

Зоологический журнал, 2020, T. 99, № 5, стр. 569-585
Фауна крупных млекопитающих центральной части Восточно-Европейской равнины в голоцене
О. П. Бачура a, *, П. А. Косинцев a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: olga@ipae.uran.ru
** E-mail: kpa@ipae.uran.ru
Поступила в редакцию 23.08.2019
После доработки 05.09.2019
Принята к публикации 19.09.2019
Аннотация
На основании данных из многочисленных археологических памятников описана история фауны крупных млекопитающих в центральной части Восточно-Европейской равнины в голоцене. На протяжении всего голоцена во всем регионе элементы фауны таежного териокомплекса носили зональный характер. Элементы фауны широколиственных лесов расселялись здесь вслед за распространением неморальной растительности. Выделяются северный и южный варианты териофауны. В северной части в состав фауны входил соболь, в южной – благородный олень. В северной части формирование состава фауны происходило в два этапа. Ядро фауны сформировалось к началу атлантического периода. Окончательный состав сформировался к началу суббореального периода, в результате расселения ряда видов из южной части к северу. Состав териофауны в южной части региона сформировался к началу атлантического периода и до современности оставался относительно стабильным. Современный облик на всей территории региона фауна приобрела в конце позднего голоцена, когда под влиянием антропогенных и климатических факторов сократился ареал у одних видов и расширился у других.
Формирование современной фауны млекопитающих отдельных регионов представляет значительный интерес, так как позволяет выявить общие и специфические закономерности фауногенеза. Не очевидно, что эти закономерности будут одинаковыми для крупных и для мелких млекопитающих. Методика анализа истории териофауны в целом, включая и крупных и мелких млекопитающих, не разработана, поэтому эти группы рассматриваются по отдельности. К настоящему времени с разной степенью детальности изучена история формирования современной фауны крупных млекопитающих Прибалтики (Паавер, 1965), Молдавии (Давид, 1982), Волго-Уралья (Гасилин, 2005; Косинцев, Гасилин, 2005), Северного (Bachura, Kosintsev, 2007) и Южного (Kosintsev, Bachura, 2013) Урала, юго-востока Западной Сибири (Девяшин, Косинцев, 2013), Прибайкалья (Пластеева, Клементьев, 2018), юга Приморья (Гасилин, 2013). Но для большинства районов Северной Евразии таких исследований нет. Основная причина – это отсутствие достаточного количества данных.
Центральная часть Восточно-Европейской равнины относится к регионам, в которых начинались первые археологические работы в России. Одно из первых археозоологических исследований в России так же проведено по материалам из этого района (Иностранцев, 1882; Боголюбский, 1928). Позднее были изучены костные остатки еще из нескольких поселений этого региона (Андреева, 1940; Алексашина, 1950; Бибикова, 1950). Но это были единичные исследования. И только Цалкин (1956; 1962; 1971) начал регулярное изучение археозоологических материалов центральной части Восточно-Европейской равнины. Позднее костные остатки животных из археологических памятников изучал ряд исследователей (Андреева, 1972; 1974; 1991; Верещагин, Русаков, 1979; Данильченко, 1984; Кириллова, 1992; 1998). В результате, в настоящее время накоплен значительный объем данных по истории крупных млекопитающих региона, которые охватывают весь голоцен.
Цель работы: рассмотреть распространение крупных млекопитающих в течение голоцена на территории центральной части Восточно-Европейской равнины и установить время формирования современной фауны.
МАТЕРИАЛ И МЕТОДЫ
Район исследований включает территорию примерно между 62° и 54° с.ш., 29° и 45° в.д. В административном плане он включает: Псковскую, Ленинградскую, Новгородскую области, юго-западную часть Архангельской обл., Вологодскую, Ярославскую, Тверскую, Ивановскую, Владимирскую, Московскую, Смоленскую, Калужскую, Рязанскую и Нижегородскую области (рис. 1). В настоящее время на этой территории расположена зона темнохвойной тайги с участками сосновых лесов, южная граница которой проходит в западной части по 59° с.ш., а в восточной – по 56° с.ш. Южнее до границы с Брянской, Орловской и Липецкой областями находится зона смешанных широколиственно-темнохвойных лесов с сосново-широколиственных участками (Мильков, 1977).
Рис. 1.
Расположение голоценовых археологических памятников на территории Восточно-Европейской равнины: 1 – Анашкино; 2 – Березовая Слободка; 3 – Бологое; 4 – Боровский курган; 5 – Варос; 6 – Вашана; 7 – Вёкса-3; 8 – Веретье-1; 9 – Весь-5; 10 – Вишенка-3; 11 – Владычинская береговая; 12 – Воймежное; 13 – Городище; 14 – Дубокрай-5; 15 – Дютьково; 16 – Звенигородская стоянка; 17 – Дьяковское городище; 18 – Старшее Каширское; 19 – Троицкое; 20 – Настасьино; 21 – Жидиховское болото; 22 – Озерки-5; 17; 23 – Ивановское-2, 3, 4, 7; 24 – Караваиха; 25 – Круглица; 26 – Крутик; 27 – Култино-3; 28 – Мамонтово; 29 – Маслово болото 2, 4; 30 – Минино; 31 – Минино-2; 32 – Модлона; 33 – Наумово; 34 – Усвяты 4; 35 – Дяздица; 36 – Нижнее Веретье; 37 – Нижний Новгород; 38 – Городок; 39 – Николо-Ленивец; 40 – Никольская правая; 41 – Новгород; 42 – Огубское; 43 – Нушполы 2; 44 – Окаемово-5; 45 – Замостье-2; 46 – Пекуновское городище; 47 – Переяславль-Рязанский; 48 – Старая Рязань; 49 – Песчаница; 50 – Попово; 51 – Погостище 1, 14, 15; 52 – Подолье-1; 53 – Ладожская стоянка; 54 – Старая Ладога; 55 – Осыно; 56 – Покровский бугор; 57 – Псков; 58 – Ростиславльское городище; 59 – Сахтыш 2, 9; 60 – Сертея 2, 8, 10, 11; 61 – Рудня Сертейская; 62 – Свинуховское; 63 – Смоленск; 64 – Становое-4; 65 – Старый Выборг; 66 – Стрелка-1; 67 – Сулать; 68 – Тверь; 69 – Черная гора; 70 – Шагара-2; 71 – Языково-1; 72 – Ярославль; 73 – Селецкое селище; 74 – Щербинское городище.
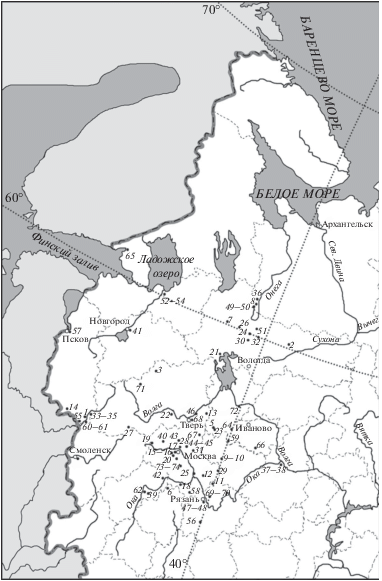
В работе использованы опубликованные и оригинальные данные о видовом составе 108 фаун крупных млекопитающих из 87 археологических памятников (рис. 1) (Боголюбский, 1928; Городцов, 1933; Андреева, 1940; 1972; 1974; 1991; Третьяков, 1941; Алексашина, 1950; Бибикова, 1950; Цалкин, 1956; 1961; 1962; 1971; Ермолова, 1976; Верещагин, Русаков, 1979; Данильченко, 1984; Кириллова, 1992; 1998; Данильченко, Карху, 1997; Жилин, 1997; 2004; Ошибкина, 1997; Калiноўскi, 1998; Кириллова, Емельянов, 1999; Кольцов, Жилин, 1999; Шэ и др., 2001; Гадзяцкая, Крайнов, 2002; Карху и др., 2003; Кузьмина, 2003; Крылович, Савинецкий, 2005; Крылович, 2006; Иванечева, 2007; Дмитриева и др., 2008; Антипина, 2009; 2013; Лапшин, 2009; Савинецкий, Крылович, 2009; Короткевич и др., 2010; Костылёва, Уткин, 2010; Лисицын, 2011; Саблин и др., 2011; Кренке, 2011; Кренке и др., 2012; Добровольская, 2012; Косорукова, 2012; Энговатова и др., 2012; Антипина, Грибов, 2013; Асылгараева, 2013; Зиновьев, 2014; Zhilin, 2014; Галимова и др., 2015; Недомолкина и др., 2015; Сердюк и др., 2018). Из этих местонахождений описано более 50 тыс. костных остатков млекопитающих (рис. 2).
