

Лесоведение, 2023, № 2, стр. 174-189
Развитие очагов массового размножения лесных насекомых на разных пространственных масштабах
В. Г. Суховольский a, *, Ю. Д. Иванова b, А. В. Ковалев c
a Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, д. 50/28, Россия
b Институт биофизики СО РАН
660036 Красноярск, Академгородок, д. 50/50, Россия
c Федеральный исследовательский центр КНЦ СО РАН
660036 Красноярск, Академгородок, д. 50, Россия
* E-mail: soukhovolsky@yandex.ru
Поступила в редакцию 30.05.2022
После доработки 31.08.2022
Принята к публикации 18.10.2022
- EDN: ARRVOL
- DOI: 10.31857/S0024114823020092
Аннотация
Работа посвящена обсуждению возможностей описания динамики развития очагов массового размножения лесных насекомых на разных пространственных масштабах. Свойства очагов массового размножения рассмотрены на микропространственном масштабе, где анализируются модели распределения особей по кормовым объектам в границах локальной территории или насаждения, и на макропространственном масштабе, где рассмотрены показатели изъятия фотосинтезирующего аппарата (листьев или хвои) по всей территории очага, включая первичные, вторичные и миграционные очаги. При анализе микропространственного распределения гусениц на деревьях в очагах на разных фазах градации была использована модель распределения особей на кормовых деревьях как фазового перехода второго рода. Макропространственные процессы, происходящие в ходе роста очага, включают, во-первых, рост существующего очага и появление новых связных поврежденных участков леса, во-вторых, появление новых несвязных вторичных очагов. В качестве характеристик очагов массового размножения используются их фрактальная размерность D и характеристики “вязких пальцев” на границе очага. Для расчетов этих характеристик применены данные спутникового зондирования. Предлагаемые подходы могут быть использованы для прогноза развития очагов массового размножения лесных насекомых. При построении и верификации моделей применены данные учетов численности сибирского шелкопряда и заселенности деревьев в очагах его массового размножения, дистанционные данные по площадям и форме очагов в районах Красноярского края в ходе вспышки массового размножения сибирского шелкопряда (Dendrolimus sibiricus Tschetv.) в 2015–2019 гг.
Насекомые являются вторым по значимости (после пожаров) фактором повреждения и гибели лесных насаждений в России вследствие развития вспышек массового размножения вредителей. Площади повреждения лесов насекомыми варьируют в разные годы в пределах от 10 до 24% общей площади повреждения лесов. Так, площадь очагов лесных насекомых в 2020 г. составляла около 3 млн га леса. Ущерб от воздействия насекомых включает, во-первых, упущенную выгоду от прекращения депонирования углерода поврежденными деревьями, оцениваемую, например, для вспышек в 2020 г. в 3.1 млн т при значениях ЧПП от 3 до 5 т га год–1 (Soukhovolsky, Ivanova, 2020). Во-вторых, вспышка сопровождается выбросами углерода в атмосферу в виде СО2 в ходе потребления фитомассы насекомыми. Расчеты показывают, что при средней массе хвои на 1 га в 3.5–4 т с 1 га будет выделено в атмосферу в пересчете на чистый углерод примерно 1.5 т С (Суховольский и др., 2006; Тарасова и др., 2015). В целом на первом этапе вспышки (до начала пожаров и гниения стволовой древесины) на территории России в 2020 г. в ходе вспышек массового размножения лесных насекомых было выброшено в атмосферу в пересчете на чистый углерод около 4.5 млн т С.
В процессе развития вспышки массового размножения лесных насекомых возникают очаги массового размножения, в которых происходит нападение насекомых на деревья, ведущее к гибели насаждений. В процессе развития вспышки массового размножения лесных насекомых наблюдаются очаги массового размножения различных типов – первичные очаги, в границах которых и произошло увеличение численности популяции вредителей, а также вторичные миграционные очаги, развитие которых связано с локальным перемещением особей в насаждении (Кондаков, 1974; Исаев и др., 2001). Миграционные очаги обычно расположены достаточно далеко от первичных очагов и возникают при массовых миграциях особей вредителей на стадии имаго. При этом предполагается, что особи в популяции вредителя на преимагинальных стадиях сезонного развития относительно малоподвижны и только на стадии имаго насекомые могут передвигаться на значительные расстояния. Совокупность первичного, вторичных и миграционных очагов образует очаг массового размножения вида, характеризуемый некоторой суммарной площадью. Риск повреждения и гибели лесных насаждений связан с динамикой развития очагов.
Достаточно часто рассматривались “точечные” модели вспышек, в которых предполагают, что особи вредителей сосредоточены в некотором локальном местообитании, в ходе вспышки изменяется только плотность популяций вредителей. Однако такие модели не позволяют описывать перемещение насекомых и смену кормовых объектов – деревьев в процессе вспышки. В связи с этим анализ и моделирование развития очагов массового размножения лесных насекомых являются важными задачами, имеющими как теоретическое, так и практическое значение для оценок рисков повреждений насаждений вредителями и планирования лесозащитных мероприятий.
При анализе точечных моделей, т.е. моделей, рассматривающих популяцию в максимально агрегированном виде, когда учитывается только суммарная численность популяции и не учитывается распределение особей по территории очага, обычно ограничиваются анализом типа пространственного распределения насекомых на пробных площадях, выделяя случайный, равномерный и агрегативный тип пространственного распределения особей в насаждении, а для количественной оценки пространственного распределения используют многочисленные индексы агрегации (Песенко, 1982).
