

Лесоведение, 2022, № 6, стр. 687-702
Биоразнообразие и география горных бореальных лесов Северного Забайкалья
Г. Н. Огуреева a, *, М. В. Бочарников a, А. А. Виноградов a
a Московский государственный университет имени М.В. Ломоносова
119991 Москва, Ленинские горы, д. 1, Россия
* E-mail: ogur02@yandex.ru
Поступила в редакцию 20.01.2022
После доработки 19.03.2022
Принята к публикации 07.06.2022
- EDN: LLWLCD
- DOI: 10.31857/S0024114822060080
Аннотация
В биогеографии успешно развивается направление географии биоразнообразия, с которым связаны многие аспекты дифференциации биоты на разных уровнях организации биотического покрова. Исследование географии биоразнообразия в данной работе опирается на базовую классификацию наземных экосистем, в рамках которой используется эколого-географический подход к интерпретации их биоразнообразия. Приводится обоснование региональных оробиомов как опорных единиц оценки видового и экосистемного разнообразия гор. Цель настоящего исследования – выявить региональную специфику типологического разнообразия горных лесных экосистем и биоты в связи с высотно-поясной структурой и высотными градиентами, определить их роль в организации горного пространства в связи с его экотопической структурой. Для достижения цели использованы статистические методы обработки цифровых картографических моделей растительности, рельефа, а также климатических условий. На примере Северовосточно-Забайкальского таежного оробиома раскрыта высотно-поясная структура его растительного покрова. На основе составленной мелкомасштабной карты лесов оробиома выявлена пространственная структура горнотаежного пояса с отображением его типологического разнообразия. Горнотаежный пояс (с двумя подпоясами) определяет региональную специфику флористического и ценотического разнообразия восточносибирских горных лесов оробиома. В составе лесного покрова пояса преобладают лиственничные леса, ограниченное участие принимают сосновые и темнохвойные леса, отмечается снижение видового состава по высотному градиенту. Наряду с общими для оробиома чертами выражены специфические особенности биоразнообразия лесов в его географических вариантах, связанные с различным соотношением типологических подразделений на разных высотных уровнях. Для горнотаежного пояса определены ключевые показатели тепло- и влагообеспеченности высотных поясов, характеризующие климатические условия формирования биоразнообразия в горах Северного Забайкалья. Успешное обращение к оробиомам при анализе биоразнообразия горных территорий определяет перспективы изучения лесов, разработки системы их мониторинга и охраны на единой биомной основе.
Разноплановое изучение географии биоразнообразия и развитие подходов к его сохранению – одна из основных проблем биогеографии и природоохранной деятельности. Обращение к концепции биомной организации биосферы возникло в связи с решением задач, вызванных глобальными процессами, в первую очередь по изменению климата. Биомы вошли в практику биогеографических исследований как определенный уровень структуры биотического покрова, а сам биом рассматривается как закономерное сочетание экосистем в географическом пространстве в определенных климатических условиях с адаптированной к ним биотой и сложившейся структурой (Вальтер, 1968; Walter,1973; Уиттекер, 1980). При классификации наземных экосистем различаются зональные биомы в равнинных условиях, биомы гор (оробиомы) и экосистемы крупных эдафических вариантов биома (педобиомы) (Walter, Breckle, 1991). На глобальном уровне выделяются зонобиомы и оробиомы I порядка, на региональном уровне – региональные биомы и оробиомы II порядка и экосистемы топологического плана (до биогеоценоза) с опорой, прежде всего, на климатические типы растительности (Köppen, 1936; Holdridge, 1947; Trewartha, Horn, 1980), жизненные формы биоты (Вальтер, 1968; Одум, 1986), адаптированные к соответствующим условиям и структурно-динамическим особенностям самих экосистем. Целостность экосистемы (специфический для нее инвариант) определяется разными масштабами современных и исторических процессов, определяющих состав, свойства, характер функционирования компонентов экосистем и специфику их отклика на меняющиеся условия среды (Сочава, 1978). Биота в этом плане является наиболее мобильным компонентом экосистем и быстро реагирует на изменения условий существования. Именно состав биоты и характер ее взаимодействия со средой определяют сохранение инварианта экосистем. Понимание общих принципов формирования экосистемного разнообразия на основе анализа состояния биоты позволяет выйти на разработку адресной системы мероприятий по нормализации экологического потенциала биома, восстановлению его экологических функций, выработке рекомендаций по организации управления природными ресурсами, что крайне важно для эффективного решения практических задач мониторинга и сохранения биоразнообразия. Особенно актуальны эти проблемы в лесоведении и лесной экологии, где определяются концепции управления лесами и принципы лесопользования.
Среди категорий экосистемного разнообразия наименее разработанным остается представление о классификации горных экосистем, их объеме и структуре, составе и условиях развития. География разнообразия горных экосистем формируется под воздействием интегрального высотного градиента в соответствии с трехмерной природой гор. Высотно-поясное распределение биотического покрова, определяемое изменением климатических параметров по градиентам теплообеспеченности в горах, является базовым представлением для классификации экосистем на всех уровнях биомного разнообразия гор. Типы поясности и их классификация (Огуреева, 1999) дают основание для выделения оробиомов I порядка, высотно-поясные спектры необходимы для выделения региональных оробиомов, определения структуры экосистемного и видового разнообразия гор (Огуреева, Бочарников, 2017). Биотическое и ценотическое разнообразие горных лесов отражает совокупность климатических условий, сложность орографии, историю развития природного комплекса гор в историческое время.
Эколого-географический подход к определению экосистемного разнообразия лесов с учетом региональных особенностей формирования растительного покрова хорошо согласуется с положениями лесной биогеоценологии и вносит свой вклад в развитие учения о лесе (Морозов, 1949; Сукачев, 1972).
Леса России составляют около 22% покрытой лесом площади планеты (Состояние …, 2020). По официальным данным Государственного лесного реестра (на 2015 г.), площадь российских лесов составляла 1183.2 млн. га (Доклад …, 2015). В работе С.А. Барталева с соавторами (Барталев и др., 2016) приводится лесопокрытая площадь по данным дистанционного зондирования земли из космоса – 725.5 млн. га (без учета площади редин и кустарников). Биом бореальных лесов занимает широкую полосу (более 1000 км) на севере континентов Евразии и Северной Америки, образуя единую циркумбореальную зону, занимающую 14.5 млн. кв. км, что составляет 27% площади лесного покрова планеты; из них на долю России приходится 73% (FAO, 2020). Горы занимают в России порядка 430 млн. га. Площади горных бореальных лесов определить пока трудно, по предварительным данным, она может составлять 140 млн. га. Лесистость горных биомов варьирует в пределах 40–80%. Горные леса выполняют важнейшие экологические средообразующие и средозащитные функции, поддерживая устойчивость горных экосистем в условиях природных и антропогенных нарушений.
Границы обширного зонобиома бореальных лесов определяются биоклиматическими показателями, среди которых ведущими являются показатели теплообеспеченности (средняя многолетняя годовая температура воздуха – от –12 до +4°С; сумма активных температур >10°С – 1000–2000; продолжительность вегетационного периода – 100–220 дней), влагообеспеченности (среднее многолетнее годовое количество осадков – 300–1500 мм; режим их сезонного распределения), а также их соотношение в пределах региональных биомов. Бореальные леса Северного Забайкалья произрастают в условиях холодного и влажного резко континентального климата.
