, а также тополевые, чозениевые и ивовые леса с участием в древостое лиственницы Каяндера распространены в континентальных районах Магаданской обл. и юга Чукотского АО, а на севере Корякского округа встречаются только в бассейне р. Пенжины, в ее верхнем течении. Ольховые леса из ольхи пушистой отсутствуют в континентальных районах севера Корякии и Магаданской обл., встречаясь лишь на Охотском побережье и в восточной части Корякского округа, испытывающей влияние Берингова моря. Ольховые леса приурочены к притеррасной пойме с мелкодисперсным аллювием.
Вычислены показатели видового разнообразия и выравненности для каждого из изученных сообществ. Наименьшим значением индекса Шеннона характеризуется пионерный чозенник редкотравный, встречающийся на молодых галечниках; а наибольшим значением этого индекса – тополевник травяно-зеленомошный, занимающий наиболее высокие уровни поймы. Остальные ассоциации показали промежуточные значения индекса Шеннона. Выявлена тенденция к увеличению показателей видового разнообразия в ходе аллювиальной сукцессии. Полученные данные позволяют предположить, что при уменьшении поемности и аллювиальности увеличивается видовое разнообразие сообществ.
Сообщества пойменных лесов имеют большое водоохранное и противоэрозионное значение, закрепляя берега нерестовых рек, создавая необходимые условия для поддержания численности лососевых рыб. Пойменные леса используются местным населением как источник древесины и недревесного сырья и нуждаются в охране.
Список литературы
Ахтямов М.Х. Ценотаксономия прирусловых ивовых, ивово-тополевых и уремных лесов поймы реки Амур. Владивосток: Дальнаука, 2001. 138 с.
Балмасова М.А., Нешатаева В.Ю. Пойменные леса // Растительность Кроноцкого государственного заповедника (Восточная Камчатка). Тр. Ботанического института им. В.Л. Комарова РАН. 1994. Вып. 16. С. 77–80.
Биркенгоф А.Л. Леса центральной части полуострова Камчатки // Тр. СОПС АН СССР. Сер. Камчатская. М.; Л.: Изд-во Акад. наук СССР, 1938. № 6. 220 с.
Бурцева Е.И., Дестякина Л.И. Геоботаническая карта долины р. Неры // Проблемы ботаники на рубеже ХХ–ХХI вв. Тез. докл., представл. II(Х) съезду РБО. Т. 1. СПб., 1998. С. 231—232.
Василевич В.И. Видовое разнообразие в еловых лесах Европейской России // Ботанический журнал. 2015. Т. 100. № 12. С. 1249–1259. https://doi.org/10.1134/S0006813615120017
Василевич В.И. Видовое разнообразие растительности // Сибирский экологический журнал. 2009. Т. 16. № 4. С. 509–517.
Василевич В.И. Видовое разнообразие травяного яруса широколиственных лесов Северо-Запада Европейской России // Ботанический журнал. 2018. Т. 103. № 8. С. 955–967.
Василевич В.И. Видовое разнообразие сообществ черноольховых лесов Северо-Запада Европейской России // Ботанический журнал. 2017. Т. 102. № 7. С. 889–900. https://doi.org/10.1134/S000681361707002X
Василевич В.И., Кессель Д.С. Видовое разнообразие сообществ березовых и сероольховых лесов Северо-Запада России // Ботанический журнал. 2017. Т. 102. № 5. С. 585–597. https://doi.org/10.1134/S0006813617050015
Васильев В.Н. Растительность Анадырского края. М.; Л.: Изд-во Акад. наук СССР, 1956. 218 с.
Воробьев Д.П. Растительность южной части побережья Охотского моря // Тр. Дальневост. фил. АН СССР. Сер. Ботан. 1937. Т. 2. С. 19–102.
Гаращенко А.В. Флора и растительность Верхнечарской котловины. Новосибирск: Наука, 1993. 280 с.
Гельтман В.С. Географический и типологический анализ лесной растительности Белоруссии. Минск: Наука и техника, 1982. 326 с.
ГОСТ 18486-87. Лесоводство. Термины и определения. Утвержден и введен в действие Постановлением Государственного Комитета СССР по стандартам от 10 декабря 1987 г. № 4445.
Зархина Е.С. Тополевые леса // Леса Дальнего Востока. М.: Лесная промышленность, 1969а. С. 188–196.
Зархина Е.С. Чозениевые леса // Леса Дальнего Востока. М.: Лесная промышленность, 1969б. С. 196–198.
Зархина Е.С. Фациальная структура дальневосточных тополевников // Проблемы рационального лесопользования на Дальнем Востоке. Хабаровск: ДальНИИЛХ, 1986. С. 31–40.
Исаев А.П., Кузнецова Л.В. Растительность // Биоразнообразие ландшафтов Токинской котловины и хребта Токинский Становик. Новосибирск: Изд-во СО РАН, 2010. С. 142–189.
Кабанов Н.Е. Лесная растительность Советского Сахалина. Владивосток: Горно-таежная станция АН СССР, 1940.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Клюкин Н.К. Климат // Север Дальнего Востока. Москва: Наука, 1970. С. 101–132.
Колесников Б.П. Чозения и ее ценозы на Дальнем Востоке // Тр. Дальневост. фил. АН СССР. Сер. Ботан. 1937. Т. 2. С. 703–800.
Кондратюк В.И. Климат Камчатки. М.: Гидрометеоиздат, 1974. 204 с.