При более детальном анализе пространственной структуры популяций лесных насекомых используется подход, заключающийся в изучении пространственной сопряженности очагов массового размножения лесных насекомых с ландшафтными особенностями территорий и составом насаждений. Так, исследования на реконструированных данных по вспышкам лесного кольчатого шелкопряда (Malacosoma disstria Hbn.) в течение почти 100 лет (с 1928 по 2006 гг.) показали, что вспышки численности будут проявлять большую синхронность и интенсивность в районах, связанных с более высокой численностью деревьев-хозяев (Sutton, Tardif, 2007; Cooke, Roland, 2007; Cooke et al., 2012; Charbonneau et al., 2012; Robert et al., 2020). Вспышки были более синхронными и более цикличными в пределах управляемых зон, содержащих большее относительное обилие осины и других лиственных пород деревьев-хозяев, по сравнению с дикой природой, где преобладают хвойные породы. Тем не менее, также наблюдалась асинхронная динамика вспышек на исследуемой территории, так что корреляции с более медленными переменными лесных ландшафтов сильно варьировали между вспышками, и сила взаимосвязи между ландшафтными переменными и моделями вспышек этого вида сильно менялась со временем – результат, который может объяснить, почему краткосрочные исследования давали противоречащие друг другу выводы.
Работы в этом направлении, выполненные, в частности, для елового почкоеда (Choristoneura fumiferana Clem. (Robert et al., 2018)), важны для выявления зон риска вспышек разных видов, однако в них не рассматриваются особенности развития и пространственной динамики отдельных вспышек. Математический аппарат, используемый при таком анализе, – многомерный регрессионный анализ и автокорреляционный анализ – достаточно хорошо разработан.
В последнее время для анализа пространственного распределения видов насекомых используется подход, который объединяет теорию сетей и сложных систем для моделирования сложных развивающихся пространственных сетей. В частности, такой подход реализован на примере жука-короеда изумрудного ясеня с использованием наборов геопространственных данных в Онтарио, Канаде (Anderson, Dragicevic, 2018).
Второе направление исследований при изучении пространственной динамики вспышек лесных насекомых – анализ пространственных кросс-корреляций между вспышками разных видов на территории (Barbour, 1990). Такой подход позволяет выявить синхронность и временные сдвиги между вспышками разных видов на одной территории или одного вида на разных территориях. Техника расчетов также очень хорошо развита, и используются методы корреляционного и спектрального анализа временных рядов. Проблема заключается в получении информации о достаточно длинных временных рядах динамики численности популяций лесных насекомых.
Еще один подход к анализу пространственно-временной динамики развития очагов массового размножения лесных насекомых состоит в использовании данных о площадях очагов в разные годы и переходе от представления данных в пространстве к временным рядам суммарной динамики площадей очагов. При таком подходе нет необходимости в определении координат очагов во времени, достаточно получить данные по площадям повреждений, имеющимся в распоряжении служб охраны лесов, а математический аппарат, используемый при анализе, сводится к стандартным методам статистического анализа временных рядов (Soukhovolsky et al., 2022).
Наиболее сложным по необходимости сбора данных и методам анализа этих данных является пространственный анализ динамики развития очагов массового размножения лесных насекомых, включающий анализ форм границ очагов, скорости распространения очагов в лесных массивах и связи этих показателей с ландшафтными характеристиками, структурой и породным составом насаждений. В течение многих лет получить подобную информацию с точными координатами динамически изменяющихся очагов массового размножения было крайне трудно технически. Теоретические исследования по сходной тематике не позволяли учитывать конкретные особенности изучаемых экосистем (Колмогоров и др., 1937; Fisher, 1937; Свирежев, 1987). Однако в настоящее время информацию по таким характеристикам очагов стало возможным получить, используя GPS-позиционирование, данные со спутников и беспилотных летательных аппаратов (Wolter et al., 2008; Князева и др., 2019). И в связи с появлением доступных данных возникла задача разработки моделей, позволяющих анализировать динамику развития очагов и изменения их формы во времени. Математический аппарат, используемый для решения этой задачи, включает уравнения в частных производных, фрактальный анализ, методы, заимствованные из статистической физики.
Настоящая работа посвящена обсуждению возможностей описания динамики развития очагов массового размножения лесных насекомых на разных пространственных масштабах.
ОБЪЕКТЫ И МЕТОДИКА
При построении и верификации моделей будут использоваться данные учетов численности сибирского шелкопряда и заселенности деревьев в очагах его массового размножения и дистанционные данные по площадям и форме очагов в районах Красноярского края в ходе вспышки массового размножения сибирского шелкопряда в 2015–2019 гг.
Вспышки массового размножения лесных насекомых связаны с формированием участков поврежденных деревьев – очагов массового размножения. Площадь очага зависит от численности популяции вредителей, состояния деревьев, ландшафтной структуры территории (Исаев и др., 2001). Суммарная площадь очагов и скорость роста этой площади характеризует риски повреждения насаждений, и для оценки уровня воздействия насекомых необходимы прогнозы развития очагов.
