

Лесоведение, 2023, № 2, стр. 150-160
Предикторы повреждения насекомыми лесных насаждений по спутниковым данным на примере сибирского шелкопряда
А. В. Ковалев a, *, П. Е. Цикалова a
a Красноярский научный центр СО РАН
660036 Красноярск, Академгородок, д. 50, Россия
* E-mail: sunhi.prime@gmail.com
Поступила в редакцию 17.06.2022
После доработки 18.06.2022
Принята к публикации 18.06.2022
- EDN: AKBVXY
- DOI: 10.31857/S002411482302002X
Аннотация
Вспышки массового размножения сибирского шелкопряда (Dendrolimus sibiricus Tschetv.) в таежных лесах Сибири начинаются с очагов площадями в единицы га и достигают площадей в сотни тысяч га, что ведет к значительным повреждениям лесов. Границы очагов изменяются во времени и пространстве в зависимости от внешних факторов, динамики популяции и состояния кормовых деревьев. Борьбу с вредителем необходимо начинать за 1–2 года до достижения пика плотности популяции насекомого. В связи с этим важно знать предикторы повышения численности вредителя, заблаговременно определять момент начала вспышек и зоны, в пределах которых возникают вспышки. Для оценки необходимых условий возникновения вспышки предлагается метод оценки состояния лесных насаждений по данным дистанционного зондирования Земли (ДЗЗ).
Насекомые являются одним из основных факторов повреждении и гибели лесов. Так, на территории США в период с 2000 по 2016 гг. насекомые повредили около 8 миллионов га (Kosiba et al., 2018). Для многих видов характерны колебания плотности популяции, происходящие синхронно в нескольких местах, иногда с удивительной регулярностью во времени (Liebhold et al., 2004). Лесные насекомые, например: еловая листовертка (Choristoneura fumiferana Clem.), лиственничная листовертка (Zeiraphera griseana Hb.), непарный шелкопряд (Lymantria dispar L.), осенне-зимняя огневка – подвержены сильным повторяющимся колебаниям численности с повышением плотности популяции на несколько порядков (Williams, Liebhold, 2000; Bjørnstad et al., 2002; Ims et al., 2004; Johnson et al., 2006; Tenow et al., 2012). Вспышки лесных насекомых вызывают крупномасштабную дефолиацию и гибель видов деревьев-хозяев, некоторые из поврежденных лесных насаждений простираются на миллионы гектаров. Вспышки насекомых изменяют сукцессию и состав лесов, а также круговорот питательных веществ, что имеет серьезные последствия для функций и услуг экосистемы (Boyd et al., 2013).
Вспышки вредителей могут сильно повлиять на состояние экосистемы и, следовательно, иметь далеко идущие последствия для местной экономики, зависящей от конкретных экосистемных услуг (Lewis et al., 2010; Fahse, Heurich, 2011; Nelson et al., 2013). В настоящее время установлено, что вспышки обусловлены факторами разного масштаба, т. е. численностью паразитов на уровне древостоя, взаимосвязанностью на уровне ландшафта и влиянием климата на региональном уровне (Senf et al., 2017; Seidl et al., 2016). Эти выводы совпадают с глобальным ростом повреждений от насекомых, в том числе под влиянием антропогенного изменения климата (Seidl et al., 2017; Kautz et al., 2017). Эмпирические данные подтверждают актуальность этих многочисленных факторов, но комплексное понимание их соответствующих ролей, их взаимодействий и результирующей сложной динамики все еще отсутствует, что ограничивает способность прогнозировать вспышки вредителей. В связи с этим существует потребность в лучшем количественном понимании причинно-следственных связей и межмасштабных взаимодействий, ведущих к крупномасштабным вспышкам (Raffa et al., 2008; Senf et al., 2017).
Как правило, динамика популяций вредителей описывается на уровне насаждений с использованием моделей популяций или системной динамики (Fernández, Fort, 2009; Lewis et al., 2010; Fahse, Heurich, 2011). Разработка, параметризация и проверка моделей динамики численности лесных насекомых, основанных на процессах, связанных с динамикой на уровне местной популяции, сопровождаются двумя основными трудностями: (1) построение таких моделей требует всестороннего знания многих процессов, происходящих на уровне местной популяции, и (2) для моделирования необходима оценка плотности насекомых. Однако эти требования трудно выполнимы, поскольку взаимосвязь между процессами в насаждениях и динамикой популяций насекомых в основном изучается в отдельных местах (Royama, 1984). Более того, полученные выводы могут быть слабыми или ненадежными (Peters et al., 2004).