Экосистемное разнообразие бореальных лесов отражает различия климатических условий и в целом высокое ландшафтное разнообразие территории. В пределах зонобиома бореальных лесов России выделено 32 региональных биома, в том числе 16 равнинных (Биоразнообразие …, 2020) и 16 горных биомов (Карта …, 2018). В характеристику оробиомов включены сведения о биоклиматической ситуации (климатопы), в которой представлены высотно-поясные спектры, их высотные пределы и дифференциация по высотному градиенту экосистем, представлены географо-генетические комплексы формаций и количественная оценка биоты. Биоразнообразие бореальных лесов России формируется на основе трех географо-генетических комплексов формаций: урало-южносибирских (темнохвойных), восточносибирских, или ангаридских (светлохвойных), и южноохотских (темнохвойных) (Сочава, 1978).
Цель настоящего исследования – выявить региональную специфику типологического разнообразия горных лесных экосистем и биоты в связи с высотно-поясной структурой и высотными градиентами, определить их роль в организации горного пространства исходя из его экотопической структуры на примере Северовосточно-Забайкальского таежного оробиома. Фоновое участие в высотном спектре принимают горнотаежные экосистемы ангаридского и урало-южносибирского комплекса формаций, образующие пояс на абсолютных высотах около 300–1000 м. Устойчивое развитие горнотаежных экосистем может нарушиться в связи с глобальными трендами в изменении климата, под воздействием которых прогнозируются существенные изменения в структуре биоразнообразия, а также с интенсификацией освоения природных ресурсов, особенно лесопользования, в зоне Байкало-Амурской магистрали.
ОБЪЕКТЫ И МЕТОДИКА
В пределах бореальной области Сибири горнотаежные типы поясности с определяющей ролью хвойных сибирских лесов представлены наиболее разнообразно. В горах Байкало-Джугджурской природной области выражен трехчленный высотно-поясной ряд: горнотаежный пояс лиственничных лесов – пояс редколесий и кедрового стланика (Pinus pumila (Pall.) Regel) – гольцово-тундровый пояс, где в горнотаежном поясе преобладают лиственничные леса ангаридского комплекса формаций из лиственницы Гмелина (Larix gmelinii (Rupr.) Rupr.) (Белов, 1973; Панарин и др., 1980; Огуреева, 1999). Фитоценотическое и флористическое разнообразие бореальных лесов горнотаежного пояса оробиома послужило объектом исследования.
Северовосточно-Забайкальский региональный оробиом относится к Забайкальской группе горнотаежных биомов бореального класса (оробиомов I порядка). Он занимает обширные орографически сложные системы Северо-Байкальского, Станового и Патомского нагорий с преобладающими высотами хребтов 2000–2500 м.
С Северо-Байкальским нагорьем и западной частью Станового нагорья связан Северобайкальский вариант биома. На современный облик рельефа оказывает влияние древнее и современное горно-долинное оледенение, что определяет альпинотипный резко расчлененный рельеф в высокогорных областях Верхнеангарского и Северо-Муйского хребтов, открытых с северо-запада преобладающим воздушным массам. К зоне тектонических разломов приурочены межгорные котловины Байкальской рифтовой зоны (Верхнеангарская, Муйско-Куандинская). Патомское нагорье, с которым связано развитие Патомского варианта биома, имеет складчатое основание с хорошо выраженными участками древних поверхностей выравнивания; преобладающие высоты – 1200–1300 м. В условиях преобладания денудационных процессов и выхода пород различной устойчивости к денудации и выветриванию выражена ярусность рельефа. Кодаро-Каларский вариант биома развит на хребтах восточной части Станового нагорья. Открытый с северо-запада Кодарский хребет характеризуется альпинотипным рельефом, а на расположенных в более континентальных условиях с развитием криогенных процессов Южно-Муйском и Удоканском хребтах преобладают куполообразные вершины, обширные нагорные террасы. К Байкальской рифтовой зоне приурочен ряд межгорных котловин (крупнейшая из них –Чарская).
В целом биом формируется в условиях низкой тепло- и достаточной либо избыточной влагообеспеченности. Средние годовые температуры повсеместно отрицательные (от –3°С –в низкогорьях Патомского нагорья до –6°С – в среднегорьях и –9°С – в высокогорьях Кодарского, Каларского, Удоканского хребтов). Суммы активных температур выше 10°С изменяются от 0 до 1800, годовое количество осадков – от 300 до 600 мм. Северобайкальский и Патомский варианты характеризуются умеренно-холодным климатом (средняя многолетняя температура года – выше –6°C, июля – выше +16°C) с повышенным увлажнением, способствующим участию темнохвойных лесов в низкогорьях хребтов и господством в горнотаежном поясе лиственничных лесов (средняя многолетняя температура года составляет –6…–9°C, июля – +13…+15°C). Кодаро-Каларский вариант с хорошо развитыми высокогорными комплексами имеет наиболее холодный континентальный климат (средние годовые температуры опускаются до –10°С и ниже).
С изменениями в обеспеченности теплом, влагой и их соотношением связаны смены высотно-поясных подразделений растительности, поясов и подпоясов, которые выделяются в соответствии с дифференциацией господствующих типов растительности, фитоценотического оптимума фоновых сообществ в пределах вариантов биома. Высотные изменения в разнообразии горнотаежных экосистем, выраженные в типологическом разнообразии сообществ, составе ценофлор, пространственной структуре в системе экотопических условий, лесотаксационных показателях, находящие связь с климатическими градиентами, получили интегральное выражение в высотно-поясных комплексах лесов, аналогично выделенных в горах Южной Сибири (Поликарпов и др., 1986; Назимова и др., 2005). Инверсии температур в межгорных котловинах определяют их положение в структуре растительного покрова и всего высотно-поясного спектра в соответствии с высотным градиентом. География биоразнообразия и пространственная структура экосистем оробиома выражена также во флористическом составе преобладающих растительных формаций и высотных спектров (Бочарников, 2020).
Анализ биоразнообразия горнотаежных лесов Северовосточно-Забайкальского биома проводится на основе впервые созданной карты лесов, позволяющей провести сравнительную характеристику его трех географических вариантов. Составление карты лесов проведено на основе выявления факторных характеристик, наиболее точно описывающих пространственную структуру лесного покрова, и установления количественных связей между выбранными характеристиками и определенного на основе спектральных данных типологического разнообразия лесов. В качестве исходных данных об актуальном лесном покрове использована карта растительного покрова России (Барталев и др., 2015). Типологическое разнообразие лесов определено на основе оцифрованной карты растительности юга Восточной Сибири (Белов, 1973), публикаций (Курнаев, 1973; Панарин и др., 1980; Рысин, 2010) и авторских материалов, полученных при обследовании лесов в 2017–2019 гг. Всего выделено 12 фоновых лесных эпиформаций: 6 – лиственничных; 2 – сосновых; 2 – еловых; 1 – пихтово-кедровых; 1 – березовых лесов из шерстистой березы (Betula lanata (Regel) V. Vassil.). К производным лесам отнесены сообщества березовых (береза плосколистная (B. platyphtlla Sukaczev)) лесов. По долинам рек распространены условно коренные сообщества лесов с участием лиственницы, чозении, тополя, березы в составе серийных долинных рядов.