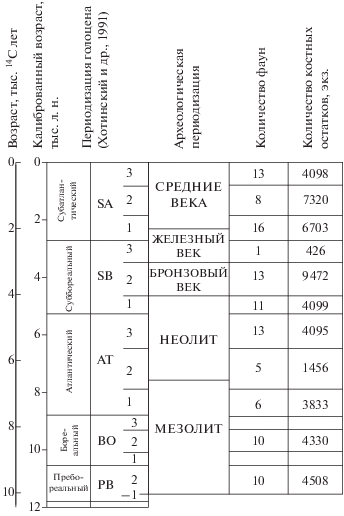
Все фауны датированы по археологическим материалам. Ряд памятников – многослойные, содержащие материалы разных временны́х периодов. Регион очень хорошо изучен археологически (Формозов, 1961; Седов, 1982; Археология СССР, 1987а; 1987б; 1989; Археология, 1996). В последние годы для всех археологических периодов региона получены серии радиоуглеродных дат (Археология, 1996; Тимофеев и др., 2004; Zaretskaya et al., 2005; Hartz et al., 2010; Жилин, 2014; Лозовский и др., 2014; Зайцева и др., 2016; Piezonka et al., 2016). Это позволило надежно установить последовательность, хронологические позиции археологических культур и привязать их к климатостратиграфической шкале голоцена (рис. 2). Хроностратиграфические подразделения голоцена приведены согласно Хотинскому с соавторами (1991). В работе использованы калиброванные даты. Фаунистические данные получены для всех археологических периодов – от мезолита до позднего средневековья (рис. 2). Наиболее многочисленные выборки имеются для этапа 2 пребореального периода, этапа 2 бореального периода, атлантического периода, этапов 1 и 2 суббореального периода и субатлантического периода (рис. 2). Это позволяет дать характеристики фаунам крупных млекопитающих для всех периодов голоцена.
Все местонахождения, которые были включены в анализ, представляют собой поселения древнего человека. Практически все костные остатки попали в археологические местонахождения в результате промысловой деятельности человека. Таким образом, использованный материал является тафономически однородным. Это снижает вероятность получения существенно смещенных оценок при его анализе. Фауны отдельных местонахождений характеризуют промысловую териофауну. Последняя не в полной мере совпадает с понятием “фауна крупных млекопитающих”, так как включает виды, относимые к мелким млекопитающим (Формозов, 1948). К последним относятся обыкновенный еж (Erinaceus europaeus L. 1758), выхухоль (Desmana moschata L. 1758), белка (Sciurus vulgaris L. 1758), ласка (Mustela nivalis L. 1758) и горностай (Mustela erminea L. 1758). Остатки этих видов, а также росомахи (Gulo gulo L. 1758), рыси (Lynx lynx L. 1758), темного хоря (Mustela putorius L. 1758) и европейской норки (Mustela lutreola L. 1761) найдены в ограниченном количестве местонахождений. Это не позволяет охарактеризовать их ареалы в голоцене, но позволяет определить их наличие или отсутствие в составе фаун.
РЕЗУЛЬТАТЫ
Анализ видовых списков фауны крупных млекопитающих в голоценовых местонахождениях показал следующее. На рассматриваемой территории обитают c начала голоцена и до настоящего времени: обыкновенный еж, заяц-беляк (Lepus timidus L. 1758), белка, бобр (Castor fiber L. 1758), волк (Canis lupus L. 1758), лисица (Vulpes vulpes L. 1758), бурый медведь (Ursus arctos L. 1758), лесная куница (Martes martes L. 1758), европейский барсук (Meles meles L. 1758), горностай, ласка, европейская норка, выдра (Lutra lutra L. 1758), лось (Alces alces L. 1758) и северный олень (Rangifer tarandus L. 1758) (табл. 1). Такие виды как обыкновенный еж, заяц-беляк, бобр, волк, лисица, бурый медведь, лесная куница, горностай, ласка, рысь, лось и северный олень с середины пребореального периода были распространены вплоть до 61° с.ш. Остатки барсука, норки и выдры пребореального–конца атлантического периодов известны только из местонахождений, расположенных южнее 58° с.ш. В местонахождениях суббореального и субатлантического периодов остатки этих видов найдены вплоть до 60° с.ш. Вероятно, с пребореального и до конца атлантического периодов северная граница ареалов этих видов в регионе не выходила за пределы 58° с.ш. Их расселение к северу произошло только в начале суббореального периода. В настоящее время европейский барсук и европейская норка распространены до 65° с.ш. Ареал выдры в настоящее время простирается до северного побережья Кольского п-ова (Гептнер и др., 1967).
Таблица 1.
Млекопитающие центра Восточно-Европейской равнины в конце позднего плейстоцена и в голоцене
| Вид | Предпочитаемые местообитания | YD-PBO-1 | PBO2 | BO1 | BO2 | AT1 | AT2 | AT3 | SB1 | SB2 | SB3 | SA1 | SA2 | SA3 | XIX век |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sciurus vulgaris | Таежные | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Marmota bobak | Степные | + | – | – | – | – | – | – | – | – | – | – | – | + | – |
| Castor fiber | Околоводные биотопы в лесной зоне | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Lepus timidus | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Lepus europaeus | Лесостепные | – | – | – | – | – | – | – | – | – | – | – | + | + | |
| Erinaceus europaeus | Таежные | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Desmana moschata | Околоводные биотопы в лесной зоне | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Lynx lynx | Таежные | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Canis lupus | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Vulpes lagopus | Тундровые | + | – | +** | – | – | – | – | – | – | – | – | +** | – | – |
| Vulpes vulpes | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Ursus arctos | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Lutra lutra | Околоводные биотопы в лесной зоне | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Gulo gulo | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Martes foina | Лесостепные | – | – | – | – | – | +? | – | – | – | – | – | – | – | |
| Martes martes | Лесные | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Martes zibellina* | Таежные | ? | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Meles meles | Лесостепные | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Mustela erminea | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Mustela lutreola | Околоводные биотопы в лесной зоне | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Mustela nivalis | Широкий спектр | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Mustela putorius | Широколиственные леса | – | – | – | + | + | + | + | + | + | + | + | + | + | |
| Equus ferus | Лесостепные | + | – | – | – | – | – | – | + | – | – | – | – | – | – |
| Sus scrofa | широколиственные леса, лесостепные | – | ? | + | + | + | + | + | + | + | + | + | + | + | |
| Alces alces | Таежные | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Capreolus capreolus | Лесостепные | – | ? | + | + | + | + | + | + | + | + | + | + | + | |
| Rangifer tarandus | Тундровые и таежные | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Cervus elaphus | Широколиственные леса, лесостепные | – | – | – | + | + | + | + | + | + | + | + | – | – | |
| Bos primigenius | Лесостепные | – | – | – | – | – | + | + | + | – | – | – | – | – | |
| Bison priscus | Перигляциальные степи позднего плейстоцена | + | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Mammuthus primigenius | Перигляциальные степи позднего плейстоцена | + | – | – | – | – | – | – | – | – | – | – | – | – | – |
Остатки выхухоли найдены только в слоях пребореального периода местонахождений Становое-4 и Ивановское-7 (Кириллова, 1998; Жилин, 2004; Zhilin, 2014). По данным Кирикова (1966), выхухоль была распространена в регионе до XIX века. Вероятно, этот вид обитал здесь в течение всего голоцена.