Это, однако, не показывает, как локальные явления трансформируются в крупномасштабную динамику. Хотя существуют модели, описывающие пространственную динамику (Zhou, Liebhold, 1995; Logan et al., 1998), многомасштабная перспектива вспышек вредителей в основном представлена через феноменологию (Peters et al., 2004; Raffa et al., 2008) или эмпирический подход (Bjørnstad et al., 2002; Seidl et al., 2016; Senf et al., 2017).
Сибирские леса являются одним из крупнейших на нашей планете экологических объектов. Они занимают площадь 2.6 млн км2, а запас древесины превосходит 30–36 млрд м3. В Восточно-Сибирской тайге преобладают светлохвойные лиственничные леса. Основными факторами гибели хвойных лесов Сибири являются лесные пожары, а на втором месте по площадям повреждения находится воздействие лесных насекомых. В лесах Сибири и Дальнего Востока наиболее вредоносным насекомым-фитофагом является сибирский шелкопряд (Dendrolimus sibiricus Tschetv.) (Коломиец, 1962; Рожков, 1965; Болдаруев, 1969; Плешанов, 1982; Эпова, Плешанов, 1995; Юрченко, Турова, 2002; Михайлов, Сумина, 2012; Павлов и др., 2018). Суммарные площади очагов массового размножения сибирского шелкопряда на территории Сибири составляют десятки миллионов гектаров леса (Эпова, Плешанов, 1995).
Однако для успешной борьбы с вредителями необходима комплексная количественная оценка перспективы вспышек вредителей, которая может быть получена как сумма вероятностей необходимых условий реализации вспышки на данной территории. Развитие вспышек определяется совокупностью факторов: погодных (температурой и осадками в течение сезона), трофических (состоянием корма), физиологических (состоянием особей вредителей и устойчивостью древесных растений к нападению вредителей), а также численностью популяций вредителей (Кондаков, 1974). Для сохранения лесов важно оценить риски вспышек и заблаговременно определить моменты начала вспышек и территории, в пределах которых начинаются вспышки.
Однако для обширных лесов Сибири вести регулярный наземный мониторинг на всей территории и регулярно оценивать плотность популяций вредителей технически и экономически невозможно. В связи с этим необходимы методы, позволяющие заблаговременно выявить территории, на которых в ближайшее время возможны вспышки массового размножения и в пределах которых можно вести локальный мониторинг. Настоящая работа посвящена описанию метода, позволяющего заблаговременно выявить насаждения, в которых за счет ослабления регуляторных механизмов снижается устойчивость к ослаблению насекомыми, что является одним из основных условий вспышки вредителя. Исходные сведения для такой оценки будут получены по данным дистанционного зондирования земли, что позволяет проводить обследования на удаленных участках сибирской тайги.
ОБЪЕКТЫ И МЕТОДИКА
В настоящей работе в качестве объектов исследования выступали популяции сибирского шелкопряда. Анализ проводился для пихтовых насаждений таежной зоны в очаге массового размножения сибирского шелкопряда на территории Енисейского района Красноярского края. Вспышка массового размножения произошла в 2015–2017 гг. В ходе вспышки насекомые повредили и уничтожили около 1 млн га насаждений пихты сибирской (Abies sibirica Ledeb.) и сосны сибирской (Pinus sibirica (Loud.)).
Для оценки состояния насаждений изучались две трансекты. На рис. 1 показано расположение начальных точек изучаемых трансект. Направление и длина трансект выбирались так, чтобы получить максимально однотипные по рельефу и растительному покрову точки наблюдений.
Рис. 1.
Расположение начальных точек изучаемых трансект. p1 – (58°42′19.15″ N; 91°1′44.60″ E), трансекта с поврежденными древостоями с 2015 года. p2 – (58°36′55.40″ N; 90°15′1.00″ E), тренсекта с неповрежденными древостоями.