Формирование очагов связано с миграцией особей. Перемещение насекомых и постоянная смена их местообитания вызваны необходимостью поиска корма, так как во многих случаях кормовые ресурсы в очагах уничтожаются полностью и необходимо искать новые насаждения с доступным кормовым ресурсом.
В настоящей работе свойства очагов массового размножения рассмотрены на двух пространственных масштабах:
1. На микропространственном масштабе проанализировано распределение особей по кормовым объектам в границах локальной территории или насаждения.
2. На макропространственном масштабе рассмотрены показатели изъятия фотосинтезирующего аппарата (листьев или хвои) по всей территории очага, включая первичные, вторичные и миграционные очаги.
На микропространственном масштабе в качестве характеристик очага используются показатели средней плотности гусениц Х в расчете на одно дерево в локальном насаждении и доля А учетных единиц (например, деревьев), на которых были обнаружены насекомые. Эти данные получают в ходе учетов численности насекомых на разных фазах градационного цикла популяций. Для описания очага в данном случае используется параметр порядка q – доля учетных единиц, на которых при учетах не было обнаружено насекомых, и рассматривается связь между средней плотностью Х насекомых на участке и параметром порядка q. Для описания этой связи в настоящей работе использована модель распределения особей на кормовых деревьях как фазового перехода второго рода (Суховольский и др., 2005; Суховольский и др., 2008; Isaev et al., 2017).
Согласно этой модели связь между квадратом доли незаселенных деревьев q2 = (1 – A)2 и средней плотностью популяции выражается следующим образом:
(1)
${{q}^{2}} = \left\{ \begin{gathered} 0,\,\,\,\,X \geqslant {{X}_{c}} \hfill \\ a - bX,\,\,\,\,0 \leqslant X < {{X}_{c}} \hfill \\ \end{gathered} \right.,$Если распределение особей по деревьям случайное и описывается пуассоновской моделью, то доля qp деревьев, на которых не встречаются насекомые, зависит от плотности популяции Х и описывается уравнением
Если распределение особей по деревьям групповое, то в этом случае ${{q}^{2}}(X) < {{({{q}_{p}}(X))}^{2}}.$Макропространственные процессы, происходящие в ходе роста очага, включают, во-первых, рост существующего очага и появление новых связных поврежденных участков леса, во-вторых, появление новых несвязных вторичных очагов (островов) и возникновение очага массового размножения с фрактальной размерностью D.
В настоящей работе для описания динамики очага использовался экологический аналог модели Дарси (Суховольский и др., 2008), описывающий течение жидкости в двумерном пространстве сквозь пористые среды (Клеман, Лаврентович, 2007):
где U (x, y) – скорость движения фронта; η – “вязкость” среды; р (x, y) – “давление” популяции на насаждение; b – константа.Если движущая граница раздела остается прямой в горизонтальной плоскости, то ее положение во времени и пространстве задается уравнением $U = xt.$ В этом случае движение границы поддерживается градиентом давления, значение которого находится из уравнения (1):
где р0 – константа.В экологическом аналоге закона Дарси показатель “вязкости” η характеризует величину, пропорциональную времени, необходимому для освоения кормового растения в очаге и вне его (если устойчивость кормового растения к насекомым высокая, то время освоения большое, если устойчивость низкая, то и время освоения незначительное), “давление” ∇р – отношение плотностей насекомых в очаге и вне его.
При большой “вязкости” и малом давлении (“спящий” очаг) при однократном измерении форма границы очага будет характеризоваться фрактальным размером, близким к 1. Когда более “вязкая” среда толкает менее “вязкую”, граница раздела сред устойчива.
Если же вдоль оси y возникает небольшое периодическое возмущение с волновым числом k и амплитудой А(t):
то при малой вязкости и высоком давлении (агрессивная форма вредителя с высокой плотностью популяции) фрактальная размерность границы раздела приблизится к 2, и эта граница будет иметь большое число выступов – “пальцев”. В данном случае возмущение станет неустойчивым, и амплитуда возмущения со временем вырастет. Наибольшие градиенты давления возникнут на вершинах границы. Согласно уравнению Дарси, вершина начнет двигаться быстрее, чем остальные участки границы раздела, и небольшое начальное возмущение разовьется в быстро растущий “вязкий палец”.Ранее экологический аналог модели “вязких пальцев” использовался для описания движения границы очага массового размножения такого относительно малоподвижного вида как непарный шелкопряд (Lymantria dispar L.) в Северной Америке, на территории восточных штатов США, от канадско-американской границы до Аппалачей (Суховольский и др., 2008) (рис. 1).
Рис. 1.
Плотность насекомых по данным мониторинга активности имаго с помощью феромонных ловушек, (штат Огайо, 1998 год, по данным факультета энтомологии технологического института штата Вирджиния). 1–6 – плотность имаго по мере ее убывания.