Создание инвентаризационной карты горнотаежных лесов проведено с использованием пошагового дискриминантного анализа и определением оптимального сочетания факторов, которые объясняют разграничение исходно известных типологических подразделений и возможность их формирования с разной вероятностью. Орографические характеристики (абсолютная высота, крутизна, экспозиция, кривизна склонов, относительное превышение, глубина заложения речных долин) использованы в качестве переменных, в соответствии с изменением которых выражена пространственная структура экологических факторов. Абсолютная высота использована для характеристики высотного градиента, в соответствии с которым в пределах горнотаежного пояса выражены высотные подпояса. Крутизна и кривизна склонов определяют различия в трофности местообитаний, экспозиция – теплообеспеченности в пределах подпоясов. Относительное превышение и глубина заложения речных долин служат дополнительными характеристиками перераспределения экотопических условий по элементам горного рельефа. В результате анализа определена роль каждой переменной и дана общая оценка качества выделения типологических подразделений лесов как интегральная характеристика.
На основе дискриминирующих функций неизвестные типологические подразделения отнесены к изначально известным по максимальным значениям вероятностей, рассматриваемых в качестве основного критерия при выборе соответствующего класса). Анализ сходства экотопических условий условно коренных лесов и развивающихся на их месте производных позволил выявить типологическое разнообразие березовых лесов как производных вариантов хвойных горнотаежных лесов.
С использованием алгоритма, реализованного при проведении дискриминантного анализа, получены три цифровые карты лесов (отдельно для Северобайкальского, Патомского и Кодаро-Каларского вариантов), что позволило составить карту Северовосточно-Забайкальского оробиома в целом. Картографическая реализация метода дискриминантного анализа для каждого варианта в отдельности обусловлена региональной спецификой формирования лесов в связи с орографической и ландшафтной структурой территории, что находит отражение в разнообразии типов поясности растительности с особенностями смены высотно-поясных подразделений на карте и их типологического состава. Качество дискриминации лесных формаций на основе выбранных морфометрических параметров варьирует от 51% (лиственничные леса Кодаро-Каларского варианта) до 96% (сосновые леса Кодаро-Каларского варианта).
Итоговая мелкомасштабная карта лесов оробиома получена в результате фильтрации растрового изображения с целью исключения единичных пикселей и образуемых ими мелких фрагментов в соответствии с возможностью отображения при выбранном масштабе картографирования (м. 1 : 3 000 000).
Карта лесов оробиома использована в анализе пространственной структуры фитоценотического разнообразия горнотаежного пояса. Абсолютная высота послужила ключевым критерием оценки высотно-поясной дифференциации типологических единиц бореальных лесов, позволившей на основе их распространения обосновать выделение высотных подпоясов и полос в пределах горнотаежного пояса. Оценка фитоценотического разнообразия проведена с учетом флористического разнообразия сосудистых растений горнотаежного пояса и видового состава основных ценофлор по опубликованным данным.
Ключевые биоклиматические показатели получены по данным глобальной климатической модели Chelsa (Karger et al., 2017). Она содержит информацию о пространственном распределении средних многолетних годовых и месячных показателей (стандартный набор 19 биоклиматических переменных BioClim) за период 1979–2013 гг., приведенных для пространственного разрешения в 30″. Использованы показатели средних многолетних годовых, июльских и январских температур и количества осадков.
На основе представлений о связях растительности с определенными климатопами, совокупность характеристик которых определяет функционирование биоты, для высотных поясов и подпоясов в пределах оробиома, рассчитаны количественные значения показателей тепло- и влагообеспеченности. Полученные совокупности значений позволили определить средние, стандартные отклонения от средних и экстремумы биоклиматических характеристик. Они использованы в качестве опорных данных для анализа современного разнообразия и пространственной структуры растительного покрова в системе климатических условий.
Пространственные операции с данными, в том числе сопряженный анализ растровых слоев, их векторизация, выполнены в геоинформационной среде программы ArcGIS (приложение ArcToolbox) и программы SAGA GIS. Статистический анализ выполнен в программе Statistica.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Горнотаежный пояс Северовосточно-Забайкальского оробиома распространен на высотной амплитуде от 300 до 1000 м и занимает площадь около 130 тыс. км2 (более 50% от площади биома). Каждый биом можно рассматривать как ценофонд или “фонд экосистем” разного уровня (Тишков, 1992). Оробиом характеризуется достаточно высокой степенью сохранности природных экосистем и развитием малонарушенных лесов, среди которых преобладают сообщества лиственничной формации (Атлас …, 2003). В структуре регионального биома, прежде всего, важно выделение климаксных сообществ, определяющих высотно-поясной характер биома и сопутствующих им эдафических или экологических экосистем. Специфика экосистем определяется составом биоты и характером ее взаимодействия со средой обитания в первую очередь с климатическими условиями.
Возможности развития растительного покрова горнотаежного пояса определены тепло- и влагообеспеченностью: значениями средней годовой температуры от –7 до –3°C, средней температуры июля от +14 до +19°C, суммами активных температур >10°С от 1000 до 1800, годовым количеством осадков от 320 до 440 мм (Бочарников, 2019). Определенные значения данных показателей в пределах их амплитуд характеризуют климатопы типологических подразделений бореальных лесов в границах горнотаежного пояса. Значительной амплитудой значений характеризуется нижний подпояс, представленный разнообразием типологического состава лиственничных, сосновых и темнохвойных лесов. Низкие средние годовые температуры (–7…–5°C) при достаточно высокой теплообеспеченности теплого сезона (сумма активных температур выше 10 °C более 1400, биологически активная температура более 4.5°C) определяют развитие лиственнично-сосновых лесов в нижних частях хребтов Станового нагорья и днищах межгорных котловин Байкальской рифтовой зоны. Пихтовые и еловые леса низкогорных хребтов Патомского нагорья произрастают при меньшей континентальности климата, характеризуяcь более высокими средними годовыми температурами (около –5…–3°C) при немного меньшей теплообеспеченности вегетационного сезона (сумма активных температур выше 10°C – 1200–1600). Верхний подпояс горнотаежного пояса, основу которого составляют лиственничные леса, характеризуется уменьшением теплообеспеченности (средняя годовая температура опускается до –9°C, сумма активных температур – до 1000). Важный ботанико-географический рубеж, связанный с верхней границей леса и приходящийся на высоты около 900–1200 м, определяется снижением теплообеспеченности вегетационного сезона (средняя температура июля опускается ниже +15°C, а сумма активных температур не превышает 1200). В целом высотная дифференциация лесов пояса надежно маркируется термическими показателями, а также соотношением тепла и влаги, прежде всего, теплого периода года.