Расстояние между начальными точками трансект – 46 км. Шаг между точками для каждой трансекты – 1 км. Трансекта 1 длиной около 11 км – в юго-западном направлении, все насаждения на трансекте одновременно повреждались с 2015 года; трансекта 2 длиной 23 км – в западном направлении c неповрежденными до 2020 года насаждениями.
В настоящий момент спутниковые дистанционные оценки состояния насаждений используются в основном для определения нанесенного насекомыми ущерба. При этом одним из основных методов, используемых при анализе, является измерение различных вариантов вегетационного индекса растительности, основанного на разнице отражения красного и ближнего инфракрасного излучения (Tucker, Sellers 1986). Вегетационный индекс позволяет оценить продуктивность и физиологические свойства растительной компоненты экосистемы (Liu et al., 2017; Rechid et al., 2009) и представляет собой спектральный индикатор фотосинтеза и интенсивности метаболизма растений (Bayarjargal et al., 2006; Cunha, Richter, 2014). Данные ДЗЗ широко используются для картирования пространственной динамики очагов вспышек насекомых (Verbesselt et al., 2012; Liang et al., 2014; Senf et al., 2017). Однако основной целью таких исследований является оценка ущерба, нанесенного вспышкой, и расчет площади потери. Такой индекс достоверно и оперативно показывает деградацию кроны деревьев в ходе развития вспышки массового размножения насекомых. В то же время ослабление древостоев и снижение противодействия дерева насекомым развиваются в течение нескольких лет до начала резкого подъема численности вредителя, но не регистрируются по показателям NDVI, и попытки использовать данные дистанционного зондирования для оценки устойчивости лесных насаждений к внешним воздействиям не приводят к успеху (Spruce et al., 2011; Thayn, 2013; Olsson et al., 2016).
В настоящей работе при использовании дистанционных данных делается попытка перейти от количественных измерений биомассы растений по показателям абсолютных значений вегетативного индекса к оценке реакции фотосинтезирующего аппарата на изменения состояния окружающей среды. Входные и выходные параметры для такой модели могут быть получены одновременно по данным ДЗЗ и быть синхронизированы по времени и месту наблюдения. Мы использовали данные спутников Terra/Aqua, действующих в рамках программы NASA EOS (Earth Observing System). Исходные данные ДЗЗ со спутников Terra/Aqua доступны для свободного скачивания на сервере NASA. Основным прибором для сбора необходимой информации на спутнике является спектрорадиометр среднего разрешения MODIS (Moderate-Resolution Imaging Spectroradiometer). В качестве показателя, характеризующего состояние насаждений, в настоящей работе предложено использовать показатель восприимчивости вегетационного индекса растительность (NDVI) в течение сезона к изменению радиационной температуры земной поверхности (LST). В данной работе показатель NDVI вычисляется по стандартной формуле:
где NIR и Red – нормализованные значения интенсивности отражения в ближнем инфракрасном и красном диапазонах спектра для данной точки земной поверхности. Пространственное разрешение для данных показателей составляет 250 × 250 м. Типичные временные ряды годичной динамики NDVI и LST приведены на рис. 2.Рис. 2.
Типичные временные ряды годичной динамики NDVI и LST. (2010 г.), 1 – проба р101 (очаг с 2015 г.), 2 – проба р201 (контроль).