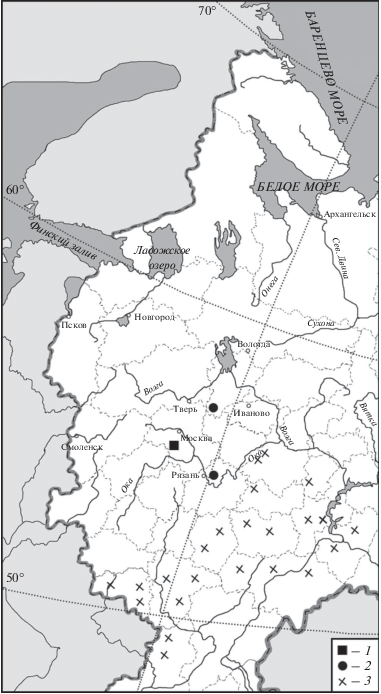
Немногочисленные остатки степного сурка (Marmota bobak Müller 1776) в центральной части Восточно-Европейской равнины найдены в нескольких голоценовых местонахождениях. В XVIII– XIX веках сурки обитали в Орловской, Тамбовской, Тульской и Рязанской губерниях (Кириков, 1980) (рис. 3). В настоящее время северная граница ареала сурка не выходит за пределы Тамбовской и Нижегородской областей (рис. 3). Одна находка костей сурка происходит из Щербинского городища в Московской обл. Согласно радиоуглеродной дате возраст этой находки 1610 лет назад (Савинецкий, 1993), т.е. она относится к концу субатлантического периода. Две другие находки сурка происходят из местонахождений середины суббореального периода Шагара-2 в Рязанской обл. и Ивановское-2 в Ярославской об. (Кириллова, Емельянов, 1999; Костылёва, Уткин, 2010; рис. 3). Радиоуглеродных дат для этих двух находок нет. Четвертая находка обнаружена при раскопках могильника так же суббореального периода во Владимирском ополье (Гроссет и др., 1965). Согласно радиоуглеродной дате возраст этой находки 29130 лет назад (Савинецкий, 1993). 4000–3500 тыс. лет назад в регионе не было подходящих условий для обитания сурка. Скорее всего, кости сурка из Шагары-2 и Ивановского-2 относятся к позднему плейстоцену либо к началу нашей эры.
Рис. 3.
Находки Marmota bobak в голоцене в центральной части Восточно-Европейской равнины в голоцене: 1 – находки субатлантического периода, 2 – недатированные находки из голоценовых памятников, 3 – современные колонии.
На настоящий момент нет возможности решить вопрос о времени заселения региона зайцем-русаком (Lepus europaeus Pallas 1778). В большинстве случаев остатки зайца определены без видовой принадлежности. По данным Кирикова (1966) заяц-русак уже в первой половине XVIII века встречался в окрестностях Петербурга. На территории Валдайского поозерья заяц-русак известен с X века (Тишков, 2013). В настоящее время ареал зайца-русака полностью охватывает рассматриваемый регион вплоть до низовьев реки Онеги (Громов, Ербаева, 1995).
Немногочисленные остатки росомахи и рыси не позволяют реконструировать их ареалы на данной территории. Вероятно, эти животные населяли регион в течение всего голоцена. Численность их всегда была низкой. Современная южная граница ареала росомахи проходит по Тверской и Вологодской областям (Гептнер и др., 1967). Современный ареал рыси занимает всю изучаемую территорию, а его южная граница проходит по Смоленской, Брянской и Рязанской областям (Гептнер и др., 1972).
Темный хорь согласно полученным данным заселяет данную территорию в начале атлантического периода и обитает здесь до современности (табл. 1). Небольшое количество остатков темного хоря не позволяет реконструировать северную границу его ареала в регионе. Самая северная находка этого вида известна из местонахождения Минино (59°22′ с.ш.). В настоящее время темный хорь заселяет всю территорию региона вплоть до южной части Карелии (Гептнер и др., 1967).
В материалах памятника середины бореального периода Веретье-1 и памятника начала суббореального периода Кубенино в Архангельской обл. (рис. 4) определены остатки соболя (Martes zibellina L. 1758) (Гасилин, Саблин, 2013). Кириков (1960; 1966) указывает на обитание соболя в XVI–XIX веках в двух уездах Архангельской губернии (рис. 4). На стоянке начала субатлантического периода Чудозерной в Карелии найдены две кости Martes, видовая принадлежность которых не определена (рис. 4; Верещагин, Русаков, 1979). Скорее всего, эти остатки так же принадлежат соболю. Вероятно, в течение всего голоцена (или, по крайней мере, с бореального периода) по северу центральной части Восточно-Европейской равнины проходила южная граница ареала соболя. Южнее 60° с.ш. этот вид, скорее всего, не расселялся.
Рис. 4.
Находки представителей рода Martes в центральной части Восточно-Европейской равнины в голоцене: 1 – пребореальный период, 2 – бореальный период, 3 – атлантический период, 4 – суббореальный период, 5 – субатлантический период, 6 – восточная граница современного ареала M. foina, 7 – южная граница современного ареала M. zibellina, 8 – северная граница современного ареала M. zibellina и M. martes.
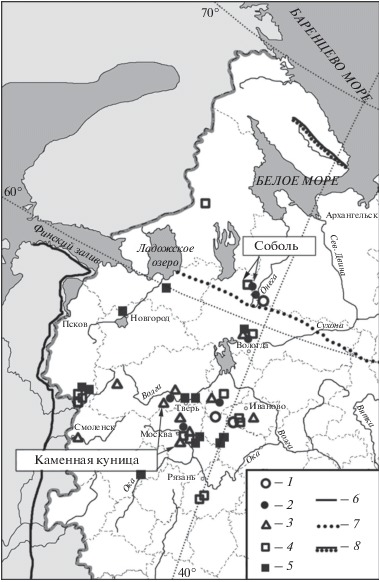
В памятнике Веретье-1 найдены остатки песца (Vulpes lagopus L. 1758) (Ошибкина, 1997). Другая находка остатков песца происходит из Пекуновского городища в Тверской обл. (Бибикова, 1950). Известны заходы песца далеко в лесную зону из тундры (Гептнер и др., 1967; Данилов и др., 2018) и эти находки обусловлены именно этим явлением.
Остается открытым вопрос об обитании в регионе каменной куницы (Martes foina Erxleben 1777). Остатки каменной куницы указаны только для местонахождений конца атлантического периода Замостье-2 (Калiноўскi, 1998) в Московской обл. и Бологое (Спицын, 1903) в Тверской обл. (рис. 4). Диагностика этого вида по костным остаткам затруднена (Гасилин, Косинцев, 2013). В памятнике Замостье-2 среди остатков, отнесенных к каменной кунице, найдены нижние челюсти и черепа, что вызывает доверие к данным определениям. Находки из памятника Бологое нуждаются в подтверждении. В настоящее время каменная куница не обитает в регионе, граница ее ареала проходит значительно западнее (Гептнер и др., 1967) (рис. 4). Для XVII–XIX веков так же нет упоминаний об обитании здесь каменной куницы (Кириков, 1960; 1966). Возможно, каменная куница обитала на западе региона очень короткий период времени в конце атлантического периода.
Северный олень обитал на данной территории на протяжении всего голоцена до начала XX века. Почти до конца XIX века южная граница ареала вида начиналась у юго-западного берега Ладожского озера, далее шла к озеру Ильмень, а от него на Валдайскую возвышенность. В ХХ столетии произошло очень резкое и быстрое отступление границы к северу на 350–650 км, причем, очевидно, исключительно под влиянием уничтожения и вытеснения этих животных человеком (Тишков, 2013).