Современная структура отражает также длительное историческое развитие, в ходе которого ангаридские, берингийские, южносибирские комплексы растительных формаций находят свое положение в структуре высотной поясности при разнообразии экотопических условий в пределах поясов. В обобщенном виде структура фитоценотического разнообразия горнотаежного пояса и отдельных его компонентов выглядит следующим образом. Преобладают леса восточносибирской формации лиственницы Гмелина (Larix gmelinii), доля которых составляет порядка 80% в составе лесного покрова биома (табл. 2). Типологический состав в целом представлен преобладающими кедровостланиковыми, рододендроновыми, ерниковыми, кустарничково-зеленомошными группами лиственничных лесов, которые определяют разнообразие горнотаежного пояса в условиях холодного континентального климата низкогорных и среднегорных хребтов Северного Забайкалья. Их широкое распространение в оптимальных условиях на высотах 600–900 м охватывает широкий спектр экотопических условий, связанных с развитием морфоструктур, литологией горных пород и почв, характеризуя выраженную климатическую обусловленность в пределах пояса. Лиственничные с редким участием сосны или кедра кедровостланиковые леса представлены баданово-зеленомошными, багульниково-зеленомошными, лишайниковыми сообществами. Бруснично-зеленомошный и голубично-зеленомошный напочвенный покров характерен для ерниковых елово-лиственничных лесов. В сообществах лиственничных кустарничково-зеленомошных лесов с высокой активностью брусники обыкновенной (Vaccinium vitis-idaea L.) не выражен подлесок.
Таблица 1.
Результаты дискриминантного анализа типологических подразделений лесов оробиома по морфометрическим показателям. Для типологических подразделений растительности приведены показатели F-статистики, Wilks’ Lambda, качества детерминации (%), число пикселей на карте (N). Уровень значимости (p-value): ** < 0.000; * < 0.05; – незначим при p < 0.05
| Географи-ческий вариант оробиома | N | F | Морфометрические показатели рельефа (Wilks’ Lambda) | Точность выделения групп, % | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| абсолютная высота | экспозиция склона | крутизна склона | относительное превышение | глубина заложения долин | ||||||
| Лесные формации и группы формаций | Лиственничные | 1 | 392 236 | 6147 | 0.85** | 0.74** | 0.75** | 0.76** | 0.76** | 49 |
| 2 | 507 642 | 5388.2 | 0.95** | 0.67– | 0.70* | 0.67– | 0.67– | 53 | ||
| 3 | 341 277 | 992.94 | 0.96** | 0.94** | 0.95** | 0.93** | 0.93** | 53 | ||
| Сосновые | 1 | 14 041 | 379.07 | 0.89** | 0.88* | 0.89** | 0.90** | 0.90** | 74 | |
| 2 | 46 202 | 78.101 | 1.00** | 1.00** | 1.00** | 1.00** | 1.00** | 98 | ||
| 3 | 2845 | 112.50 | 0.96** | 0.81** | 0.81** | 0.81** | 0.81** | 95 | ||
| Темнохвойные | 1 | 2406 | 125.52 | 0.72** | 0.64– | 0.63– | 0.67* | 0.63– | 89 | |
| 2 | 65 325 | 284.30 | 0.96** | 0.92** | 0.90** | 0.90** | 0.90** | 60 | ||
| 3 | 16 068 | 22.924 | 1.00** | 1.00** | 1.00** | 1.00** | 1.00** | 99 | ||
| Березовые | 1 | 40 996 | 437.47 | 0,71** | 0,61** | 0,64** | 0,64** | 0.66** | 36 | |
| 2 | 63 295 | 880.3 | 0.83** | 0.54– | 0.55– | 0.54– | 0.55– | 43 | ||
| 3 | 10 425 | 19.081 | 0.96** | 0.94** | 0.94** | 0.94** | 0.94** | 43 | ||
Таблица 2.
Распространение горнотаежных лесов Северовосточно-Забайкальского оробиома и его географических вариантов по занимаемой площади
| Типологические подразделения горнотаежных эпиформаций (% от лесопокрытой площади) | Лесопокрытая площадь (тыс. км2) | Лесистость (% от общей площади) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||||
| Оробиом | 39 | 8 | 17 | 19 | 1 | 0 | 4 | 3 | 2 | 5 | 2 | <1 | 101 | 44 | |
| Географические варианты | 1 | 38 | 11 | 9 | 18 | 3 | 0 | 5 | 5 | 2 | 5 | 4 | <1 | 41 | 50 |
| 2 | 35 | 10 | 8 | 28 | 0 | 0 | 7 | 0 | 2 | 8 | 2 | 0 | 33 | 48 | |
| 3 | 45 | 0 | 40 | 10 | 0 | 0 | 0 | 3 | 2 | 0 | 0 | <1 | 27 | 35 | |
Типологический состав лесных эпиформаций: лиственничной: 1 – кедровостланиковые; 2 – рододендроновые (рододендрон золотистый); 3 – ерниковые (береза тощая (Betula exilis), береза растопыренная (B. divaricata)); 4 – кустарничково-зеленомошные; 5 – рододендроновые (рододендрон даурский); 6 – разнотравные; сосновой: 7 – рододендроновые (рододендрон даурский); 8 – разнотравные; еловой: 9 – кедровостланиковые; 10 – кустарничково-зеленомошные; пихтово-кедровой: 11 – кустарничково-зеленомошные; березовой: 12 – травяные. Географические варианты оробиома: 1 – Северобайкальский; 2 – Патомский; 3 – Кодаро-Каларский.
Дополняют типологическое разнообразие пояса сосновые (7%) и темнохвойные (9%) леса урало-южносибирского комплекса формаций. Сосновые (сосна обыкновенная (Pinus sylvestris L.) леса характерны для нижнего, более теплообеспеченного подпояса, а также межгорных котловин, в которых при большей амплитуде годовых температур увеличена продолжительность и теплообеспеченность вегетационного сезона, а широкое развитие аллювиальных песчаных отложений препятствует развитию мерзлоты. Наиболее распространены сосновые с участием ели и пихты рододендроновые (рододендрон даурский (Rhododendron dauricum L.)) леса с хорошо развитым кустарничково-зеленомошным покровом. Темнохвойные леса имеют ограниченное распространение и представлены фрагментами пихтовых (пихтой сибирской (Abies sibirica)), кедровых (сосной сибирской (Pinus sibirica)) и еловых (елью сибирской (Picea obovata)) лесов, часто по долинам рек, достигая наибольшего распространения в низкогорьях Патомского и Северо-Байкальского нагорий в условиях менее континентального климата. Березовые (из березы плосколистной) леса являются типичной стадией восстановления светлохвойных и темнохвойных лесов после пожаров и рубок. Тополевые (тополь душистый (Populus suaveolens)), чозениевые (кореянка земляничниколистная (Chosenia arbutifolia), березовые (береза пушистая (Betula pubescens), береза плосколистная) и осиновые (осина обыкновенная (Populus tremula L.)) леса участвуют в составе серийных рядов по долинам рек, в днищах котловин. Кедровый стланик формирует подгольцовый пояс, на границе которого в контактной полосе образуются сложные сочетания его сообществ с таежными лесами и редколесьями, ценофлоры которых взаимно обогащаются. Восточносибирский горнотундровый комплекс является основой растительного покрова высокогорий биома. Современные климатические условия ограничивают его связи с горной тайгой, при этом проникновение горнотундровых видов осуществляется по благоприятным экотопам в пределах горнотаежного пояса (выходы горных пород, каменистые россыпи).