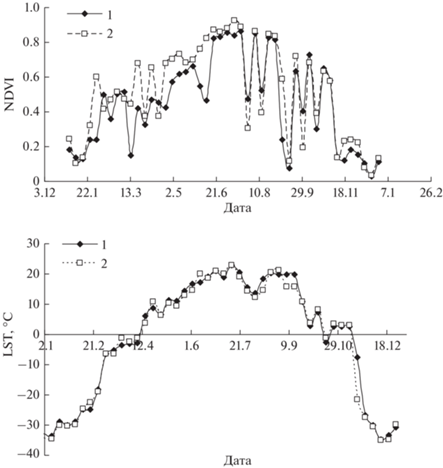
Поскольку рост и состояние кроны деревьев существенно зависит от изменений температуры окружающей среды, то анализ связей между NDVI и LST позволит оценить адаптивный ресурс деревьев в насаждении. В этом случае изменение NDVI можно рассматривать как некоторую функцию от изменений погодных условий и для отдельного года, переменную LST – как входную в системе, а переменную NDVI – как выходную. Если предполагать, что выходной сигнал с некоторым запаздыванием τ зависит от входного, то эту связь можно записать через некоторый оператор F:
Ранее было показано, что по сезонным временным рядам NDVI и LST можно оценить оператор F в виде спектральной функции отклика H(f) (Kovalev, Soukhovolsky, 2021):
где Фyx – кросс-корреляционная функция, связывающая ряды первых разностей NDVI(t) и LST(t) в течение сезона, Фxx – автокорреляционная функция ряда первых разностей LST, FT – спектральное преобразование Фурье.Полученная спектральная функция H(f) характеризует скорость и интенсивность воздействия погоды на состояние фотосинтетического аппарата древостоя. Типичный вид спектра функции отклика Н(f) представлен на рис. 3.
Рис. 3.
Типичный вид спектра функции Н(f) отклика NDVI на изменение LST (пробная площадь p103 в очаге массового размножения сибирского шелкопряда, 2016 г.).

Значение спектральной функции отклика H(f0) на некоторой частоте f0 характеризуют интенсивность и скорость v = 1/f0 отклика NDVI на изменение LST. Составляющие спектра функции отклика на низких частотах (не более f = 0.16) характеризуют сильно инерционную реакцию NDVI на изменение LST; составляющие спектра на высоких частотах (от f = 0.35 до f = 0.5) характеризуют быстрый отклик NDVI на изменение LST. При изменении физиологических свойств деревьев в насаждении реакции на внешние изменения температуры ускоряются или замедляются, что меняет соотношение высокочастотных и низкочастотных компонентов спектра.
В дальнейшем анализе используются мощность низкочастотной составляющей LF спектра (частоты от 0 до 0.16 1/день) и мощность высокочастотной составляющей спектра SH в диапазоне f = 0.35–0.50 1/день).
(4)
${\text{LF}} = \int\limits_{\text{0}}^{{\text{0}}{\text{.16}}} {H{\text{(}}f{\text{)}}df} ,\,\,\,\,{\text{HF}} = \int\limits_{0.35}^{0.50} {H(f)df} .$Пары этих значений сравниваются для поврежденных и контрольных насаждений.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Насколько величины NDVI в течение сезона характеризуют состояние насаждений? Для поврежденных и контрольных пробных площадей были вычислены средние значения ❬NDVI❭ фотосинтетического индекса в течение сезона продолжительностью t0:
(5)
$\left\langle {{\text{NDVI}}} \right\rangle = \frac{1}{{{{t}_{0}}}}\int\limits_0^{{{t}_{0}}} {{\text{NDVI}}(\tau )d\tau } .$Полученные данные NDVI и LST усреднялись для поврежденных и контрольных участков. На рис. 4 приведены отношения средних сезонных значений ❬NDVI❭ для насаждений в очаге и в контроле.
Рис. 4.
Динамика средних сезонных значений ❬NDVI❭ в очагах массового размножения сибирского шелкопряда (1) и в неповрежденном насаждении (2). 3 – стандартные ошибки ❬NDVI❭ для насаждений в зоне вспышки; 4 – стандартные ошибки ❬NDVI❭ для неповрежденных насаждений.
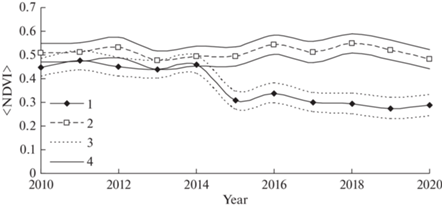
Из рис. 4 следует, что до начала вспышки (2015 г.) значимых различий NDVI деревьев в будущих очагах массового размножения и в контрольных неповрежденных насаждениях не наблюдалось. Только после повреждений хвои такие различия появились. Таким образом, по суммарным показателям NDVI не удается получить заблаговременную оценку рисков нападения насекомых на насаждения.