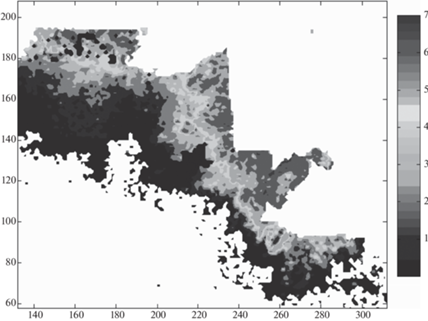
Технология измерения площади и границ очага массового размножения насекомых по дистанционным данным в настоящее время достаточно хорошо разработана. Для оценки развития размеров очагов в этой работе использованы данные дистанционного зондирования очагов сибирского шелкопряда с помощью комплекса Sentinel-2 – группы спутников ДЗЗ Европейского космического агентства. При этом применялся мультиспектральный комплекс с оптическим пространственным разрешением до 10 м. Интервал повторения снимка поверхности (без учета облачности) – пять дней. Регулярная съемка земной поверхности ведется с 2016–2017 гг. Для получения графических изображений и основных статистик (размер, временная динамика, функции распределения вегетативных индексов и т.п.) для изучаемых территорий использовался программный интерфейс доступа: apps.sentinel-hub.com/eo-browser/.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Закономерности микропространственного распределения насекомых на пробных площадях
Пусть отдельная микропространственная единица характеризуется средней плотностью Х популяции вредителей (особей на ученую единицу) и параметром порядка q – долей учетных единиц, на которых в ходе учета не были обнаружены насекомые. Распределение насекомых по деревьям в процессе развития очага массового размножения изменятся от ситуации в стабильно разреженном состоянии, когда плотность популяции очень низка, почти все деревья не заселены и q → 1, и до ситуации, когда плотность популяции настолько высока, что все деревья на пробной площади населены и q = 0. Для описания связи между плотностью популяции Х и заселенностью деревьев в насаждении нами использована модель (1). Для расчетов применены данные по абсолютной и относительной заселенностям сибирского шелкопряда, приведенные в работах В.О. Болдаруева (1969) и Ю.П. Кондакова (1974), и данные учетов Центра защиты леса Красноярского края в ходе вспышки массового размножения в разных лесничествах на территории Енисейского района Красноярского края (рис. 2–4).
Рис. 2.
Связь между квадратом доли незаселенных деревьев q2 и логарифмом плотности Х популяций сибирского шелкопряда в очагах массового размножения в 1953–1969 гг. (Кондаков, 1974). Плотности популяции: 1 – ниже критической плотности освоения Xc, 2 – выше критической плотности освоения Xc.

Рис. 3.
Связь между квадратом доли незаселенных деревьев q2 и логарифмом плотности Х популяции сибирского шелкопряда в очагах массового размножения на территории Красноярского края в 2017 г. Плотности популяции: 1 – ниже критической плотности освоения Xc, 2 – выше критической плотности освоения Xc.
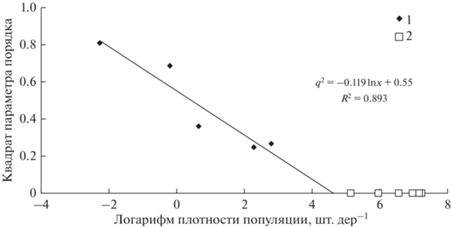
Рис. 4.
Cвязь между квадратом доли незаселенных деревьев q2 на пробных площадях и логарифмом плотности X популяции в очагах массового размножения сибирского шелкопряда на территории Восточной Сибири (Болдаруев, 1969). 1 – пуассоновское распределение; пробные площади: 2 – с неполным заселением деревьев насекомыми, 3 – деревья, полностью заселенные насекомыми.

Как видно из рис. 2–4, распределение гусениц сибирского шелкопряда по деревьям на пробных площадях на разных фазах градации при различной плотности популяции очень хорошо описывается уравнением (1). Этот тип распределения отличается от случайного распределения, описываемого уравнением (2). На рис. 5 приведена разность между квадратом параметра порядка q2 и расчетным значением (qp)2 при пуассоновском распределении особей по деревьям.
Рис. 5.
Разность между пуассоновской моделью распределения гусениц по деревьям и моделью фазового перехода второго рода.

Следовательно, при увеличении плотности популяции сибирского шелкопряда распределение гусениц по деревьям начинает отклоняться от случайного и особи группируются лишь на части деревьев. Такое поведение можно рассматривать с точки зрения увеличения воздействия насекомых на осваиваемые деревья. При этом критическая плотность популяции, по достижении которой все деревья оказываются освоенными насекомыми, для разных фаз вспышек различается. С увеличением доли населенных деревьев возможности регистрации повреждений дистанционными методами возрастают, а по зависимости квадрата доли поврежденных деревьев возможно косвенно оценить плотность популяции на пробной площади.
2. Фрактальная размерность очагов
Форма очага массового размножения, как и любого двумерного ограниченного множества, характеризуется двумя показателями: его площадью и фрактальной размерностью (Федер, 1991). Оценив фрактальную размерность очага, можно рассчитать соотношение плотностей популяции и времени освоения кормового объекта, а по фрактальной размерности границы раздела очага и неповрежденного леса можно оценить характер дальнейшего развития очага. Для расчетов фрактальной размерности очага необходимо оценить площадь очага S и длину L его границ. Площадь S, заключенная внутри контура границы, фрактально зависит от длины границы L (Федер, 1991):
илиВ ходе развития вспышки очаги характеризуются показателями связности. Связным очаг будет в том случае, когда две любые точки внутри очага могут быть соединены линией, не выходящей за пределы границ очага. В противном случае очаг будет несвязным.