Оробиом относительно слабо изучен в ботанико-географическом плане, и о его истинном разнообразии можно судить пока лишь предварительно. Общее видовое богатство биома составляет около 1200 видов сосудистых растений (Малышев и др., 2000), 300 видов мохообразных и более 700 видов лишайников (Карта …, 2018). Наиболее разнообразна флора Северобайкальского варианта биома (около 1130 в.), около 720 видов зарегистрировано в Кодаро-Каларской части биома и около 630 видов – в Патомском варианте, наименее изученном в этом отношении. Наибольшее число видов приходится на флору горнотаежного пояса (около 1000 видов) (табл. 4). Три четверти этого разнообразия (около 840 видов) сосредоточено в нижнем подпоясе лиственнично-сосновых и лиственнично-еловых с участием пихты и кедра лесов. В верхнем подпоясе лиственничных лесов, преобладающем по занимаемой площади, насчитывается около 780 видов. Флористическое богатство подгольцового пояса составляет более 620 видов, при этом редколесный и стланиковый подпояса имеют примерно одинаковое разнообразие – по 550–560 видов; в горнотундровом поясе произрастает около 470 видов, в гольцовом зарегистрировано около 80 видов сосудистых растений (Иванова, Чепурнов, 1983). Число видов сосудистых растений на 100 км2 в горнотаежном поясе оценивается в 450–600 видов, в пересчете на 10 000 км2 – 650–800 видов (Малышев, 1994). В целом по высотному градиенту происходит уменьшение флористического разнообразия поясов, выраженное в сокращении общего числа видов и в уменьшении их видовой насыщенности.
Таблица 3.
Типологическое разнообразие горнотаежных лесов на разных высотных уровнях Северовосточно-Забайкальского оробиома и его географических вариантов (в % от общей лесопокрытой площади для каждой высотной ступени; показатель лесистости дан справа от диаграмм). Географические варианты оробиома: см. табл. 1
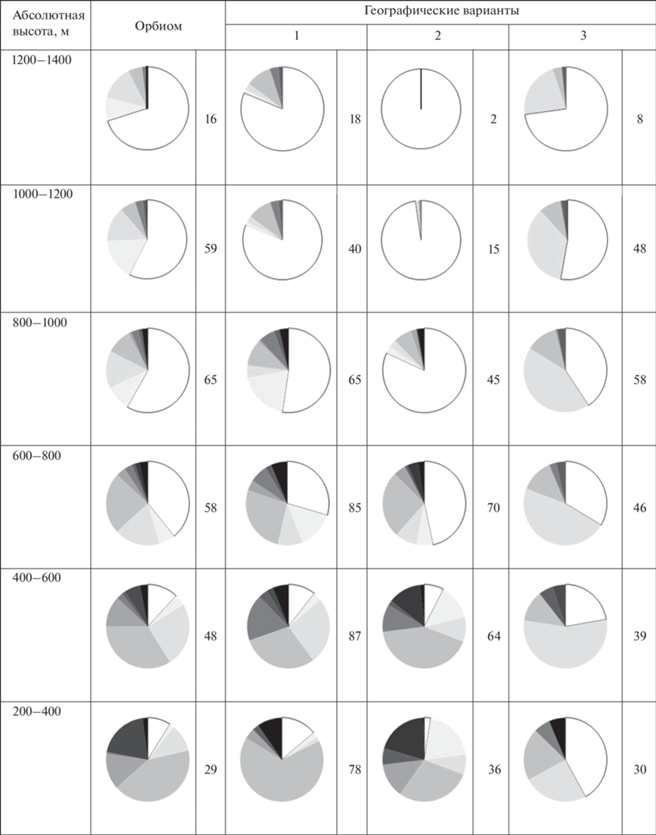
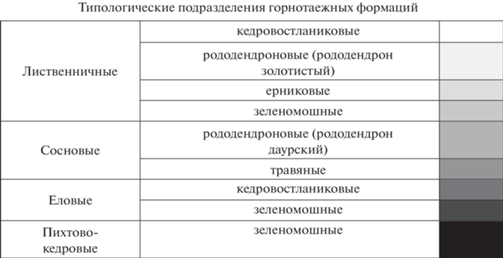
Таблица 4.
Видовое разнообразие сосудистых растений горнотаежного пояса оробиома и его географических вариантов: 1 – Северобайкальский; 2 – Патомский; 3 – Кодаро-Каларский
| Высотно-поясные подразделения | Оробиом | Географические варианты | ||||
|---|---|---|---|---|---|---|
| Высотный пояс | Высотные подпояса и ценофлоры | 1 | 2 | 3 | ||
| Горнотаежный | 1000 | 950 | 600 | 570 | ||
| Лиственнично-сосновых, елово-лиственничных и пихтово-кедровых лесов | 840 | 800 | 510 | 430 | ||
| Ценофлоры сосновых лесов | 630 | 610 | 380 | 340 | ||
| Лиственничных лесов | 780 | 750 | 530 | 520 | ||
| Ценофлоры лиственничных лесов | 620 | 590 | 410 | 430 | ||
| Подгольцовый | Лиственничных редколесий | 560 | 540 | 330 | 440 | |
Флора оробиома является бореальной. Ее основу составляют виды голарктической (25%), евро-азиатской (13%), евро-сибирской (5%), южносибирской (более 10%) ареалогических групп (Бочарников, 2019). При этом флора мало специфична, о чем свидетельствует относительно небольшое количество эндемичных (около 80) видов, а с учетом высокой роли видов с обширным ареалом, указывает на давние связи с флорами Голарктики. Большая их часть является региональными эндемиками и встречается, в основном в северобайкальской части биома, флора которого обогащена горностепными элементами, находящимися здесь на границе ареала. Степные эндемики (около 20 видов) сосредоточены в нижнем подпоясе горной тайги. Специфику флоры биома представляют и другие редкие виды растений. Развитие плейстоценовых реликтовых видов связано с горно-долинным оледенением хребтов в четвертичный период, в ходе которого активизировались процессы миграций видов между хребтами и их высотными поясами (всего более 20 видов). (Малышев, Пешкова, 1984). 14 видов сосудистых растений включены в Красную книгу РФ.
Фитоценотическое разнообразие и пространственная структура горнотаежных лесов биома
Горнотаежные леса составляют основу высотно-поясного спектра оробиома, при этом региональная специфика находит выражение в биоразнообразии его географических вариантов (рис. 1). Каждый вариант биома, отражая общие закономерности формирования разнообразия экосистем, имеет при этом свои специфические черты, определяемые географическим положением и связанным с ним характером экологических условий в ландшафтной структуре горной территории. При создании карты лесов для каждого варианта выбраны морфометрические показатели с высокой значимостью (табл. 1). Варианты горнотаежных лесов характеризуются различиями в типологических и структурных показателях разнообразия: в фитоценотическом составе формаций, богатстве их ценофлор, соотношении господствующих и сопутствующих им сообществ, высотных пределах их распространения.