Далее рассмотрим возможность использования для заблаговременной оценки рисков нападения вредителей на насаждения спектральных функций отклика. Для сопоставления спектральных функций отклика поврежденных и неповрежденных пробных площадей удобно “свернуть” эту функцию и представить ее в виде двух параметров, характеризующих мощности спектра на низких LF и высоких HF частотах (4). В дальнейшем анализе использовались усредненные показатели по трансектам 1 и 2. На рис. 5 приведены средние характеристики компонентов LF и HF для трансект 1 и 2 в течение 2010–2020 гг.
Рис. 5.
Низкочастотные (LF) и высокочастотные (HF) характеристики функций отклика для насаждений на трансектах 1 и 2. 1 – трансекта 1, 2010–2020 гг.; 2 – трансекта 2, 2010–2011 гг.; 3 – трансекта 2, 2012–2014 гг.; 4 – трансекта 2, 2015–2017 гг.; 5 – трансекта 2, 2018, 2020 гг.
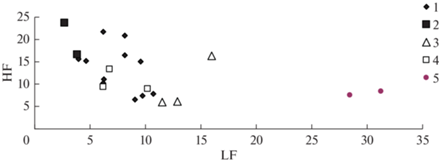
Как видно из рис. 5, данные для неповрежденных насаждений на трансекте 1 за все годы исследований характеризуются кластером в границах LF = 4–10 и HF = 7–22. Для насаждений, поврежденных в 2015 г., характеристики в 2010–2011 гг. близки к характеристикам насаждений для трансекты 1. Значения LF и HF для насаждений на трансекте 2 перед началом вспышки в 2012–2014 гг. отличаются от характеристик насаждений на трансекте 1. После сильных повреждений, когда насаждения потеряли хвою, характеристики LF и HF очень сильно отличались от соответствующих показателей неповрежденных насаждений.
Результаты более детального анализа показателей LF и HF для поврежденных сибирским шелкопрядом насаждений вдоль трансекты 2, начиная с 2015 г., приведены на рис. 6.
Рис. 6.
Низкочастотные (LF) и высокочастотные (HF) характеристики функций отклика для насаждений на трансекте 2. 1 – до вспышки, 2011г.; 2 – перед вспышкой, 2013 гг.; 3 – после повреждений, 2020 г.
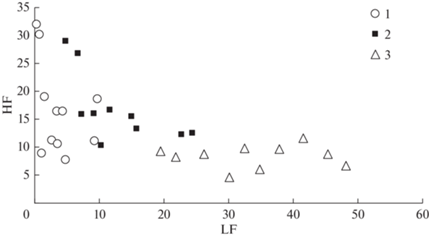
Как видно, характеристики функции отклика отличаются в разные периоды воздействия насекомых на насаждения. Для различения данных разных лет использовался линейный дискриминантный анализ. Анализ по критерию Wilks’ Lambda λW = 0.2040, при F (4.54) = 16.387 и p < < 0.00001 показал значимость различий значений LF и HF для разных лет. Согласно классификационной матрице (табл. 1), по показателям LF и HF возможно разделить более 80% насаждений вдоль трансекты 2.
Таблица 1.
Классификационная матрица для дискриминации насаждений вдоль трансекты 2
| Percent | G_1:1 | G_2:2 | G_3:3 | |
|---|---|---|---|---|
| G_1:1 | 90.91 | 10 | 1 | 0 |
| G_2:2 | 70.00 | 2 | 7 | 1 |
| G_3:3 | 80.00 | 0 | 2 | 8 |
| Total | 80.65 | 12 | 10 | 9 |
Таким образом, в 2013 г., т.е. за два года до начала вспышки, характеристики функций отклика уже отличались от соответствующих показателей за 4 года до начала вспышки. Это позволяет использовать данные дистанционного зондирования для заблаговременной оценки рисков повреждений насаждений.
Насколько пространственно однородны характеристики насаждений вдоль трансекты? Если значимых различий величин LF и HF в насаждениях вдоль трансекты нет, то регрессионное уравнение связи значений LF или HF с номером насаждения вдоль трансекты будет незначимым. Если насаждения вдоль трансекты 1, не повреждавшиеся сибирским шелкопрядом, будут однородны по характеристикам функций отклика, то связи между средними многолетними значениями ❬LF❭ и ❬HF❭ и номером n (n = 1, … 24) насаждения вдоль трансекты 1 будут незначимыми. На рис. 7 отражены эти связи между ❬LF❭, ❬HF❭ и n.