Котляров И.И. Краткая характеристика лиственничников юга Магаданской области // Биологические проблемы Севера. Вып. 42. Магадан, 1971. С. 188–197.
Куваев В.Б. Растительность Восточного Верхоянья // Растительность Крайнего Севера и ее освоение. 1956. Т. 2. С. 132–186.
Левич А.П. Структура экологических сообществ. М.: МГУ, 1980. 181 с.
Лесоустроительная инструкция. Утверждена приказом Минприроды России от 29 марта 2018 г. № 122. 42 с.
Москалюк Т.А. Структура и продуктивность лесов Северного Охотоморья. Владивосток: ДВО АН СССР, 1988. 144 с.
Нешатаев В.Ю. Проект Всероссийского кодекса фитоценологической номенклатуры // Растительность России. 2001. № 1. С. 62–70.
Нешатаев В.Ю., Потокин А.Ф., Томаева И.Ф. Добрыш А.А., Чернядьева И.В., Потемкин А.Д., Егоров А.А. Растительность, флора и почвы Верхне-Тазовского государственного заповедника. СПб, 2002. 164 с.
Нешатаева В.Ю. Растительность полуострова Камчатка. М.: Товарищество научных изданий КМК, 2009. 537 с.
Нешатаева В.Ю., Нешатаев В.Ю., Кириченко В.Е. Растительный покров территории Северной Корякии (Камчатский край) и ее геоботаническое районирование // Вестник СПбУ. Науки о Земле. 2020. Т. 65. № 2. С. 1–32. https://doi.org/10.21638/spbu07.2020.210
Нешатаева В.Ю., Нешатаев В.Ю., Кораблев А.П., Катютин П.Н. Пойменные леса Пенжинского района Камчатского края // Ботанический журнал. 2018. Т. 103. № 10. С. 1212–1239.
Нешатаева В.Ю., Нешатаев В.Ю., Откидач М.С. Пойменные леса Парапольского участка Корякского заповедника (Пенжинский р-н Камчатского края) // Тр. Кроноцкого государственного природного биосферного заповедника. 2017. Вып. 5. С. 82–95.
Осипов С.В. Растительный покров таежно-гольцовых ландшафтов Буреинского нагорья. Владивосток: Дальнаука, 2002. 378 с.
Полежаев А.Н. Растительный покров побережья Тауйской губы // Биологическое разнообразие Тауйской губы Охотского моря. Владивосток: Дальнаука, 2005. С. 644–666.
Розенберг Г.С. Информационный индекс и разнообразие: Больцман, Котельников, Шеннон, Уивер // Самарская Лука. 2010. Т. 19. № 2. С. 4–25.
Синельникова Н.В. Эколого-флористическая классификация пойменных лесов Магаданской области // Сибирский экологический журнал. 1995. № 4. С. 383–389.
Синельникова Н.В. Таежные лиственничные леса союза Roso acicularis-Laricion cajanderi all. nov. на северо-востоке России // Растительность России. 2016. № 28. С. 125–138.
Сосудистые растения советского Дальнего Востока / Под ред. С.С. Харкевича. Л.; СПб.: Наука, 1985–1996. Т. 1–8.
Сукачев В.Н. Типы лесов и типы лесорастительных условий. M.: Гослестехиздат, 1945. 37 с.
Сукачев В.Н. Основные принципы лесной типологии // Тр. Совещания по лесной типологии. М.: Изд-во АН СССР, 1951. С. 7–19.
Тихомиров Б.А. Краткий очерк долинной растительности Пенжинского района // Тр. Дальневост. фил. АН СССР. 1935. Т. 1. С. 85–112.
Тюлина Л.Н. Лесная растительность среднего и нижнего течения р. Юдомы и низовьев р. Маи. М.: Изд-во АН СССР, 1959. 222 с.
Тюлина Л.Н. Растительность западного побережья Камчатки // Тр. КИЭП ДВО РАН. Вып. 2. Петропавловск-Камчатский, 2001. 302 с.
Шелудякова В.А. Чозения в Якутской АССР // Ботанический журнал. 1943. Т. 28. № 1. С. 30–34.
Яровой М.И. Растительность бассейна р. Яны и Верхоянского хребта // Советская ботаника. 1939. № 1. С. 21–40.
Andreev M.P., Kotlov Yu.V., Makarova I.I. Checklist of lichens and lichenicocous fungi of the Russian Arctic // The Bryologist. 1996. V. 99. № 2. P. 137–169.
Ignatov M.S., Afonina O.M., Ignatova E.A. et al. Check-list of mosses of East Europe and North Asia // Arctoa. 2006. № 15. P. 1–130. https://doi.org/10.15298/arctoa.15.01
Krestov P.V. Forest Vegetation of Easternmost Russia (Russian Far East). Forest vegetation of Northeast Asia. Kluwer Academic Publishers. Dordrecht, 2003. P. 93–180.
Miyawaki A. A general survey of japanese vegetation // Veroff. Geobot. Inst. ETH. Stiftung Rubel. Zurich, 1988. № 98. P. 74–99.
Pielou E.C. Ecological Diversity. N.Y.: Gordon & Breach Sci. Publ., 1975. 165 p.
Qian H., Krestov P.V., Fu P., Wang Q., Song J.-S., Chourmouzis C. Phytogeography of the Far East Asia. Forest vegetation of Northeast Asia. Kluwer Academic Publishers, Dordrecht. 2003. P. 51–91.
Дополнительные материалы отсутствуют.