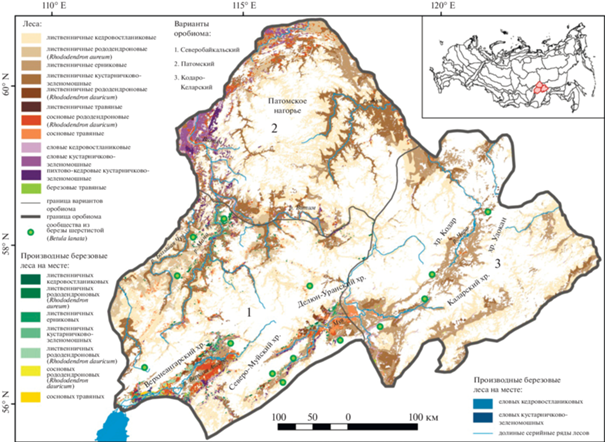
Северобайкальский вариант отличается наибольшим уровнем разнообразия экосистем в связи с тесным контактом с Байкальской котловиной и прилегающим к ней горным обрамлением рифтовой зоны как областей с высокой контрастностью и мозаичностью современных условий, выраженных в течение длительной истории развития и оставивших след, в том числе в виде реликтовых элементов биоты (Малышев, Пешкова, 1984; Намзалов, 2021). Горнотаежные и высокогорные (подгольцовые, горнотундровые) экосистемы занимают примерно равные площади горной территории, в пределах которой формируется ценотическое разнообразие варианта. В горнотаежном поясе преобладают лиственничные леса по всей амплитуде пояса (500–900 м). Они проникают по долинам рек в пределы подгольцового пояса, где представлены лиственничными с участием сосны и березы лесами с подлеском из золотистого рододендрона (Rhododendron aureum), приуроченные к увлажненной северо-западной части Северо-Байкальского нагорья в верховьях долин рек Верхней Чуи, Левой Мамы (табл. 2, рис. 2).
Рис. 2.
Высотная структура бореальных лесов Северовосточно-Забайкальского оробиома и его географических вариантов (приведены медианы, квартили 25 и 75%, максимальные и минимальные значения). Географические варианты оробиома: 1 – Северобайкальский; 2 – Патомский; 3 – Кодаро-Каларский. Типологические подразделения горнотаежных эпиформаций – см. табл. 1.
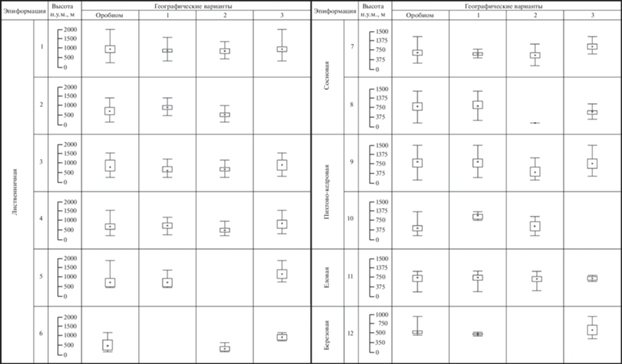
Фитоценотическое разнообразие горнотаежного пояса этого варианта имеет выраженную высотную дифференциацию, связанную с изменением соотношения типологических подразделений на различных высотных уровнях. Этим обосновывается не только выделение подпоясов сосновых и лиственничных лесов (нижний и верхний подпояса соответственно), но и полос в пределах подпоясов, которые отличаются характерными соотношениями фоновых и сопутствующих сообществ. Выбранный шаг в 200 м позволяет выявить различия в структуре фитоценотического разнообразия определенных формаций, которые обусловлены высотным градиентом (табл. 3). По всему высотному спектру господствующие позиции занимают лиственничные леса. При этом до 800 м выражено наибольшее типологическое разнообразие лесов, в котором сосновые леса занимают до 20% площади, а пихтово-кедровые и еловые – до 10%. Они произрастают также и на более высоких уровнях, однако там их доля не превышает 3%. Выше 800 м выражено заметное преобладание кедровостланиковых лиственничных лесов, на которые приходится около 1/2–3/4 доли площади.
Повышенный уровень разнообразия варианта выражен и в отношении видового богатства сообществ горнотаежного пояса (табл. 4). Оно связано, в том числе и с разнообразием ценофлор горнотаежных лесов, обогащенных южносибирскими элементами, которые типичны для темнохвойных лесов урало-южносибирского комплекса формаций, а также степными элементами, представленными как реликтовыми древнесредиземноморскими палеоэндемичными видами, так и неоэндемиками (Малышев, Пешкова, 1984). Флористический состав горнотаежного пояса обогащен также видами, характерными для сопутствующих формаций – пойменных лугов и низинных болот, широко представленных в Верхнеангарской котловине и на северном побережье Байкала. В связи с высокой мозаичностью экологических условий местообитаний и формированием связанных с ними экосистем в этом варианте наиболее ярко выражены различия между видовым богатством ценофлор горнотаежных лесов и общим флористическим разнообразием пояса.
Патомский вариант. Широкое распространение горнотаежного пояса обусловлено относительно невысокими горными хребтами Патомского нагорья (до 1500–1700 м), в верхних частях которых он граничит с подгольцовым поясом; горные тундры получают развитие только на наиболее высоких хребтах. При фоновом участии лиственничных лесов заметную роль играют темнохвойные сообщества, приуроченные к более тепло- и влагообеспеченным низкогорьям в нижних частях долин правых притоков реки Лены. Большую часть высотного спектра занимают кедровостланиковые лиственничные леса (рис. 2). Особенно высока их роль в верхней части горнотаежного пояса на контакте с подгольцовыми стланиковыми и редколесными сообществами (на высотах 1000–1200 м), где формируются сложные гетерогенные экосистемы за счет взаимного проникновения их компонентов.
Для варианта характерна высотная дифференциация фитоценотического разнообразия горнотаежного пояса (табл. 3). На высотах 200–600 м развиты разные типы лиственничных, сосновых и темнохвойных лесов с достаточно равномерным распределением площадей, занимаемых сообществами. На высотах 600–800 м господствующее положение принадлежит лиственничным лесам (до 90%). Выше 800 м на лиственничные кедровостланиковые леса приходится более 4/5 площади.
Патомский вариант значительно уступает Северобайкальскому по уровню флористического разнообразия горнотаежного пояса (табл. 4). Это связано, прежде всего, с уменьшением роли видов южносибирского комплекса в сложении ценофлор горнотаежных формаций, а также с ограниченным развитием высокогорий, дополняющих разнообразие за счет элементов ценофлор горнотундровых сообществ. Около 1/4 видового состава сосудистых растений горнотаежного пояса составляют виды луговых и болотных экосистем, распространенных в нижних частях горных хребтов нагорья.
Кодаро-Каларский вариант. Формирование растительного покрова на наиболее массивных хребтах Станового нагорья обуславливает широкое распространение высокогорной растительности при более ограниченном по сравнению с другими вариантами распространением горнотаежных лесов. Характерно господство лиственничных лесов и незначительное участие сосновых лесов, имеющих локальное распространение в днищах межгорных котловин и нижних частях хребтов (до 700–800 м). По долинам рек на всем высотном протяжении пояса встречаются еловые леса, а на верхнюю границу леса (около 1100–1200 м) выходят редколесья с участием кедра, однако в силу локального распространения небольшими фрагментами они не находят отражения на мелкомасштабной карте. Среди лиственничных лесов господствующее положение занимают кедровостланиковые и кустарничково-зеленомошные типы.
Плавное изменение фитоценотического разнообразия характеризует Кодаро-Каларский вариант. На всех высотных уровнях горнотаежного пояса преобладают лиственничные кедровостланиковые и ерниковые леса (более 2/3 от лесопокрытой площади), соотношение площадей которых несколько варьирует (табл. 3). Сосновые и темнохвойные леса встречаются на высотах до 600 м, занимая около 10% площади. Структура фитоценотического разнообразия горнотаежного пояса этого варианта представлена в пределах одного подпояса лиственничных лесов.