Рис. 7.
Пространственные связи для показателей ❬LF❭ (1) и ❬HF❭ (2) для отдельных участков вдоль трансекты 1.

Как следует из рис. 7, пространственные корреляции показателей LF и HF вдоль трансекты 1 незначимы, то есть характеристики не поврежденных сибирским шелкопрядом насаждений однородны на протяжении по меньшей мере 24 км, и можно полагать, что состояние насаждений вдоль этой трансекты достаточно однородно.
Эффективный энтомологический мониторинг должен позволять определить локальную зону и время начала вспышки. Одним из условий начала вспышки массового размножения вредителей являются необходимые показатели погоды. Однако воздействие погодных факторов может оказать влияние на все таежные территории, и выделить зону возможных вспышек по этим показателям невозможно. Площади зон эффективного воздействия погодных факторов также могут быть очень велики – до 90 000 км2 (Исаев и др., 2015), что затрудняет определение зон будущих вспышек. К тому же снижение ГТК ниже 1 не является необходимым условием развития вспышки, и часто информация о сухой и теплой погоде на территории не позволяет однозначно говорить о будущей реализации вспышки сибирского шелкопряда (Тарасова, Волков, 2021).
Более надежным показателем места развития будущей вспышки сибирского шелкопряда являются характеристики низко- и высокочастотных составляющих функции отклика NDVI на изменение LST в течение сезона. Выполненные расчеты показывают, что характеристики функций отклика начинают отличаться от нормы за два сезона до начала вспышки. Таким образом, по дистанционным данным показателей LF, HF можно определить изменения состояния насаждений и предсказать возможные повреждения древостоя. Относительный рост значений LF и снижение HF по сравнению с контрольными насаждениями можно трактовать как уменьшение скорости реакции деревьев в насаждении в ответ на возможное изъятие листового аппарата деревьев и использовать как индикатор ухудшения состояния насаждения и повышения риска атаки на него насекомых-вредителей. Использование методов дистанционного зондирования для оценки состояния и устойчивости древесных растений открывает возможность диагностики состояния древесных растений в любой точке планеты без необходимости наземного обследования территорий.
ЗАКЛЮЧЕНИЕ
Предложенный метод позволяет достаточно надежно оценить риски вспышек сибирского шелкопряда. Для выявления зоны вспышки необходимы расчеты значений компонентов функций отклика на территории. Следует отметить, что используемые дистанционные данные разрешением 250м доступны и могут быть получены с сайта NASA для любой точки на планете. Но в настоящее время активно вводятся в эксплуатацию отечественные и зарубежные спутники с разрешение до 1 м. Наличие подобных регулярных наблюдений показателей NDVI и LST позволит существенно повысить точность оценки состояния насаждений. При использовании мощных вычислительных возможностей это открывает возможность для мониторинга рисков вспышек сибирского шелкопряда на всей территории тайги. Для проверки возможности оценки рисков возникновения вспышек других видов лесных насекомых необходимы дополнительные исследования.
Список литературы
Болдаруев В.О. Динамика численности сибирского шелкопряда и его паразитов. Улан-Удэ: Бурят. книж. из-во, 1969. 162 с.
Исаев А.С., Пальникова Е.Н., Суховольский В.Г., Тарасова О.В. Динамика численности лесных насекомых-филлофагов: модели и прогнозы. М.: Товарищество научных изданий КМК, 2015. 276 с.
Коломиец Н.Г. Паразиты и хищники сибирского шелкопряда. Новосибирск: Наука, 1962. 172 с.
Кондаков Ю.П. Закономерности массовых размножений сибирского шелкопряда // Экология популяций лесных животных Сибири. Новосибирск: Наука, 1974. С. 206–265.
Михайлов Ю.З., Сумина Н.Ю. Сибирский шелкопряд Dendrolimus superans (Butler, 1877) и борьба с ним в Иркутской области // Байкальский зоологический журн. 2012. № 3(11). С. 25–29.
Павлов И.Н., Литовка Ю.А., Голубев Д.В., Астапенко С.А., Хромогин П.В. Новая вспышка массового размножения Dendrolimus sibiricus Tschetv. в Сибири (2012–2017 гг.): закономерности развития и перспективы биологического контроля // Сибирский экологический журн. 2018. № 4. С. 462–478.
Плешанов А.С. Насекомые – дефолианты лиственничных лесов Восточной Сибири. Новосибирск: Наука, 1982. 209 с.
Рожков А.С. Массовое размножение сибирского шелкопряда и меры борьбы с ним. М.: Наука, 1965. 178 с.
Тарасова О.В., Волков В.Е. Влияние погодных условий на развитие вспышек массового размножения сибирского шелкопряда в Средней Сибири // Сибирский лесной журн. 2021. № 5. С. 49–59.
Эпова В.И., Плешанов А.С. Зоны вредоносности насекомых-филлофагов Азиатской России. Новосибирск: Наука. Сибирская издательская фирма РАН. 1995. 147 с.
Юрченко Г.И., Турова Г.И. Паразиты сибирского и белополосого шелкопрядов на Дальнем Востоке России // Энтомологические исследования в Сибири. Вып. 2. Красноярск: КФ РЭО. 2002. С. 75–86.
Bayarjargal Y., Karnieli A., Bayasgalan M., Khudulmur S., Gandush C., Tucker C.J. A comparative study of NOAA-AVHRR derived drought indices using change vector analysis // International J. Remote Sensing. 2006. V. 105. № 1. P. 9–22.
Bjørnstad O.N., Peltonen M., Liebhold A.M., Baltensweiler W. Waves of larch budmoth outbreaks in the European Alps // Science. 2002. V. 298. № 5595. P. 1020–1023.
Boyd I.L., Freer-Smith P.H., Gilligan C.A., Godfray H.C.J. The consequence of tree pests and diseases for ecosystem services // Science. 2013. V. 342. P. 823.
Cunha M., Richter C. A time-frequency analysis on the impact of climate variability with focus on semi-natural montane grassland meadows // IEEE Trans. Geosci. Remote Sens. 2014. V. 52. № 10. P. 6156–6164.
Fahse L., Heurich M. Simulation and analysis of outbreaks of bark beetle infestations and their management at the stand level // Ecological Modelling. 2011. V. 222. № 11. P. 1833–1846.
Fernández A., Fort H. Catastrophic phase transitions and early warnings in a spatial ecological model // J. Statistical Mechanics: Theory and Experiment. 2009. V. 9. P. P09014.
Ims R.A., Yoccoz N.G., Hagen S.B. Do sub-Arctic winter moth populations in coastal birch forest exhibit spatially synchronous dynamics? // J. Animal Ecology. 2004. V. 73. P. 1129–1136.
Johnson D.M., Liebhold A.M., Tobin P.C. Bjørnstad O.N. Allee effects and pulsed invasion by the gypsy moth // Nature. 2006. V. 444. P. 361–363.
Kautz M., Meddens A.J.H., Hall R.J. Arneth A. Biotic disturbances in northern hemisphere forests-a synthesis of recent data, uncertainties and implications for forest monitoring and modeling // Global Ecology and Biogeography. 2017. V. 26. № 5. P. 533–552.
Kosiba A.M., Meigs G.W., Tait E.R. Spatiotemporal patterns of forest damage and disturbance in the northeastern United States: 2000–2016 // Forest Ecology and Management. 2018. V. 430. P. 94–104.
Kovalev A., Soukhovolsky V. Analysis of Forest Stand Resistance to Insect Attack According to Remote Sensing Data // Forests. 2021. V. 12. P. 1188. https://doi.org/10.3390/f12091188
Lewis M.A., Nelson W., Xu. C. A structured threshold model for mountain pine beetle outbreak // Bulletin of Mathematical Biology. 2010. V. 72. P. 565–589.
Liang L., Chen Y., Hawbaker T., Zhu Z., Gong P. Mapping mountain pine beetle mortality through growth trend analysis of time-series landsat data // Remote Sens. 2014. V. 6. P. 5696–5716.