Для понимания процессов миграции насекомых необходимо изучение механизмов формирования очагов. Эти механизмы различны на разных фазах вспышек. В начальной фазе вспышки преобладают конденсирующие миграции, когда на участках с наиболее благоприятными для развития вредителей экологическими условиями происходит концентрация численности насекомых за счет притока иммигрантов с окружающих территорий (Исаев и др., 2001). При этом формируются первичные очаги вредителя. После повреждений деревьев в первичных очагах и уничтожения кормовых ресурсов на них насекомые перемещаются во вторичные очаги. Этот процесс можно рассматривать как диффузионный, сопровождающийся перемещением границ очага при сохранении его связности. У насекомых-филлофагов на фазе максимума вспышки наблюдаются адвективные миграции – массовый вылет бабочек из первичных и вторичных очагов и их перемещение воздушными потоками. Вторичные очаги можно разделить на связные и несвязные миграционные. Связные вторичные очаги формируются на границах очагов предыдущих лет, несвязные миграционные очаги формируются на определенных расстояниях от очагов предыдущих сезонов. Появление несвязных очагов можно объяснить мозаичностью породного состава насаждений, где доступные для питания насекомыми участники сосуществуют с участками, на которых отсутствуют кормовые породы насекомых. Формирование таких очагов приводит к появлению мозаичной структуры поврежденных древостоев. С точки зрения статистической теории такие процессы можно рассматривать как полеты Леви (Шредер, 2001).
При затухании очагов вредителей интенсивно развиваются дисперсионные миграции, в ходе которых происходит рассеивание популяций из очагов массового размножения в окружающие лесные массивы, образуются рефугии (станции переживания).
При сильных повреждениях деревьев вредителями, приводящими к повреждениям крон насекомыми – филлофагами или к повреждению луба стволов деревьев насекомыми-ксилофагами, ведущего к последующему усыханию хвои или листового аппарата деревьев, оценку зоны и площадей очагов достаточно просто определить, используя дистанционные методы. В качестве примера на рис. 6 приведен вид со спутника MODIS очагов массового размножения сибирского шелкопряда в Енисейском районе Красноярского края.
Рис. 6.
Пространственная структура очагов массового размножения сибирского шелкопряда (Енисейский район Красноярского края, 2015 г.). Очаги выделены черным цветом.

Как видно из рис. 6, форма и площади миграционных очагов сильно различаются. Каждый новый очаг можно рассматривать как источник формирования нового связного или несвязного очага. К характеристикам очагов относятся показатели общей многолетней динамики повреждений насаждений на всей территории, охваченной вспышкой массового размножения вредителей, и динамики локальных очагов, функцию распределения по площадям очагов, пространственную корреляцию очагов.
Показанный на рис. 6 в масштабе 1 пиксел = 4 × × 4 м очаг массового размножения сибирского шелкопряда несвязен и состоит из большого числа (свыше 200) микроочагов с разной площадью повреждений и длиной границ. Для каждого такого микроочага оценивались площадь и длина границ локальных поврежденных участков леса. Площади и длина границ очагов вычислялись по данным дистанционной съемки с использованием оригинальных вычислительных программ, созданных в среде Borland Delphi.
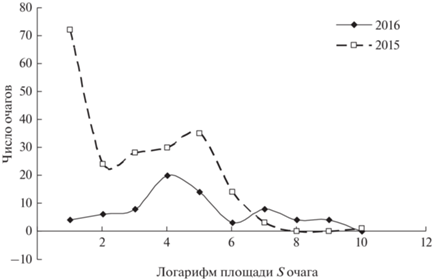
На рис. 7 приведены функции распределения микроочагов по площадям в ходе вспышки массового размножения вредителя на территории Енисейского района Красноярского края в 2015 и 2016 гг.
Рис. 7.
Функции плотности распределения площадей микроочагов вспышки массового размножения сибирского шелкопряда на территории Енисейского района Красноярского края в 2015 и 2016 гг.
При помощи дистанционных данных для каждого микроочага были вычислены его площадь и длина границ. Зная эти величины и основываясь на выражении (7), можно оценить величину фрактальной размерности очагов $D = \frac{2}{\alpha }$ (Шредер, 2001). На рис. 8 и 9 показаны связи между длиной периметра очагов и их площадью, а данные расчетов уравнений типа (7) приведены в табл. 1.
Рис. 8.
Связь между логарифмами длины границ и площади микроочагов на территории Енисейского района Красноярского края (2015 г.)

Рис. 9.
Связь между логарифмами длины границ и площади микроочагов на территории Енисейском района Красноярского края (2016 г.)
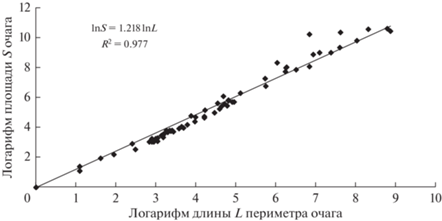
Таблица 1.