Остатки благородного оленя (Cervus elaphus L. 1758) в регионе известны из местонахождений начиная с середины бореального до середины субатлантического периодов (Кольцов, Жилин, 1999; Саблин и др., 2011; Галимова и др., 2015) (рис. 5). Почти все кости вида найдены в памятниках, расположенных южнее 57° с.ш. Самая северо-западная находка благородного оленя известна из слоя середины субатлантического периода Старая Ладога – 59°59′ с.ш., 32°17′ в.д. (Цалкин, 1956), а самая северо-восточная – из материалов конца атлантического периода памятника Вёкса-3 – 59° с.ш., 40° в.д. (Недомолкина и др., 2015; рис. 5). Обитание благородного оленя на данной территории в конце субатлантического периода (SA3) вызывает сомнения. Авторы определения костных остатков указывают благородного оленя в материалах этого времени с вопросительным знаком (Антипина, Маслов, 1993; Кренке, 2011). В ряде памятников остатки благородного оленя представляют собой остатки рогов, которые могли быть привезены в виде сырья или готовых изделий из других регионов (Цалкин, 1962; Ермолова, 1976). В письменных источниках нет упоминаний об обитании благородного оленя в окрестностях Москвы. Вероятно, этот вид истребили здесь еще до основания города, в первом тысячелетии н.э. (Гептнер и др., 1961). В Калужской обл. остатки оленя известны не позднее первого тысячелетия н.э. (Цалкин, 1956).
Рис. 5.
Находки Cervus elaphus в центральной части Восточно-Европейской равнины в голоцене: 1 – бореальный период, 2 – атлантический период, 3 – суббореальный период, 4 – субатлантический период, 5 – граница восстановленного ареала (Гептнер и др., 1961).
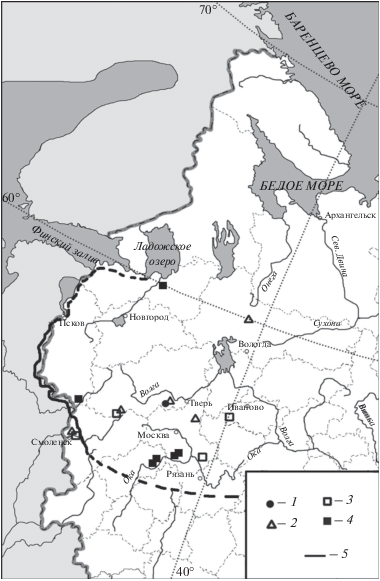
На основании всего выше сказанного, можно полагать, что благородный олень постоянно обитал в центральной части Восточно-Европейской равнины с конца раннего до середины позднего голоцена. В это время, согласно проанализированным данным, основной ареал благородного оленя располагался южнее 57°–58° с.ш. (рис. 5). В конце атлантического периода северная граница ареала вида продвинулась до 59°–60° с.ш., а позднее восстановилась в прежних границах (рис. 5). В конце субатлантического периода ареал вида сместился в прибалтийские области на восток до Пскова и южного побережья Ладожского озера, и дальше на юг по Белоруссии, западнее Смоленской обл. (Гептнер и др., 1961).
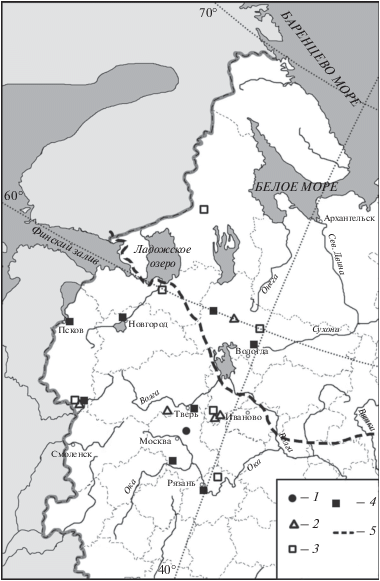
Согласно проанализированным данным кабан (Sus scrofa L. 1758) (рис. 6) и косуля (Capreolus capreolus L. 1758) (рис. 7) появляются в регионе в середине бореального периода и обитают здесь до современности (табл. 1). Причем, в бореальном периоде оба вида обитали в регионе к югу от 57°–58° с.ш. Дальше к северу до 60° с.ш. эти виды смогли расселиться только в середине атлантического периода (рис. 6, 7). Из стоянки конца атлантического – начала суббореального периода Чудозерной (62°47′ с.ш.; 38°37′ в.д.) в центре Карелии известна единственная самая северная находка рога косули (рис. 7; Верещагин, Русаков, 1979). В настоящее время известны заходы косуль к северу от основного ареала вплоть до Мурманска (Данилов и др., 2018). Возможно, эта находка обусловлена этим явлением. Возможен и другой вариант: рог могли принести люди из более южных районов, например с южного побережья Ладожского озера, где косули водились в этот период времени.
Рис. 6.
Находки Sus scrofa в центральной части Восточно-Европейской равнины в голоцене: 1 – бореальный период, 2 – атлантический период, 3 – суббореальный период, 4 – субатлантический период, 5 – граница восстановленного ареала (Гептнер и др., 1961), 6 – современная северная граница ареала (Данилов и др., 2018).
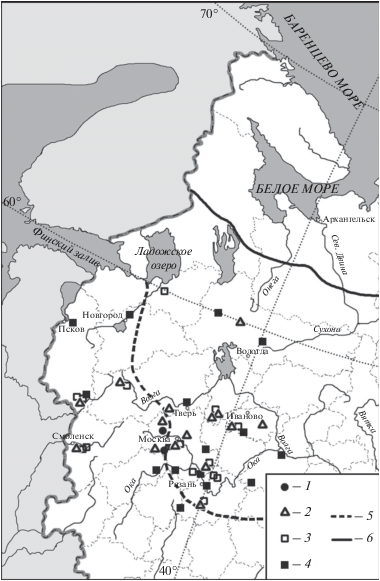
Рис. 7.
Находки Capreolus capreolus в центральной части Восточно-Европейской равнины в голоцене: 1 – бореальный период, 2 – атлантический период, 3 – суббореальный период, 4 – субатлантический период, 5 – граница восстановленного ареала (Гептнер и др., 1961).
Интенсивное истребление кабанов и косуль и сокращение их ареалов в регионе происходили во второй половине XIX века и в первые десятилетия XX века (Гептнер и др., 1961). В настоящее время ареалы видов полностью восстановились. Северная граница ареала кабана продвинулась дальше к северу по отношению к исторической границе в южную часть Карелии (Данилов и др., 2018).
Тур (Bos primigenius Bojanus 1825) был широко распространен в Прибалтике (Паавер, 1965). Восточнее Прибалтики Громовой (1931; 1949) упомянута находка роговых стержней тура из стоянки Ладога, которая датируется неолитом (SB1). Там же на берегу Ладожского озера остатки тура найдены в археологическом памятнике середины суббореального периода Подолье-1 (Галимова и др., 2015; рис. 8). Кроме того, остатки тура были найдены в двух археологических памятниках конца атлантического периода Погостище I и Векса-3 в Вологодской обл. (Брюсов, 1951; Недомолкина и др., 2015), в двух памятниках атлантического и середины суббореального периодов на территории Псковской обл. – Наумово, Усвяты 4 и Дяздица, в памятнике атлантического-суббореального периодов Сертея X в Смоленской обл., а также в памятнике конца атлантического периода Бологое в Тверской обл. (Верещагин, Русаков, 1979; Саблин и др., 2011). К сожалению, нам не известно памятников атлантического и суббореального периодов с территории современной Новгородской области, где, скорее всего, тур также обитал в этот период. Согласно имеющимся данным, основная часть ареала тура в голоцене располагалась на территории Прибалтики (Паавер, 1965). Во время климатического оптимума голоцена в конце атлантического и до середины суббореального периодов восточная граница ареала тура продвинулась до территории современной Вологодской обл. (39°–40° в.д.) (рис. 8). Южная граница ареала вида в регионе в это время проходила, вероятно, не южнее 56° с.ш. (рис. 8). Уже в конце суббореального периода ареал тура восстановился в прежних границах.