Liebhold A.M., Koenig W.D., Bjørnstad O.N. Spatial synchrony in population dynamics // Annual Review of Ecology, Evolution, and Systematics. 2004. V. 35. P. 467–490.
Liu Yan, Hill M.J., Zhang Xiaoyang, Wang Zhuosen, Richardson A.D., Hufkens K., Filippa G., Baldocchi D.D., Ma Siyan, Verfaillie J., Schaaf C.B. Using data from Landsat, MODIS, VIIRS and Pheno Cams to monitor the phenology of California oak/grass savanna and open grassland across spatial scales // Agricultural and Forest Meteorology. 2017. V. 237–238. P. 311–325.
Logan J.A., White P., Bentz B.J., Powell J.A. Model analysis of spatial patterns in mountain pine beetle outbreaks // Theoretical Population Biology. 1998. V. 53. № 3. P. 236–255.
Nelson W.A., Bjørnstad O.N., Yamanaka T. Recurrent insect outbreaks caused by temperature-driven changes in system stability // Science. 2013. V. 341. № 6147. P. 796–799.
Olsson P.O., Lindstrom J., Eldundh L. Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI // Remote Sensing of Environment. 2016. V. 181. P. 42–53.
Peters D.P.C., Pielke R.A., Bestelmeyer B.T., Allen C.D., Munsonmcgee S., Havstad K.M. Cross-scale interactions, nonlinearities, and forecasting catastrophic events // Proceedings of the National Academy of Sciences. 2004. V. 101. P. 15130–15135.
Raffa K.F., Aukema B.H., Bentz B.J., Carroll A.L., Hicke J.A., Turner M.G., Romme W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions // Bioscience. 2008. V. 58. № 6. P. 501–517.
Rechid D., Raddatz T.J., Jacob D. Parameterization of snow-free land surfacealbedo as a function of vegetation phenology based on MODIS data and appliedin climate modelling // Theoretical and Applied Climatology. 2009. V. 95. P. 245–255.
Royama T. Population dynamics of the spruce budworm Choristoneura fumiferana // Ecological Monographs. 1984. V. 54. № 4. P. 429–462.
Seidl R., Müller J., Hothorn T., Bässler C., Heurich M., Kautz M. Small beetle, large-scale drivers: how regional and landscape factors affect outbreaks of the european spruce bark beetle // J. Applied Ecology. 2016. V. 53. № 2. P. 530–540.
Seidl R., Thom D., Kautz M., Martin-Benito D., Peltoniemi M., Vacchiano G., Wild J., Ascoli D., Petr M., Honkaniemi J. Forest disturbances under climate change // Nature Climate Change. 2017. V. 7. № 6. P. 395.
Senf C., Campbell E.M., Pflugmacher D., Wulder M.A., Hostert P. A multi-scale analysis of western spruce budworm outbreak dynamics // Landscape Ecology. 2017. P. 1–14.
Spruce J.P., Sader S., Ryan R.E., Smoot J., Kuper P., Ross K., Prados D., Russell J., Gasser G., McKellip R., Hargrove W. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks // Remote Sensing of Environment. 2011. V. 115. P. 427–437.
Tenow O., Nilssen A.C., Bylund H. Geometrid outbreak waves travel across Europe // J. Animal Ecology. 2012. V. 82. № 1. P. 84–95.
Thayn J.B. Using a remotely sensed optimized Disturbance Index to detect insect defoliation in the Apostle Islands, Wisconsin, USA // Remote Sensing of Environment. 2013. V. 136. P. 210–217.
Tucker C.J., Sellers P.J. Satellite remote sensing of primary production // J. Remote Sensing. 1986. V. 7. P. 1395–1416.
Verbesselt J., Zeileis A., Herold M. Near real-time disturbance detection using satellite image time series // Remote Sensing of Environment. 2012. V. 123. P. 98–108.
Williams D.W., Liebhold A.M. Spatial synchrony of spruce budworm outbreaks in eastern North America // Ecology. 2000. V. 81. P. 2753–2766.
Zhou G., Liebhold A.M. Forecasting the spatial dynamics of gypsy moth outbreaks using cellular transition models // Landscape Ecology. 1995. V.10. № 3. P. 177–189.
Дополнительные материалы отсутствуют.