Связь между длиной L границ микроочагов и их площадью S в очагах сибирского шелкопряда на территории Красноярского края
| Переменные | Коэффициенты | Стандартная ошибка | t-критерий | p-level |
|---|---|---|---|---|
| Енисейск, 2015 | ||||
| ln s0 | –0.651 | 0.063 | –8.04 | 0.000000 |
| α | 1.235 | 0.016 | 77.81 | 0.000000 |
| adjR2 | 0.981 | |||
| F-критерий Фишера | 6054 | |||
| Фрактальный размер D | 1.62 | |||
| Енисейск, 2016 | ||||
| ln s0 | –0.587 | 0.115 | –5.12 | 0.000003 |
| α | 1.330 | 0.024 | 55.71 | 0.000000 |
| adjR2 | 0.979 | |||
| F-критерий Фишера | 3103.1 | |||
| Фрактальный размер D | 1.50 | |||
| Ирбей, 2019 | ||||
| ln s0 | –0.614 | 0.050 | –12.38 | 0.000000 |
| α | 1.320 | 0.014 | 94.24 | 0.000000 |
| adjR2 | 0.986 | |||
| F-критерий Фишера | 8881.6 | |||
| Фрактальный размер D | 1.51 | |||
Как видно из табл. 1, связь между логарифмами длин границ и площадей микроочагов на территории Енисейского района линейна, согласно t- и F-критериям, коэффициенты уравнения (7) значимы, и уравнение очень хорошо описывает эту связь. Коэффициент детерминации R2 очень близок к 1, и уравнение (7) позволяет учесть свыше 98% дисперсии площадей микроочагов. Фрактальная размерность микроочагов в 2016 г. несколько меньше фрактальной размерности микроочагов в 2015 г. и указывает на то, что вклад диффузионного механизма в формирование очага начинает возрастать.
3. Голографический принцип в оценке свойств очагов массового размножения лесных насекомых
При анализе характеристик очагов массового размножения лесных насекомых возникает вопрос: можно ли сопоставить характеристики очага в целом, такие как численность насекомых, состояние особей, состояние кормовых растений, миграционная активность насекомых, площадь повреждений со свойствами границы очага: ее фрактальной размерностью и динамикой. Такое сопоставление по аналогии с работами по космологии (Сасскинд, 2013) можно охарактеризовать как голографический принцип: свойства очага массового размножения кодируются свойствами его границы.
При фрактальной размерности границы очага, существенно большей 1, эта граница будет иметь большое число “пальцев” – выступов, что характерно для очага агрессивного вида в неоднородном по устойчивости деревьев древостое. Очаги с малой площадью, характеризуемые отсутствием “вязких пальцев” на их границах, можно рассматривать как “спящие”, и риск расширения их невелик.
Оценка и прогноз динамики площадей неустойчивых или уже поврежденных насаждений открывают возможность расчета моделей динамики развития очагов, характеристик как насекомых-вредителей, так и состояния кормовых древесных растений.
Для оценки параметров модели перспективно использование данных дистанционного зондирования. На рис. 10 приведено отредактированное изображение фрагмента очага массового размножения сибирского шелкопряда в Ирбейском районе Красноярского края в 2019 г. полученное с использованием спутника Sentinel-2.
Рис. 10.
Граница очага массового размножения сибирского шелкопряда в Ирбейском районе Красноярского края в 2019 г., полученная с использованием спутника Sentinel-2.

Как видно из рис. 10, очаг массового размножения несвязен и можно выделить зону полного повреждения деревьев, зону неповрежденных деревьев и зону локальных очагов, которую по терминологии теории перколяции будем называть “решеточными зверями” (lattice animals) (Тарасевич, 2002). Точно так же, как и для очагов в Енисейском районе, была вычислена связь между длиной границ и площадью “решеточных зверей” и определена их фрактальная размерность (рис. 11 и табл. 1).
Рис. 11.
Связь между логарифмами длины и площади микроочагов в Ирбейском районе Красноярского края (2019 г.)

Как видно из рис. 11, связь между длиной периметра и площадью очагов в ходе вспышки массового размножения сибирского шелкопряда на территории Ирбейского района имеет ту же форму, что и связи на территории Енисейского района. Как следует из табл. 1, фрактальная размерность очагов на территории Ирбейского района та же, что и фрактальная размерность очагов на территории Енисейского района.
Для оценки связей между характеристиками очагов и их границ необходимо ввести количественные характеристики границ. Для этого, выделив на снимке (рис. 10) границу, запишем регрессионное уравнение для координат X и Y границы очага Y = K – PХ (рис. 12).
Рис. 12.
Расчет связи координат X и Y границы локального очага массового размножения сибирского шелкопряда в Ирбейском районе.
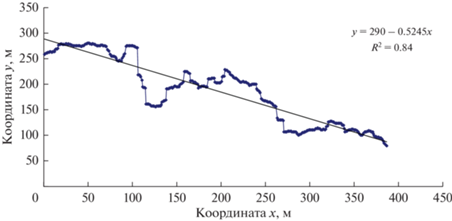
После такого расчета координаты границы очага были трансформированы с помощью преобразования $Z = Y(X) - (K - PX).$ Результаты этого преобразования отражены на рис. 13.
Рис. 13.
“Вязкие пальцы” на границе очага массового размножения после трансформации координат границы очага массового размножения сибирского шелкопряда в Ирбейском районе в 2019 г.

Как видно из рис. 13, на границе очага формируются выступы и впадины – “вязкие пальцы”. В качестве интегральных характеристик границы будем использовать мощность W спектра длины границы, рассматриваемого как аналог временного ряда, частоту fmax спектра (при максимальном значении частоты, равной частоте Найквиста fN =1/2 спектра, характеризующего максимальное значение спектральной мощности (Андерсон, 1976), и период λ “вязких пальцев” – величину, обратную значению fmax. Эти значения приведены в табл. 2.
4. Дистанционные расчеты динамики площадей очага в течение сезона
Для анализа связи между площадью очага и характеристиками его границ будем использовать данные дистанционного зондирования очага в Ирбейском районе с помощью спутника Sentinel-2. На рис. 14 представлено распределение значений NDVI по пикселам 10 × 10 м на пробной площади в Ирбейском районе в течение сезона 2019 г., частично поврежденной сибирским шелкопрядом.
Рис. 14.
Повреждения насаждения сибирским шелкопрядом (Ирбейский район Красноярского края, с марта по сентябрь 2019 г.). Дистанционное зондирование (спутник Sentinel-2). 1 – 24.04.2019; 2 – 30.06.2019; 3 – 30.10. 2019.
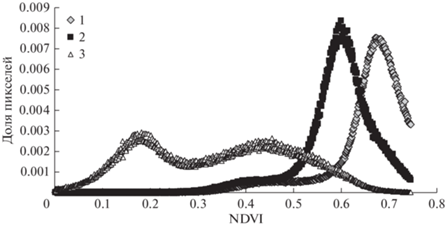
Сравнивая кривые на рис. 14, сделаем вывод, что повреждение насаждения сибирским шелкопрядом ведет к изменению формы кривой NDVI в течение сезона. Используя эти данные, можно оценить скорость объедания насаждения вредителем. На рис. 15 представлена кривая изменения доли неповрежденной части в насаждении (доля пикселов с NDVI > 0.8) в течение сезона 2019 г. в очаге массового размножения сибирского шелкопряда в Ирбейском районе Красноярского края.
Рис. 15.
Изменение доли неповрежденной части в насаждении в течение сезона 2019 г. в очаге массового размножения сибирского шелкопряда в Ирбейском районе Красноярского края.
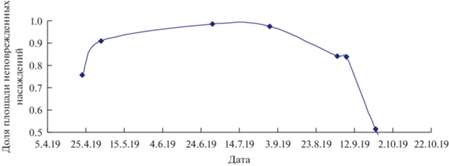
По кривой на рис. 15 вычисляем среднюю скорость роста площади очага, составляющую 0.0075 пикселов/сутки. Данные по средней скорости роста площади очага можно сопоставить с частотой максимума спектра, fmax = 0.0077.
ЗАКЛЮЧЕНИЕ
Настоящую работу можно рассматривать как один из возможных подходов к моделированию динамики очагов массового размножения лесных насекомых. В отличие от достаточно хорошо исследованных точечных моделей динамики численности лесных насекомых, для оценки и прогноза процессов роста очагов массового размножения еще не существует общепризнанных моделей.
Во многом это связано с неопределенностью описания механизмов формирования очага. Так, в классической модели Колмогорова–Петровского–Пискунова (первоначально разработанной для описания захвата территории некоторым геном) территория области заселения по определению предполагается связной. Точно так же связна освоенная территория и в диффузионной модели. Однако наблюдения показывают, что реальные очаги несвязны и необходимы модели, учитывающие эти особенности процесса захвата территории вредителем. Еще один неясный момент в теории роста очагов массового размножения насекомых – связь характеристик скорости роста с формой границ очага и видом “вязких пальцев” на границе. Первичный анализ, проведенный в настоящей работе, указывает на возможность существования подобной связи, однако для точной оценки этой связи необходимы дополнительные исследования.
Представленные в настоящей работе данные указывают на перспективность использования дистанционных данных для описания и моделирования динамики очага массового размножения. Важным вопросом является вопрос о существовании критического значения площади очага и его критической фрактальной размерности. Если такие характеристики существуют, то при значениях ниже критических развития очага не будет.
По представленным в настоящей работе данным, исследованные очаги одного вида – сибирского шелкопряда характеризуются фрактальной размерностью D = 1.5–1.6, но неясно, действительно ли существует видоспецифичная фрактальная размерность очагов отдельных видов насекомых или же эта фрактальная размерность определяется характеристиками ландшафта и пространственным распределением древесных пород в насаждениях.
Представляется, что дальнейшие исследования позволят уточнить возможные механизмы развития очагов массового размножения лесных насекомых и предложить надежные модели, которые позволили бы прогнозировать динамику очагов и допустимые повреждения насаждений, что весьма важно при планировании задач защиты леса от насекомых-вредителей.
Список литературы
Андерсон Т. Статистический анализ временных рядов. М.: Мир, 1976. 755 с.
Болдаруев В.О. Динамика численности сибирского шелкопряда и его паразитов. Улан-Удэ: Бурят. книж. из-во, 1969. 162 с.
Исаев А.С., Хлебопрос Р.Г., Кондаков Ю.П., Недорезов Л.В., Киселев В.В., Суховольский В.Г. Популяционная динамика лесных насекомых М.: Наука, 2001. 374 с.
Клеман М., Лаврентович О.Д. Основы физики частично упорядоченных сред. М.: Физматлит, 2007. 680 с.
Князева С.В., Королева Н.В., Эйдлина С.П., Сочилова Е.Н. Оценка состояния растительности в очаге массового размножения сибирского шелкопряда по спутниковым данным // Лесоведение. 2019. № 5. С. 385–398.
Колмогоров А.Н., Петровский И.Г., Пискунов Н.С. Исследование уравнения диффузии, соединенной с возрастанием вещества, и его применение к одной биологической проблеме // Бюллетень МГУ. Серия А. Математика и механика. 1937. Т. 1. № 6. С. 1–25.
Кондаков Ю.П. Закономерности массовых размножений сибирского шелкопряда // Экология популяций лесных животных Сибири. Новосибирск: Наука, 1974. С. 206–265.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
Сасскинд Л. Битва при черной дыре. СПб.: Питер, 2013. 448 с.
Свирежев Ю.М. Нелинейные волны, диссипативные структуры и катастрофы в экологии. М.: Наука, 1987. 368 с.
Суховольский В.Г., Пальникова Е.Н., Тарасова О.В., Карлюк А.Ю. Модель вспышки массового размножения лесных насекомых как фазового перехода второго рода // Доклады Академии наук. 2005. Т. 403. № 4. С. 551–553.
Суховольский В.Г., Тарасова О.В., Ковалев А.В. Моделирование эмиссии углерода в ходе массового размножения лесных насекомых // Лесоведение. 2006. № 5. С. 22–28.
Суховольский В.Г., Исхаков Т.Р., Тарасова О.В. Оптимизационные модели межпопуляционных взаимодействий. Новосибирск: Наука, 2008. 162 с.
Тарасевич Ю.Ю. Перколяция: теория, приложения, алгоритмы. М.: Едиториал УРСС, 2002. 112 с.
Тарасова О.В., Калашникова И.И., Кузнецова В.В. Энергетический баланс потребления корма насекомыми-филлофагами: оптимизационная модель // Сибирский лесной журн. 2015. № 3. С. 83–92.
Федер Е. Фракталы. М.: Мир, 1991. 261 с.
Шредер М. Фракталы, хаос, степенные законы. Ижевск: РХД, 2001. 528 с.
Anderson T.M., Dragicevic S. Network-agent based model for simulating the dynamic spatial network structure of complex ecological systems // Ecological Modelling. 2018. V. 389. P. 19–32.
Barbour D.A. Synchronous fluctuations in spatially separated populations of cyclic forest insects // Population dynamics of forest insects. Intercept Limited, Andover, UK. 1990. P. 339–346.
Charbonneau D., Lorenzetti F., Doyon F., Mauffette Y. The influence of stand and landscape characteristics on forest tent caterpillar (Malacosoma disstria) defoliation dynamics: the case of the 1999–2002 outbreak in northwestern Quebec // Canadian J. Forest Research. 2012. V. 42. P. 1827–1836.
Cooke B.J., MacQuarrie C.J.K., Lorenzetti F. The dynamics of forest tent caterpillar outbreaks across east-central Canada // Ecography. 2012. V. 35. P. 422–435.
Cooke B.J., Roland J. Trembling aspen responses to drought and defoliation by forest tent caterpillar and reconstruction of recent outbreaks in Ontario // Canadian J. Forest Research. 2007. V. 37. № 9. P. 1586–1598.
Fisher R.A. The Wave of Advance of Advantageous Genes // Annals of Eugenics. 1937. V. 7. P. 355–369.
Isaev A.S, Soukhovolsky V.G., Tarasova O.V., Palnikova E.N., Kovalev A.V. Forest Insect Population Dynamics, Outbreaks and Global Warming Effects. Wiley, N.Y., 2017. 298 p.
Robert L.E., Sturtevant B.R., Cooke B.J., James P.M., Fortin M.J., Townsend P.A., Wolter P.T., Kneeshaw D. Landscape host abundance and configuration regulate periodic outbreak behavior in spruce budworm (Choristoneura fumiferana Clem.) // Ecography. 2018. V. 41. P. 1556–1571.
Robert L.-E., Sturtevant B.R., Kneeshaw D., James P.M.A., Fortin M.-J., Wolter P.T., Townsend P.A., and Cooke B.J. Forest landscape structure influences the cyclic-eruptive spatial dynamics of forest tent caterpillar outbreaks // Ecosphere. 2020. V. 11(8):e03096. https://doi.org/10.1002/ecs2.3096
Soukhovolsky V., Ivanova Y. Modeling Production Processes in Forest Stands: An Adaptation of the Solow Growth Model // Forests. 2018. № 9. P. 391–403.
Soukhovolsky V., Kovalev A., Tarasova O. et al. Wind Damage and Temperature Effect on Tree Mortality Caused by Ips typographus L.: Phase Transition Model // Forests. 2022. V. 13. № 2. 180 p. https://doi.org/10.3390/f13020180
Sutton A., Tardif J. Dendrochronological reconstruction of forest tent caterpillar outbreaks in time and space, western Manitoba, Canada // Canadian J. Forest Research. 2007. V. 37. № 9. P. 1643–1657.
Wolter P.T., Townsend P.A., Sturtevant B.R., Kingdon C.C. Remote sensing of the distribution and abundance of host species for spruce budworm in Northern Minnesota and Ontario // Remote Sensing of Environment. 2008. V. 112. № 10. P. 3971–3982.
apps.sentinel-hub.com/eo-browser
Дополнительные материалы отсутствуют.