Рис. 8.
Находки Bos primigenius в атлантическом (1) и суббореальном (2) периодах, а также Equus ferus (3) в суббореальном периоде в центральной части Восточно-Европейской равнины.
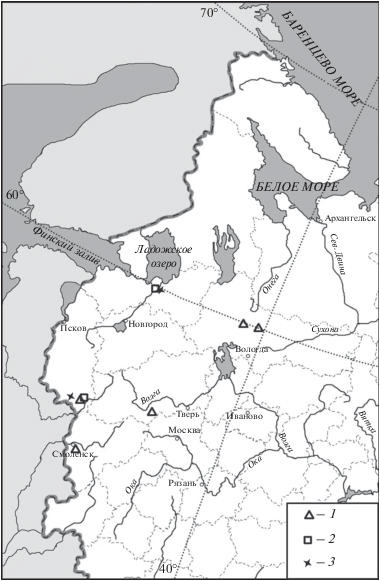
Тарпан (Equus ferus Boddaert 1785) в голоцене был распространен к западу и югу от рассматриваемого региона – в Восточной Прибалтике (Паавер, 1965) и в восточноевропейских лесостепи и степи (Кузьмина, 1997). На рассматриваемой территории остатки тарпана найдены в неолитической (SB1) стоянке на Ладоге (Громова, 1949), а также на поселении середины суббореального периода Дубокрай-5 в Псковской обл. (Саблин др., 2011) (рис. 8). Таким образом, в конце атлантического–в начале суббореального периодов восточная граница ареала дикой лошади временно продвинулась на восток до Ладоги (рис. 8). Позднее ареал тарпана восстановился в прежних границах.
ОБСУЖДЕНИЕ
На протяжении всего голоцена в центральной части Восточно-Европейской равнины обитали эврибионтные виды, виды широколиственных и бореальных лесов, лесостепные и азональные виды, большая часть из которых составляет современную фауну региона (табл. 1). Скудные сведения по фауне позднего плейстоцена региона не дают возможности определить начальный этап перестройки фауны млекопитающих от мамонтового териокомплекса к голоценовому териокомплексу. На рубеже позднего плейстоцена и раннего голоцена в центре Восточно-Европейской равнины еще обитали последние представители мамонтовой фауны (табл. 1). Самые поздние даты для мамонта (Mammuthus primigenius Blumenbach 1799) с этой территории из Вологодской обл. в районе г. Череповца – 11200 лет назад (Stuart et al., 2002), а также вблизи пос. Большая Сельменга – 11 620 лет назад (Яшина, 2002). Самые поздние остатки бизона (Bison priscus Bojanus 1827) известны из местонахождения Золоторучье-1, которое датируется рубежом DR3 и PB1 – 11 970 лет назад (Hartz et al., 2010). На р. Оять в Вологодской обл. (60° с.ш., 35° в.д.) найден череп бизона, который датируется 3250 лет назад (Markova et al., 2015). Но эта дата вызывает сомнение и нуждается в проверке.
В то же самое время в фауне пребореального периода (PB2) из местонахождения Становое-4 в Ивановской обл. (11 300 лет назад) присутствуют только лесные, лесостепные и азональные виды млекопитающих (Жилин, 2005; 2014). Следовательно, вымирание последних представителей мамонтовой фауны в центральной части Восточно-Европейской равнины произошло в течение нескольких сотен лет.
Эврибионтные виды (заяц-беляк, волк, лисица, бурый медведь, росомаха, ласка и горностай), тундрово-лесной вид северный олень обитают на данной территории с позднего плейстоцена (табл. 1). Азональные – околоводные виды (выхухоль, бобр, европейская норка), а также ряд таежных (обыкновенный еж, белка, рысь, лось, а в северной части соболь), лесных (лесная куница) и лесостепных видов (европейский барсук) заселяют данную территорию в середине пребореального периода (табл. 1). Расселение этих видов на данную территорию происходило на фоне быстрого распространения здесь лесов. Так, если в начале пребореального времени (11 700–11 200 лет назад) для центральной части Восточно-Европейской равнины реконструируются редкослойные березовые и сосновые леса с разнообразной луговой растительностью, то ближе к середине этого периода (11 200–10 800 лет назад) фиксируются значительное увеличение доли древесных пород и сокращение лугов (Спиридонова, Алешинская, 1996, 1999; Алешинская, 2001; Борисова, 2018). К периоду климатического оптимума пребореального периода (10 800–10 500 лет назад) в Волго-Окском междуречье леса становятся единственным зональным типом растительности (Нейштадт, 1957; Хотинский, 1977; Хотинский и др., 1991). К середине бореального периода окончательно исчезли безлесные участки и (в растительности) стали преобладать леса таежного типа вплоть до Архангельской обл. Во второй половине бореального периода на данной территории появляются широколиственные леса с липой и вязом (Борисова, 2018). Именно в этот период в центральную часть Восточно-Европейской равнины расселяются виды широколиственных лесов кабан и благородный олень, а также лесостепной вид – косуля. В течение атлантического периода климатические и ландшафтные зоны были смещены к северу и широколиственные породы деревьев были распространены вплоть до широты Ладожского озера (Филимонова, Лаврова, 2015; Борисова, 2018). На этом фоне на рассматриваемую территорию расселяется темный хорь – обитатель широколиственных лесов, который остается здесь до настоящего времени. В это же время продвигаются к северу ареалы благородного оленя, кабана и косули, а с прилегающих западных территорий расселяется тур, тарпан и каменная куница. В начале суббореального периода на фоне похолодания климата преобладающим типом растительности становятся темнохвойные леса, а широколиственные леса сокращаются. В это время ареалы благородного оленя, тура, тарпана и каменной куницы восстанавливаются в границах бореального периода, а ареалы барсука, выдры и норки, напротив, расширяются к северу.
В середине субатлантического периода из южной части Восточно-Европейской равнины к северу до 55° с.ш. расселяется сурок. Столь далекое проникновение сурков в лесную зону было обусловлено, вероятно, подсечной системой земледелия, в результате которой возникали многочисленные луговые (остепненные) участки, по которым и могли проникать сурки. После смены системы земледелия с подсечной на паровую возникли постоянные пашни, и сурок здесь исчез (Гроссет и др., 1965). К середине субатлантического периода с данной территории исчезает благородный олень. Причины сокращения его ареала не совсем понятны. В начале субатлантического периода на данной территории был теплый климат, и распространена темнохвойная тайга с примесью широколиственных пород деревьев. Позднее происходит похолодание климата и постепенная смена лесов на сосновые и березовые (Борисова, 2018). Возможно, климатические факторы, наряду с антропогенным прессом, могли быть причиной сокращения ареала благородного оленя. Вероятно, с середины субатлантического периода данная территория постепенно заселяется зайцем-русаком.
Подводя итог, можно сказать, что на протяжении всего голоцена на территории центра Восточно-Европейской равнины были распространены виды таежного териокомплекса, а виды широколиственных лесов и лесостепи расселялись сюда вслед за распространением неморальной растительности. Иными словами таежные элементы фауны на всей территории центра Восточно-Европейской равнины носили в течение всего голоцена зональный характер. А элементы фауны широколиственных лесов представляли собой своего рода “неморальных вселенцев”. Териофауна не была единой на всей территории региона. Выделяются северный и южный варианты териофауны, которые располагались к северу и к югу от 57° с.ш. соответственно. На севере региона в состав териофауны входил соболь, и периодически из лесотундры заходили песцы. В южной части региона на протяжении почти всего голоцена обитал благородный олень. Состав териофауны в южной части региона сформировался к началу атлантического периода, и до современности оставался относительно стабильным. Здесь кратковременно обитали в оптимуме голоцена каменная куница и тур, а в конце субатлантического периода – сурок. В северной части формирование состава фауны происходило в два этапа: ядро фауны сформировалось к началу атлантического периода, а окончательный состав сформировался лишь к началу суббореального периода, когда сюда расселились барсук, выдра, норка, кабан и косуля. В северной части в оптимуме голоцена происходили флуктуации ареалов тура и тарпана с прилегающих западных территорий, а также благородного оленя из южной части региона. Современный облик на всей территории региона фауна приобрела в конце позднего голоцена, когда под влиянием антропогенных и климатических факторов сократился ареал у одних видов и расширился у других.