Кодаро-Каларский вариант близок к Патомскому по уровню флористического разнообразия. Однако при общности его ядра соотношение сопутствующих видов различно. Вариант отличает повышенная роль высокогорных элементов за счет широкого развития подгольцового, горнотундрового и гольцового поясов, при достаточно тесном контакте с которыми развиты леса горнотаежного пояса, особенно его верхней части. При этом многие высокогорные виды принимают спорадическое участие в составе горнотаежных сообществ и не входят в состав постоянных видов их ценофлор. Также вклад в разнообразие вносит специфический комплекс видов псаммофитных сообществ, развитых на мощных аллювиальных отложениях в днищах многочисленных межгорных котловин вдоль байкальской рифтовой зоны (Дулепова, 2013). В целом для варианта отмечается наименьшая доля видов ценофлор горнотаежных лесов по отношению к общему флористическому разнообразию пояса (табл. 4).
Основные закономерности в структуре горнотаежного пояса оробиома
Пространственная организация биоразнообразия биома формируется под воздействием комплекса градиентов факторов, выраженных в соответствии с зональным, секторным и высотным его положением. Они характеризуют классическую трехмерную структуру горных территорий, находящую отражение в отдельных компонентах экосистем и экосистемах в целом. Современная структура биоразнообразия оробиома является результатом длительного исторического развития, в ходе которого ангаридские, берингийские, южносибирские комплексы растительных формаций находят свое положение в структуре высотной поясности и разнообразии экотопических условий. Дифференциация пояса на подпояс лиственнично-сосновых и кедрово-пихтово-еловых лесов и подпояс лиственничных лесов проходит в пределах 600–700 м. Нижний подпояс имеет небольшую высотную амплитуду (300–600 м), в пределах которой сохраняется высокая ценотическая значимость их сообществ, прежде всего, сосновых и лиственничных рододендроновых и кустарничково-зеленомошных лесов. Для них характерна достаточно высокая лесистость при заметном участии луговых и болотных экосистем в структуре растительного покрова. Верхний подпояс имеет большую высотную амплитуду, проникая до высот 1000–1200 м, где у верхней границы леса на контакте с подгольцовым поясом формируется сложная пространственная структура растительного покрова с постепенным снижением роли лиственничных лесов и увеличением доли высокогорных экосистем.
Лиственничные леса являются определяющими для высотно-поясного спектра в условиях холодного континентального климата Северного Забайкалья (рис. 1). Их широкое распространение в оптимальных условиях на высотах 600–900 м затушевывает разнообразие экотопических условий при выраженной климатической доминанте в структуре разнообразия горнотаежного пояса. В типологическом разнообразии преобладают леса кедровостланиковой, ерниковой и кустарничково-зеленомошной групп, при этом ценотический оптимум принадлежит кедровостланиковым лиственничным лесам, занимающим до 40% площади в пределах большей части высотной амплитуды пояса.
Дополняют типологическое разнообразие горнотаежного пояса ограниченно распространенные сосновые, пихтово-кедровые и еловые леса. Сосновые леса сосредоточены в более теплообеспеченных низкогорьях хребтов, а темнохвойные леса приурочены к более влажным и менее континентальным условиям западной части оробиома (Северо-Байкальское нагорье). Динамические тенденции в развитии лесного покрова оробиома проявляются в сукцессионых рядах производных сообществ. Восстановление основных групп климаксных горнотаежных лесов после пожаров и рубок идет через сообщества березовых (береза плосколистная) лесов. Они занимают около 3% от лесопокрытой площади оробиома и будут иметь тенденцию к увеличению площади в связи с возрастанием антропогенного влияния. Дополняют экосистемное разнообразие пояса долинные ряды лугово-болотной растительности, формирующиеся в межгорных котловинах с обширными днищами в нижней части высотного спектра. На крутосклонных поверхностях коренных склонов сводово-глыбовых поднятий формируется мозаичная структура из лиственничных редколесий, ерниковых и кедровостланиковых сообществ, каменистых россыпей, особенно у верхней границы леса на контакте с высокогорными поясами.
На фоне преобладающих экосистем лиственничных лесов и редколесий горнотаежного пояса биома в его составе встречаются редкие сообщества. Реликтовый вид береза шерстистая (Betula lanata) образует самостоятельную полосу криволесий у верхней границы леса в диапазоне высот 1100–1300 м. В контактной полосе ее сообщества сочетаются с зарослями кедрового стланика и ольховника; она встречается в составе лиственничных редколесий, также спускается в горнотаежный пояс и участвует в составе лиственничных и еловых лесов верхней полосы. Реликтовый вид карагана гривастая (Caragana jubata) встречается в составе лиственничных редколесий (1100–1600 м). Ее ценотический ареал находится в горах южной части Байкальского региона, где она образует самостоятельные сообщества в пределах высот 1700–2200 м в составе подгольцового пояса.
ЗАКЛЮЧЕНИЕ
Результаты проведенного исследования экологической организации бореальных горнотаежных лесов Северного Забайкалья подтвердили представление о важной роли дифференциации их экосистемного и биотического разнообразия в связи с биоклиматическими условиями высотно-поясных спектров горных биомов. В работе удалось выявить и показать на карте типологическое разнообразие лесных экосистем с оценкой их ценофлор. Оробиомы характеризуют региональное единство экосистем, в пределах которого сохраняется определенный уровень разнообразия биоты и ее пространственной (высотно-поясной и внутрипоясной) структуры. Успешное обращение к оробиомам при анализе флористического и фитоценотического разнообразия различных горных территорий определяет перспективы выявления типологического разнообразия лесов и разработки системы их мониторинга на единой биомной основе (Огуреева, 2012).
Наиболее фундаментальной проблемой в российском лесном хозяйстве является необходимость замены экстенсивного эксплуатационного характера лесопользования на модель устойчивого интенсивного лесоуправления (Леса …, 2020). В этом контексте является актуальным оценка типологического разнообразия и современного состояния горных лесов, выявление закономерностей их пространственно-временной организации, которые важны и решаемы в рамках лесной экологии и востребованы в практической сфере лесопользования и природоохранной деятельности. При возрастающей роли лесных ресурсов в экономике страны усиливается пресс на лесные экосистемы, понижается уровень биоразнообразия в связи с трансформацией фоновых и редких лесных экосистем, снижается их резистентность к неблагоприятным воздействиям. Помимо того, прогнозируются существенные изменения в структуре разнообразия растительного покрова при наблюдаемых глобальных тенденциях в изменении климата (Johnson et al., 2011). Особенно актуальны данные проблемы для зоны Байкало-Амурской магистрали, где планируется интенсификация освоения природных, в том числе лесных ресурсов, и отмечаются высокие тренды потепления климата, что находит отклик в разнообразии и структуре растительного покрова (Kharuk et al., 2017).