Список литературы
Алексашина К.С., 1950. Остатки фауны из некоторых городищ Верхнего Поволжья // Материалы по археологии Верхнего Поволжья. № 13. М.–Л.: Изд-во АН СССР. С. 148–151.
Алешинская А.С., 2001. Палеогеографические условия обитания древнего человека в мезолите в Волго-Окском междуречье (по палинологическим данным). Автореф. дис. …канд. геогр. наук. М.: ООО “МАКС Пресс”. 25 с.
Андреева Е.Г., 1940. Фауна Кондраковского городища // Проблемы происхождения эволюции и породообразования домашних животных. Т. 1. М.–Л.: Изд-во АН СССР. С. 359–373.
Андреева Е.Г., 1972. Изучение костных остатков животных из раскопа Пронской экспедиции // Бюллетень МОИП. Отд. биол. Т. 68. С. 92–96.
Андреева Е.Г., 1974. Фауна древней стоянки Черная гора // Бюллетень МОИП. Отд. биол. Т. 79. № 4. С. 56–69.
Андреева Е.Г., 1991. Фауна поселения Крутик (по остеологическому материалу из археологических раскопок) // Белозерская весь (по материалам поселения Крутик IX–X вв.). Петрозаводск: Карельский науч. центр АН СССР. С. 182–186.
Антипина Е.Е., 2009. Ростиславльское городище дьяковского времени: археозоологические материалы из раскопок 2002-2006 годов // Аналитические исследования лаборатории естественно-научных методов. Москва: Изд-во “Таус”. С. 146–171.
Антипина Е.Е., 2013. Домашние животные в жизнеобеспечении городища Настасьино (ранний железный век) // Аналитические исследования лаборатории естественнонаучных методов. Вып. 3. М.: Институт археологии РАН. С. 142–158.
Антипина Е.Е., Грибов Н.Н., 2013. Жизнеобеспечение городища Городок и его социальный статус в нижегородской поселенческой структуре (по археозоологическим данным) // Зоологический журнал. Т. 92. № 9. С. 1001–1013.
Антипина Е.Е., Маслов С.П., 1993. К фауне позвоночных животных Переяславля-Рязанского (некоторые экологические, хозяйственные и бытовые аспекты) // Экологические проблемы в исследованиях средневекового населения Восточной Европы. М.: Институт археологии РАН. С. 224–231.
Археология. Неолит северной Евразии, 1996. М.: Наука. 380 с.
Археология СССР. Мезолит СССР, 1989. М.: Наука. 351 с.
Археология СССР. Финно-угры и балты в эпоху средневековья, 1987а. М.: Наука. 510 с.
Археология СССР. Эпоха бронзы лесной полосы СССР, 1987б. М.: Наука. 471 с.
Асылгараева Г.Ш., 2013. Исследования остеологических материалов Нижегородского кремля // Поволжская археология. № 3 (5). С. 103–115.
Бибикова В.И., 1950. Фауна Пекуновского городища // Материалы по археологии Верхнего Поволжья. № 13. М.–Л.: Изд-во Академии наук СССР. С. 133–147.
Боголюбский С.А., 1928. Палеофауна сельскохозяйственных животных Старой Рязани по раскопкам Городцова // Труды секции археологии. Т. IV. М.: РАНИОН. С. 75–89.
Борисова О.К., 2018. Развитие растительности на Верхневолжской низменности и прилегающих возвышенностях в голоцене: реконструкция по палеоботаническим данным // Стоянка Замостье 2 и развитие природной среды Волго-Окского междуречья в голоцене. СПб.: ИИМК РАН. С. 9–30.
Брюсов А.Я., 1951. Свайное поселение на р. Модлоне и другие стоянки в Чарозерском районе Вологодской области // Материалы и исследования по археологии СССР. Поселения эпохи неолита и раннего металла на севере европейской части СССР. № 20. М.–Л.: Изд-во АН СССР. С. 7–76.
Верещагин Н.К., Русаков О.С., 1979. Копытные северо-запада СССР. Л.: Наука. 308 с.
Гадзяцкая О.С., Крайнов Д.А., 2002. Энеолитическое поселение Стрелка 1 в бассейне р. Клязьма // Российская археология. № 2. С. 111–131.
Галимова Д.Н., Аскеев Б.В., Аскеев О.В., Гусенцова Т.М., 2015. Археозоологический анализ неолитической стоянки Подолье-1 в южном Приладожье // Неолитические культуры Восточной Европы: хронология, палеоэкология, традиции. СПб.: ИИИМК РАН. С. 163–165.
Гасилин В.В., 2005. Фауна крупных млекопитающих лесостепной зоны Волго-Уральского региона в голоцене // Экология: от генов до экосистем: материалы конф. молодых ученых, 25–29 апр. 2005 г. Екатеринбург: Академкнига. С. 53–54.
Гасилин В.В., 2013. Крупные млекопитающие Приморья в голоцене // Зоологический журнал. Т. 92. № 9. С. 1055–1063.
Гасилин В.В., Косинцев П.А., 2013. Диагностика видов подрода Martes str. (Carnivora, Mustelidae) по метрическим признакам нижней челюсти // Зоологический журнал. Т. 92. № 2. С. 221–230.
Гасилин В.В., Саблин М.В., 2013. Виды рода Martes на северо-западе Восточно-Европейской равнины в голоцене // Труды Зоологического института РАН. Т. 317. № 2. С. 125–135.
Гептнер В.Г., Насимович А.А., Банников А.Г., 1961. Млекопитающие Советского Союза. Т. 1. Парнокопытные и непарнокопытные. М.: Высшая школа. 776 с.
Гептнер В.Г., Наумов Н.П., Юргенсон П.Б., Слудский А.А., Чиркова А.Ф., Банников А.Г., 1967. Млекопитающие Советского Союза. Т. 2 (Ч. I). Морские коровы и хищные. М.: Высшая школа. 1004 с.
Гептнер В.Г., Слудский А.А., 1972. Млекопитающие Советского Союза. Т. 2 (Ч. II). Хищные (гиены и кошки). М.: Высшая школа. 552 с.
Городцов В.А., 1933. Старшее Каширское (результаты археологических исследований в 1925–1926 гг.) // Известия Государственной Академии истории материальной культуры. Вып. 85. М.–Л.: Соцэкгиз. С. 38–39.
Громова В.И., 1931. Первобытный бык или тур в СССР // Ежегодник Зоол. музея АН СССР. Т. 32. Вып. 3. С. 293–366.
Громова В., 1949. История лошадей (род Equus) в Старом Свете // Труды Палеонтологического института. Т. 17. Часть 1. 374 с.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. СПб.: ЗИН РАН. 522 с.
Гроссет Г.Э., Динесман Л.Г., Цалкин В.И., 1965. О древнем распространении степного сурка // Бюллетень Московского общетсва испытателей природы. Отдел биологический. Вып. 2. С. 34–46.
Давид А.И., 1982. Формирование териофауны Молдавии в антропогене. Кишинев: Штиица. 151 с.
Данилов П.И., Панченок Д.В., Тирронен К.Ф., Федоров Ф.В., Белкин В.В., 2018. Изменение фауны млекопитающих Северной Палеарктики и динамика ареалов составляющих ее видов // Известия РАН. Серия биологическая. № 3. С. 301–314.
Данильченко В.П., 1984. Остеологический материал Селецкого городища // Краткие сообщения Института археологии. Вып. 178. М.: Изд-во АН СССР. С. 29–34.