В настоящее время имеется только один Витимский государственный заповедник, территория которого охватывает небольшие участки Станового и Патомского нагорий, слабо изученных в биогеографическом отношении. В этой связи возникает насущная необходимость развития сети ООПТ для сохранения горных экосистем малонарушенных лесов, особенно редких сообществ и видов, генофонда и ценофонда горной тайги вдоль интенсивно развивающейся трассы БАМ, что будет способствовать сохранению биоразнообразия и экологических функций лесов.
Список литературы
Атлас малонарушенных лесных территорий России / Под. ред. Д.Е. Аксенова, Д.В. Добрынина, М.Ю. Дубинина и др. М.: МСоЭС; Вашингтон: World Resources Inst., 2003. 187 с.
Барталев С.А., Егоров В.А., Жарко В.О., Лупян Е.А., Плотников Д.Е., Хвостиков С.А. Состояние и перспективы развития методов спутникового картографирования растительного покрова России // Современные проблемы дистанционного зондирования Земли из космоса. 2015. Т. 12. № 5. С. 203–221.
Барталев С.А., Егоров В.А., Жарко В.О., Лупян Е.А., Плотников Д.Е., Хвостиков С.А., Шабанов Н.В. Спутниковое картографирование растительного покрова России. М.: ИКИ РАН, 2016. 208 с.
Белов А.В. Карта растительности юга Восточной Сибири. Принципы и методы составления // Геоботаническое картографирование. 1973. С. 16–30.
Биоразнообразие биомов России. Равнинные биомы. Под ред. Г.Н. Огуреевой. М.: ФГБУ “ИГКЭ”, 2020. 623 с.
Бочарников М.В. Роль климата в пространственной организации растительного покрова Кодаро-Каларского оробиома // Сибирский экологический журнал. 2019. Т. 26. № 3. С. 239–252.
Бочарников М.В. Эндемизм во флоре Северовосточно-Забайкальского оробиома // Проблемы ботаники Южной Сибири и Монголии. Барнаул: АлтГУ, 2020. Т. 19. № 2. С. 229–233.
Вальтер Г. Растительность Земного шара. Т. 1. М.: Прогресс, 1968. 551 с.
Доклад о состоянии и использовании лесов Российской Федерации за 2015 год Министерство природных ресурсов и экологии РФ. 2015.
Дулепова Н.А. Растительность развеваемых песков Верхнечарской котловины (Забайкальский край) // Растительность России. 2013. № 22. С. 29–32.
Иванова М.М., Чепурнов А.А. Флора западного участка районов освоения БАМ. Новосибирск, 1983. 223 с.
Карта “Биомы России” в серии карт природы для высшей школы (м. 1 : 7 500 000) / Под. ред. Огуреевой Г.Н., Леоновой Н.Б., Емельяновой Л.Г. и др. М.: Всемирный фонд дикой природы (WWF), 2018.
Курнаев С.Ф. Лесорастительное районирование СССР. М.: Наука, 1973. 203 с.
Леса России и изменение климата. Что нам может сказать наука / Под ред. П. Лескинен, М. Линднер, П. Веркерк и др. Европейский институт леса, 2020. 140 с.
Малышев Л.И. Флористическое богатство СССР // Актуальные проблемы сравнительного изучения флор. СПб.: Наука, 1994. С. 34–87.
Малышев Л.И., Байков К.С, Доронькин В.М. Флористическое деление Азиатской России на основе количественных признаков // Krylovia. Т. 2. № 1. 2000. С.3–16.
Малышев Л.И., Пешкова Г.А. Особенности и генезис флоры Сибири (Предбайкалье и Забайкалье). Новосибирск: Наука, 1984. 264 с.
Морозов Г.Ф. Учение о лесе. М.-Л.: Госиздат, 1949. 450 с.
Назимова Д.И., Пономарев Е.И., Степанов Н.В., Федотова Е.В. Черневые темнохвойные леса на юге Красноярского края и проблемы их обзорного картографирования. Лесоведение. 2005. № 1. С. 12–18.
Намзалов Б.Б. Важнейшие узлы биоразнообразия и фитогеографические феномены горных степей Южной Сибири // Аридные экосистемы. Т. 27. № 3. 2021. С. 24–36.
Огуреева Г.Н. Структура высотной поясности растительного покрова Байкало-Джугджурской области // Вестн. Моск. ун-та. Серия 5. География. 1999. № 2. С. 50–56.
Огуреева Г.Н. Эколого-географический подход к изучению разнообразия и географии наземных экосистем // Вопросы географии. Моск. отд. РГО Т. 134. Сб. Актуальная биогеография. М.: Кодекс, 2012. С. 58–80.
Огуреева Г.Н., Бочарников М.В. Оробиомы как базовые единицы региональной оценки биоразнообразия горных территорий // Экосистемы: экология и динамика. 2017. Т. 1. № 2. С. 52–81.
Одум Ю. Экология. Т. 1. М.: Мир, 1986. 328 с.
Панарин И.И., Митрофанов Л.П., Исаева Л.Н. Горные леса зоны БАМ. Новосибирск: Наука, 1980. 224 с.
Поликарпов Н.П., Чебакова Н.М., Назимова Д.И. Климат и горные леса Южной Сибири. Новосибирск: Наука. Сиб. отд-ние, 1986. 225 с.
Рысин Л.П. Лиственничные леса России. М.: Товарищество научных изданий КМК, 2010. 343 с.
Состояние лесов мира. Леса, биоразнообразие и люди. Рим: ФАО, 2020. 201 с.
Сочава В.Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 318 с.
Сукачев В.Н. Основы лесной типологии и биогеоценологии. М.: Наука, 1972. 419 с.
Тишков А.А. Ценофонд: пути формирования и роль сукцессий // Биологическое разнообразие: подходы к изучению и сохранению. СПб: ЗИН РАН, 1992. С. 21–34.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
FAO. Global Forest Resources Assessment 2020: Main report. Rome, 2020.
Holdridge L.R. Determination of world plant formations from simple climate data // Science. 1947. V. 105. P. 367–368.
Johnson K.G., Brooks S.J., Fenberg P.B., Glover A.G., James K.E., Lister A.M., Michel E., Spencer M., Todd J.A., Valsami-Jones E., Young J.R., Stewart J.R. Climate change and biosphere response: Unlocking the collections vault // BioScience. 2011. V. 61. № 2. P. 147–153.
Karger D.N., Conrad O., Böhner J., Kawohl T., Kreft H., Soria-Auza R.W., Zimmermann N.E., Linder H.P., Kessler M. Climatologies at high resolution for the earth’s land surface areas // Scientific Data. 2017. № 4.
Kharuk V.I., Im S.T., Petrov I.A., Golyukov A.S., Ranson K.J., Yagunov M.N. Climate-induced mortality of Siberian pine and fir in the Lake Baikal Watershed, Siberia // Forest Ecology and Management. 2017. V. 384. P. 191–199.
Köppen W. Das geographische System der Klimate // Handbuch der Klimatologie. Gebrüder Borntraeger, Berlin, 1936. P. 1–44.
Trewartha G.T., Horn L.H. Introduction to climate, 5th edn. McGraw Hill, New York, USA, 1980. 437 p.
Walter H. Vegetation of the Earth in relation to Climate and Eco physiological Conditions, Springer-Verlag, New York, 1973.
Walter H., Breckle S.-W. Okologishe Grundlagen in global sicht. Stuttgart: G. Fischer, 1991. 586 p.
Дополнительные материалы отсутствуют.