Данильченко В.П., Карху А.А., 1997. Фаунистические определения материалов поселения Воймежное I // Древние охотники и рыболовы Подмосковья. М.: Институт археологии РАН. С. 130–132.
Девяшин М.М., Косинцев П.А., 2013. Копытные млекопитающие среднего и позднего голоцена юго-востока Западной Сибири // Вестник Томского государственного унверситета. Биология. № 2 (22). С. 127–140.
Дмитриева Е.Л., Сердюк Н.В., Лавров А.В., Сычевская Е.К., Зеленков Н.В., 2008. Палеозоологическая характеристика археологического памятника “Боровский курган” // Археология Подмосковья: Материалы научного семинара. Вып. 4. М.: Институт археологии РАН. С. 389–391.
Добровольская Е.В., 2012. Животные Минино-2 // Первобытные древности Евразии: к 60-летию Алексея Николаевича Сорокина. М.: Институт археологии РАН. С. 299–306.
Ермолова Н.В., 1976. Остатки млекопитающих с древнего городища Осыно // Советская археология. №3. С. 217–228.
Жилин М.Г., 1997. Памятники мезолита и раннего неолита западной части Дубненского торфяника // Древности Залесского края. Сергиев Посад: Сергиево-Посадский музей-заповедник. С. 164–196.
Жилин М.Г., 2004. Природная среда и хозяйство мезолитического населения центра и северо-запада лесной зоны Восточной Европы. М.: Академия. 142 с.
Жилин М.Г., 2005 Некоторые итоги раскопок многослойного торфяникового поселения Становое 4 // Краткие сообщения Института археологии. Вып. 219. М.: Наука. С. 21–35.
Жилин М.Г., 2014. Радиоуглеродная хронология стоянок мезолита-неолита Озерки 5 и 17 на Верхней Волге // Археология озерных поселений IV–II тыс. до н. э.: хронология культур и природно-климатические ритмы. СПб.: ООО “Периферия”. С. 55–60.
Зайцева Г.И., Лозовская О.В., Выборнов А.А., Мазуркевич А.Н., 2016. Радиоуглеродная хронология эпохи неолита Восточной Европы VII–III тысячелетия до н. э. Смоленск: Свиток. 456 с.
Зиновьев А.В., 2014. Охотничьи виды млекопитающих средневекового Новгорода Великого и его окрестностей (по археозоологическим данным) // Вестник ТвГУ. Серия биология и экология. № 4. С. 86–94.
Иванечева М.А., 2007. Погребения каменного века в Нижнем Посухонье // Русская культура нового столетия: Проблемы изучения, сохранения и использования историко-культурного наследия. Вологда: Книжное наследие. С. 6–20.
Иностранцев А.А., 1882. Доисторический человек каменного века побережья Ладожского озера. Т. XVIII. СПб.: Типография М.М. Стасюлевича. 243 с.
Калiноўскi П.Ф., 1998. Тэрыяфауна архелагiчнага помнiка Замосце (Маскоўская вобласць Расіі) // Літасфера. № 8. С. 34–44.
Карху А.А., Кириллова И.В., Жилин М.Г., 2003. Охотничий промысел древнего населения стоянки Ивановская VII // Новейшие археозоологические исследования в России. К столетию со дня рождения В.И. Цалкина. М.: Языки славянской культуры. С. 139–156.
Кириков С.В., 1960. Изменения животного мира в природных зонах СССР (XIII – XIX вв.). Лесная зона и лесотундра. М.: Изд-во АН СССР. 158 с.
Кириков С.В., 1966. Промысловые животные, природная среда и человек. М.: Наука. 346 с.
Кириков С.В., 1980. Исторические изменения в размещении байбака (XVII-XIX вв. и первая треть XX в.) // Сурки. Биоценотическое и практическое значение. М.: Наука С. 24–31.
Кириллова И.В., 1992. Костные остатки млекопитающих стоянки Языково I // Многослойные стоянки Верхнего Поволжья. М.: Институт археологии РАН. С. 113–129.
Кириллова И.В., 1998. Охотничья добыча обитателей стоянки Ивановское VII (по костным остаткам млекопитающих) // Некоторые итоги изучения археологических памятников Ивановского болота. Иваново: Ивановский государственный университет. С. 63–83.
Кириллова И.В., Емельянов А.В., 1999. Роль млекопитающих в хозяйстве энеолитического человека центральной Мещеры (на примере поселения Шагара-2) // Экология древних и современных обществ. Тезисы докладов конференции, посвященной 275-летию РАН. Тюмень: Изд-во ИПОС СО РАН. С. 109–112.
Кольцов Л.В., Жилин М.Г., 1999. Мезолит Волго-Окского междуречья. (Памятники бутовской культуры). М.: Наука. 157 с.
Короткевич Б.С., Саблин М.В., Сыромятников Е.В., 2010. Фауна городища Анашкино в эпоху поздней бронзы и раннего железного века // Археологический сборник Государственного Эрмитажа: материалы и исследования по археологии Евразии. Вып. 38. СПб.: Изд-во Гос. Эрмитажа. С. 24–29.
Косинцев П.А., Гасилин В.В., 2005. Фауна крупных млекопитающих степной зоны Волго-Уралья в голоцене // Проблемы палеонтологии и археологии юга России и сопредельных территорий: материалы международной конференции. Ростов н/Д: Изд-во ЦВВР. С. 43–45.
Косорукова Н.В., 2012. Торфяниковая мезолитическая стоянка Погостище XIV в бассейне озера Воже (по материалам исследований 2005, 2008, 2009 гг.) // История и археология Русского Севера. Вологодское археологическое общество Вологодский государственный музей-заповедник. Сборник материалов научной конференции, посвященной 60-летию со дня рождения В. Гуслистова. Вологда: Древности Севера. С. 58–63.
Костылёва Е.Л., Уткин А.В., 2010. Неолитическая стоянка Ивановское II // Вестник Ивановского государственного университета. Серия гуманитарные науки. № 4 (10). С. 32–61.
Кренке Н.А., 2011. Дьяково городище: культура населения бассейна Москвы-реки в I тыс. до н.э. – I тыс. н.э. М.: Институт археологии РАН. 548 с
Кренке Н.А., Александровский А.Л., Елькина И.И., Ершов И.Н., Лазукин А.В. и др., 2012. Новые исследования 1-й Звенигородской неолитической стоянки на Москве-реке // Археология Подмосковья. Материалы научного семинара. Вып. 8. М.: Институт археологии РАН. С. 16–34.
Крылович О.А., 2006. Археозоологический материал из средневековых поселений Вишенки III и Весь V (Владимирская обл., Суздальский р-н) // История экосистем в голоцене. М.: Товарищество научных изданий КМК. С. 116–119.
Крылович О.А., Савинецкий А.Б., 2005. Динамика промысловой фауны Северо-Запада России // Фундаментальные основы управления биологическими ресурсами. М.: Товарищество научных изданий КМК. С. 291–300.
Кузьмина И.Е., 1997. Лошади Северной Евразии от плиоцена до современности // Труды зоологического института. Т. 273. СПб.: ЗИН РАН. 224 с.
Кузьмина С.А., 2003. Новые фаунистические данные по результатам раскопок неолитических памятников Смоленской и Псковской областей // Древности Подвинья: исторический аспект. СПб.: Изд-во Гос. Эрмитажа. С. 300–316.
Лапшин В.А., 2009. Тверь в XIII – XV вв. (по материалам раскопок 1993 – 1997). СПб.: Факультет филологии и искусств СПбГУ. 540 с.
Лисицын С.Н., 2011. Стоянка Вашана и вопросы хроностратиграфии раннего мезолита Волго-Окского междуречья // Записки Института истории материальной истории. № 6. СПБ.: ИИМК РАН. С. 104–118.
Лозовский В.М., Лозовская О.В., Зайцева Г.И., Кулькова М.Э., 2014. Радиоуглеродная хронология культурных отложений эпохи мезолита и неолита стоянки Замостье 2 // Археология озерных поселений IV–II тыс. до н. э.: хронология культур и природно-климатические ритмы. СПб.: ООО “Периферия”. С. 61–64.
Мильков Ф.Н., 1977. Природные зоны СССР. М.: Мысль. 293 с.
Недомолкина Н.Г., Пиецонка Х., Лоренц С., Шмелке У., 2015. Новые археологические, остеологические и геоморфологические исследования на комплексе многослойных поселений Векса в бассейне верхней Сухоны // Тверской археологический сборник. Т. 10. Вып. 1. Тверь: ООО Издательство Триада. С. 74–84.
Нейштадт М.И., 1957. История лесов и палеогеография СССР в голоцене. М.: Изд-во АН СССР. 404 с.
Ошибкина С.В., 1997. Веретье-1. Поселение эпохи мезолита на Севере Восточной Европы. М.: Наука. 204 с.
Паавер К.А., 1965. Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту: АН Эстонской ССР. 494 с.
Пластеева Н.А., Клементьев А.М., 2018. Крупные млекопитающие Прибайкалья в голоцене // Зоологический журнал. Т. 97. № 5. С. 599–612.
Саблин М.В., Пантелеев А.В., Сыромятникова Е.В., 2011. Археозоологический анализ остеологического материала из неолитических свайных поселений Подвинья: хозяйство и экология // Труды Зоологического института РАН. Т. 315. № 2. С. 143–153.
Савинецкий А.Б., 1993. К истории распространения сурка в европейской части России // Международное (V) Совещание по суркам стран СНГ. 21–23 сентября 1993 года, с. Гайдары (Украина). Тезисы докладов. М. С. 34.
Савинецкий А.Б., Крылович О.А., 2009. Палеозоологические материалы и динамика промысловой фауны // Археология северно-русской деревни X–XIII веков: средневековые поселения и могильники на Кубенском озере. Т. 3. М.: Наука. С. 16–24.
Седов В.В., 1982. Восточные славяне в VI–XIII вв. М.: Наука. 328 с.
Сердюк Н.В., Кренке Н.А., Зеленков Н.В., Сычевская Е.К., 2018. Остеологическая коллекция селища Хотяжи 1 // Российская археология. № 3. С. 121–132.
Спиридонова Е.А., Алешинская А.С., 1996. Особенности формирования и структуры растительного покрова Волго-Окского междуречья в эпоху мезолита // Тверской археологический сборник. Вып. 2. Тверь: Тверской гос. объединенный музей. С. 65–71.
Спиридонова Е.А., Алешинская А.С., 1999. Результаты палинологического изучения мезолита Волго-Окского междуречья // Мезолит Волго-Окского междуречья. Памятники бутовской культуры. М.: Наука. С. 137–145.
Спицын А.А., 1903. Бологовская стоянка каменного века // ЗОРСА. Т. 5. Вып. 1. СПб. С. 239–277.
Тимофеев В.И., Зайцева Г.И., Долуханов П.М., Шукуров А.М., 2004. Радиоуглеродная хронология неолита Северной Евразии. СПб.: Теза. 157 с.
Тишков А.А., 2013. Тысячелетняя история изменений фауны млекопитающих Валдайского поозерья // Вопросы географии. Вып. 136. С. 385–411.
Третьяков П.Н., 1941. К истории племен Верхнего Поволжья в первом тысячелетии н.э. № 5. М.–Л.: Изд-во АН СССР. 150 с.
Филиминова Л.В., Лаврова Н.Б., 2015. Палеогеография заонежского полуострова в позднем плейстоцене и голоцене // Труды Карельского научного центра РАН. № 4. С. 30–47.
Формозов А.А., 1961. Очерки по истории Восточно-Европейской археологии. М.: Изд-во АН СССР. 126 с.
Формозов А.Н., 1948. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930–1940 гг. // Фауна и экология грызунов. Вып. 3. М.: МОИП. С. 3–110.
Хотинский Н.А., 1977. Голоцен северной Евразии. М.: Наука. 200 с.
Хотинский Н.А., Алешинская З.В., Гуман М.А., Климанов В.А., Черкинский А.Е., 1991. Новая схема периодизации ландшафтно-климатических изменений в голоцене // Известия АН СССР. Серия географическая. № 3. С. 30–42.
Цалкин В.И., 1956. Материалы для истории скотоводства и охоты в Древней Руси. По данным изучения костных остатков из раскопок археологических памятников лесной полосы Европейской части СССР. М.: Изд-во АН СССР. 185 с.
Цалкин В.И., 1961. Млекопитающие бассейна Оки и Верхней Волги в начале нашей эры // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 46. № 1. С. 23–39.
Цалкин В.И., 1962. Животноводство и охота в лесной полосе Восточной Европы в раннем железном веке // К истории животноводства и охоты в Восточной Европе. Материалы и исследования по археологии СССР. № 107. М.: Изд-во АН СССР. С. 5–96.
Цалкин В.И., 1971. Некоторые итоги изучения костных остатков животных из раскопок Москвы // Древности Московского кремля. Материалы и исследования по археологии СССР. Материалы и исследования по археологии Москвы. Т. IV. № 167. М.: Наука. С. 164–185.
Шэ Л., Шеналь-Велярд И., Велярд Л., 2001. Анализ фаунистических остатков из мезолитических слоев Замостье-2: первые результаты // Каменный век европейских равнин. Сергиев Посад: Сергиево-Посадский гос. историко-художественный музей-заповедник. С. 255–261.
Энговатова А.В., Антипина Е.Е., Бужилова А.П., Гончарова Н.Н., Добровольская М.В. и др., 2012. Археология древнего Ярославля: загадки и открытия (по материалам Ярославской экспедиции ИА РАН). М.: Институт археологии РАН. 296 с.
Яшина О.В., 2002. Краткая характеристика костей мамонта в коллекции Вологодского, Череповецкого и Тотемского музеев // Материалы третьего Всероссийского совещания по изучению четвертичного периода. Т 2. Смоленск: Ойкумена. С. 155–158.
Bachura O., Kosintsev P., 2007. Late Pleistocene and Holocene small- and large-mammal faunas from the Northern Urals // Quaternary International. V. 160. P. 121–128.
Hartz S., Terberger T., Zhilin M., 2010. New AMS-dates for the Upper Volga Mesolithic ahd the origin of microblade technology in Europe // Quartär. V. 57. P. 155–169.
Kosintsev P A., Bachura O.P., 2013. Late Pleistocene and Holocene mammal fauna of the Southern Urals // Quaternary International. V. 284. P. 161–170.
Markova A.K., Puzachenko A.Yu., Kolfschoten T. van, Kosintsev P.A., Kuznetsova T.V. et al., 2015. Changes in the Eurasian distribution of the musk ox (Ovibos moschatus) and the extinct bison (Bison priscus) during the last 50 ka BP // Quaternary International. V. 378. P. 99–110.
Piezonka H., Meadows J., Hartz S., Kostyleva E., Nedomolkina N. et al., 2016. Stone age pottery chronology in the Northeast European forest zone: New AMS and EA-IRMS results on foodcrusts // Radiocarbon. V. 58. № 2. P. 267–289.
Stuart A.J., Sulerzhitsky L.D., Orlova L.A., Kuzmin Y.V., Lister A.M., 2002. The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: a review of the current evidence // Quaternary Science Reviews. V. 21. № 14–15. P. 1559–1569.
Zaretskaya N.E., Zhilin M.G., Rarmanov V.N., Uspenskaya O.N., 2005. Radiocarbon dating of wetland meso-neolithic archaeological sites within the Upper Volga and Middle Vychegda // Geochronometria. V. 24. P. 117–131.
Zhilin M.G., 2014. Early Mesolithic Hunting and Fishing Activities in Central Russia: A Review of the Faunal and Artefactual Evidence from Wetland Sites // Journal of wetland archaeology. V. 14. P. 91–105.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал