

Зоологический журнал, 2020, T. 99, № 5, стр. 554-568
Голоценовое распространение и вымирание копытных в Северной Евразии
Н. А. Пластеева a, *, В. В. Гасилин a, **, М. М. Девяшин a, ***, П. А. Косинцев a, ****
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: natalya-plasteeva@yandex.ru
** E-mail: gasilinv@yandex.ru
*** E-mail: devjashinm@yandex.ru
**** E-mail: kpa@ipae.uran.ru
Поступила в редакцию 20.08.2019
После доработки 30.09.2019
Принята к публикации 21.10.2019
Аннотация
Обобщены и проанализированы литературные и оригинальные данные о вымерших в голоцене копытных животных Северной Евразии. Приведены данные об абсолютном и относительном возрасте и местах сбора костных находок дикой лошади (Equus ferus Boddaert 1785), европейского осла (Equus hydruntinus Regalia 1907), гигантского оленя (Megaloceros giganteus Blumenbach 1799), первобытного бизона (Bison priscus Bojanus 1825), овцебыка (Ovibos moschatus Zimmermann 1780) и тура (Bos primigenius Bojanus 1825); в сравнительном аспекте обсуждаются характер сокращения ареалов и время исчезновения видов. Сделано заключение о том, что в последнюю межледниковую эпоху изменения границ ареалов происходили с разной скоростью не только у разных видов, но и у одного вида в разных частях ареала.
В конце плейстоцена–начале голоцена произошли существенные изменения видового состава и границ ареалов ряда видов млекопитающих. Этот процесс затронул разные таксономические группы, в том числе копытных. Причины сокращения численности и последующего исчезновения крупных травоядных млекопитающих связывают с глобальным изменением климата и его последствиями (Guthrie, 2006; Lister, Stuart, 2008; Lorenzen et al., 2011; Cooper et al., 2015), которые сопровождались ростом интенсивности хозяйственного освоения природных ресурсов человеком.
Повышение средней температуры окружающей среды и значительный рост увлажненности, наблюдаемые в финале плейстоцена (Rabanus-Wallace et al., 2017), приводят к постепенной деградации перигляциальных криоаридных ландшафтов и формированию современной природной зональности. С началом голоцена на всей территории Северной Евразии начинаются быстрое развитие лесного покрова и уменьшение площади открытых ландшафтов (MacDonald et al., 2000; Хотинский, Климанов, 2002; Binney et al., 2009; Tarasov et al., 2009). Ландшафтно-климатические изменения повлекли за собой сокращение численности и исчезновение одних видов млекопитающих, расширению ареалов и появлению в Северной Евразии других видов.
В силу своих размеров, стадного образа жизни и широкого распространения крупные травоядные млекопитающие были значимы в жизнеобеспечении древних человеческих коллективов. Охота на копытных животных занимала значительное место в хозяйстве с плейстоцена, но с началом голоцена группа видов травоядных животных сократила свои ареалы и впоследствии вымерла. Эти процессы отмечаются на всей территории, однако их региональное и локальное проявление было различным.
Целью настоящей работы является обобщение и анализ сведений о находках костей исчезнувших в голоцене копытных животных Северной Евразии – дикой лошади (Equus ferus Boddaert 1785), европейского осла (Equus hydruntinus Regalia 1907), гигантского оленя (Megaloceros giganteus Blumenbach 1799), тура (Bos primigenius Bojanus 1825), первобытного бизона (Bison priscus Bojanus 1825) и овцебыка (Ovibos moschatus Zimmermann 1780). В настоящий момент подробные сведения о голоценовом распространении вымерших копытных имеются в основном для Западной и Центральной Европы (Benecke, 1999; Stuart et al., 2004; van Vuure, 2005; Sommer et al., 2011; Crees, Turvey, 2014; Crees et al., 2016; Lister, Stuart, 2019). Данные для остальной части субконтинента являются зачастую разрозненными, а для некоторых видов – неполными или прежде не рассматривались.
Нами обобщена известная информация о находках костных остатков вымерших копытных животных и их возрасте, изучен характер их географического распределения в разные периоды голоцена, и на основе полученных данных уточнены области обитания видов. Полученные результаты позволяют проследить характер деградации ареалов рассматриваемых видов животных и получить более детальную картину по хронологии их вымирания.
МАТЕРИАЛ И МЕТОДЫ
Изучены опубликованные, оригинальные и архивные сведения об остеологических находках шести видов копытных более 200 голоценовых местонахождений Северной Евразии. Область исследования охватывает Восточную Европу, Предкавказье, Урал, Казахстан, Западную, Среднюю, Южную, Северо-Восточную Сибирь и Приморье, по которым в настоящее время проходят природные зоны от арктических пустынь до полупустынь.
Возраст находок костей изучаемых видов животных оценен на основании некалиброванных радиоуглеродных дат, полученных как непосредственно по кости (табл. 1), так и косвенно – по другим органическим остаткам из вмещающего ее слоя, а также при помощи археологических и стратиграфических методов.
Таблица 1.
Некалиброванные радиоуглеродные даты по костям E. ferus, M. giganteus, B. primigenius, B. priscus и O. moschatus
| Местонахождение | Координаты с.ш., в.д. |
Лабораторный № | C14 дата | Источник |
|---|---|---|---|---|
| Лошадь дикая (Equus ferus) | ||||
| р. Часовня | 61.2, 128.2 | ГИН–14 697 | 9790 ± 60 | Боескоров и др., 2016 |
| Шолма 1 | 55.5, 47.3 | Ki–15 156 | 9200 ± 200 | Гугалинская и др., 2010 |
| р. Ньюнкаракутари | 75.2, 105.5 | ГИН–10 509 | 9010 ± 140 | MacPhee et al., 2002 |
| Шолма 1 | 55.5, 47.3 | Ki–15 155 | 8160 ± 120 | Гугалинская и др., 2010 |
| Черемухово 1-2 | 60.2, 60.0 | СОАН–5138 | 8030 ± 120 | Bachura, Kosintsev, 2007 |
| о-в Вилькицкий | 75.4, 152.3 | Beta–191 338 | 7900 ± 40 | Анисимов и др., 2009 |
| Кунда | 59.5, 26.6 | KIA–35 735 | 7385 ± 35 | Sommer et al., 2011 |
| Преображенка 6 | 55.3, 77.0 | UBA–27 425 | 7214 ± 63 | Marchenko et al., 2015 |
| Турганикское | 52.5, 53.5 | SPb–1501 | 5816 ± 80 | Моргунова и др., 2017 |
| Кяэпа | 57.5, 27.1 | KIA–35 737 | 5698 ± 73 | Sommer et al., 2011 |
| Турганикское | 52.5, 53.5 | SPb–1861 | 5615 ± 80 | Моргунова и др., 2017 |
| Туба 1 | 48.5, 38.3 | Ki–8256 | 5480 ± 80 | Манько и др., 2001 |
| Большая Элиста 1 | 46.6, 43.7 | GrA–51 016 | 5260 ± 35 | Шишлина и др., 2015 |
| Маяки | 46.3, 30.2 | Poz–24 927 | 4770 ± 40 | Pruvost et al., 2011 |
| Ойягосский Яр | 72.5, 142.5 | GrA–540 209 | 4630 ± 35* | Боескоров и др., 2013 |
| п-ов Быковский | 71.3, 129.4 | ГИН–10 256 | 4610 ± 40 | Kuznetsova et al., 2001 |
| Ново-Байрамгулово | 54.1, 59.0 | СОАН–7274 | 4415 ± 125 | Косинцев, Пластеева, 2013 |
| Новоильинка III | 53,3, 79.2 | СОАН–8318 | 4585 ± 170 | Ситников и др., 2007 |
| Новоильинка III | 53.3, 79.2 | ЛЕ–7534 | 4270 ± 170 | Ситников и др., 2007 |
| Батагайское | 67.5, 134.5 | GrA–48 709 | 4400 ± 35 | Лазарев и др., 2011 |
| р. Орда | 53.5, 81.5 | СОАН–4569 | 4315 ± 45 | Васильев и др., 2007 |
| р. Агапа | 71.6, 87.0 | ГИН–3243 | 3250 ± 60 | Сулержицкий, Романенко, 1997 |
| о-в. Котельный | 75.2, 141.0 | ГИН–13237 | 3000 ± 45 | Кузнецова, ван дер Плихт, 2009 |
| оз. Мойчоон, Хромская губа | 70.0, 143.0 | ЛУ–1084 | 2310 ± 80 | Лазарев, 1980 |
| о-в. Большой Ляховский | 73.3; 141.3 | ГИН–10 687 | 2200 ± 50 | Kuznetsova et al., 2001 |
| р. Большая Балахня | 73.6; 100.5 | ГИН–2744 | 2150 ± 200 | Сулержицкий, Романенко, 1997 |
| Олень гигантский (Megaloceros giganteus) | ||||
| Кульметовский грот | 58.5, 55.2 | OxA–10 676 | 10 260 ± 55 | Stuart et al., 2004 |
| Черниговский рудник | 55.4, 86.0 | OxA–13 026 | 10 055 ± 45 | Stuart et al., 2004 |
| Улуир | 55.1, 58.6 | OxA–19 685 | 10 020 ± 40 | Lister, Stuart, 2019 |
| Бобылек | 56.3, 57.4 | OxA–11 063 | 9960 ± 55 | Stuart et al., 2004 |
| Сосновый Тушамский | 58.3, 102.9 | GrA–56 931 | 9450 ± 50 | van der Plicht et al., 2015 |
| Сосновый Тушамский | 58.3, 102.9 | GrA–56 932 | 9405 ± 45 | van der Plicht et al., 2015 |
| Усть-Талая 1 | 55.4, 103.0 | GrA–56 936 | 9235 ± 45 | van der Plicht et al., 2015 |
| Челябинск | 55.0, 61.0 | OxA–19 681 | 8276 ± 36 | Lister, Stuart, 2019 |
| Шигирский торфяник | 60.2, 57.4 | OxA–11 064 | 7990 ± 45 | Stuart et al., 2004 |
| Сопка 2 | 55.4, 76.4 | GrA–56 934 | 7925 ± 40 | van der Plicht et al., 2015 |
| Преображенка 6 | 55.3, 76.6 | OxA–23 412 | 7889 ± 38 | Lister, Stuart, 2019 |
| Преображенка 6 | 55,3, 76.6 | GrA-56 935 | 7865 ± 40 | van der Plicht et al., 2015 |
| Мергень 6 | 56.0, 69.5 | OxA–27 615 | 7321 ± 33 | Lister, Stuart, 2019 |
| Мергень 6 | 56.0, 69.5 | OxA–27 706 | 7147 ± 38 | Lister, Stuart, 2019 |
| Редут | 64.2, 55.5 | OxA–10 705 | 7100 ± 50 | Stuart et al., 2004 |
| Редут | 64.2, 55.5 | OxA–13 014*** | 7034 ± 34 | Stuart et al., 2004 |
| Редут | 64.2, 55.5 | KIA–5668*** | 6968 ± 33 | Stuart et al., 2004 |
| Камышлов | 62.7, 56.9 | OxA–13 015*** | 6881 ± 38 | Stuart et al., 2004 |
| Камышлов | 62.7, 56.9 | KIA–5669*** | 6816 ± 35 | Stuart et al., 2004 |
| Малоархангельск | 52.4, 36.5 | OxA–19 488 | 6832 ± 33 | Lister, Stuart, 2019 |
| Малоархангельск | 52.4, 36.5 | OxA–19 487 | 6829 ± 33 | Lister, Stuart, 2019 |
| Тур (Bos primigenius) | ||||
| р. Барык | 51.1, 93.0 | СОАН–6336 | 9860 ± 160 | Лавров, Забелин, 2007 |
| Мирное | 46.3, 30.2 | GrA–37336 | 8280 ± 45 | Biagi et al., 2008 |
| Краснокаменка | 54.4, 60.1 | СОАН–5754 | 8040 ± 210 | Мосин, 2000 |
| Бугульдейка II | 52.3, 105.5 | OxA–23949 | 4966 ± 30 | Лозей и др., 2014 |
| Бугульдейка II | 52.3, 105.5 | OxA–23948 | 4940 ± 30 | Лозей и др., 2014 |
| Бизон первобытный (Bison priscus) | ||||
| Золоторучье 1 | 57.3, 38.1 | KIA–39 314 | 10 240 ± 37 | Hartz et al., 2010 |
| р. Раучуа | 69.3, 166.3 | AA–101 271 | 9497 ± 92* | Kirillova et al., 2015 |
| р. Раучуа | 69.3, 166.3 | SPb–743 | 8030 ± 70** | Kirillova et al., 2015 |
| о. Врангеля | 71.0, 178.0 | ЛУ–2801 | 9450 ± 100 | Vartanyan et al., 2008 |
| оз. Чукчалах | 71.5, 140.3 | GrA–53 290 | 9310 ± 45* | Боескоров и др., 2013 |
| оз. Чукчалах | 71.5, 140.3 | GrA–53 292 | 9295 ± 45** | Боескоров и др., 2013 |
| Куналей | 50.6,107.8 | ГИН–14 668 | 9000 ± 270 | Зарецкая и др., 2015 |
| р. Попигай | 72.8, 107.4 | Beta-148 623 | 8860 ± 40 | MacPhee et al., 2002 |
| Батагай | 67.0, 134.0 | GrA–50 838 | 8215 ± 45 | Григорьев и др., 2017 |
| р. Оять | 60.4, 35.3 | GrA–41 240 | 3045 ± 35 | Markova et al., 2015 |
| Овцебык (Ovibos moschatus) | ||||
| п-ов Таймыр | 74.0, 101.0 | AAR–11 749 | 5364 ± 49 | Lorenzen et al., 2011 |
| п-ов Таймыр | 74.0, 101.0 | OxA–17 064 | 4082 ± 30 | Campos et al., 2010 |
| мыс Челюскин (Таймыр) | 77.4, 104.2 | б/н | 3800 ± 200 | MacPhee et al., 2002 |
| п-ов Таймыр | 74.0, 101.0 | AAR–11 744 | 3372 ± 43 | Lorenzen et al., 2011 |
| п-ов Быковский | 71.3, 129.4 | ГИН–10 259 | 3180 ± 100 | Kuznetsova et al., 2001 |
| мыс Челюскин (Таймыр) | 77.4, 104.2 | ГИН–2945 | 2920 ± 50 | Сулержицкий, Романенко, 1997 |
| п-ов Таймыр | 74.0, 101.0 | OxA–17 063 | 2918 ± 28 | Campos et al., 2010 |
| Прончищева п-ов (Таймыр) | 75.1, 112.3 | ГИН–25 529 | 2900 ± 60 | MacPhee et al., 2002 |
| п-ов Таймыр | 74.0, 101.0 | OxA–17 062 | 2756 ± 27 | Campos et al., 2010 |
| п-ов Таймыр, р. Логата | 73.2, 97.0 | ГИН–3803 | 2700 ± 70 | Сулержицкий, Романенко, 1997 |
| Тумат | 59.4, 130.4 | GrA–62 450 | 2535 ± 30 | Boeskorov et al., 2017 |
Примечания. В случае наличия двух и более дат для находок из одного местонахождения приводятся их максимальное и минимальное значения. * – радиоуглеродная дата получена по кости или рогу. ** – радиоуглеродная дата получена по шерсти или мягким тканям того же образца. *** – радиоуглеродные даты получены по одному образцу.
Хронологический интервал исследования – от раннего голоцена (10 200 л. н.) до исторического времени (условно 200 л. н.). Периодизация голоцена принята со следующими границами (Хотинский и др., 1991): ранний голоцен (пребореальный и бореальный периоды, PB–BO, 10 200–8000 л. н.), средний голоцен (атлантический период AT, эпохи неолита и энеолита, 8000–4600 л. н. и суббореальный период, SB, финал энеолита–бронзовый век, 4600–2600 л. н.), поздний голоцен (субатлантический период, ранний железный век и средневековье, SA, 2600–200 л. н.).
Картографирование восстановленных ареалов выполнено точечным методом: местонахождения с находками рассматриваемых видов нанесены на карты для каждого хронологического интервала. Местонахождения с костными остатками, датированными непосредственно, и местонахождения с находками, возраст которых определен на основании радиоуглеродных дат слоя и археологическими методами, на карте отмечены разными значками. Схематическая карта позволяет получить представление об общих очертаниях ареалов видов в прошлом.
Для ряда видов имеются методические проблемы, связанные с идентификацией костных остатков. Морфологические различия костей крупных быков (роды Bos и Bison) незначительны. Наибольший диагностический вес во всех случаях имеют целые черепа и роговые стержни, которые не всегда представлены в материале. Кости посткраниального скелета представителей Bos и Bison различают морфологически с определенными методическими трудностями (Бибикова, 1958; Olsen, 1960; Stampfli, 1963). Представителя Bos относят к дикой форме B. primigenius на основании крупных, выходящих за пределы изменчивости у домашней формы B. taurus, размеров зубов и костей посткраниального скелета (Цалкин, 1970). Представителя Bison относят к бизону B. priscus также исходя из размеров костей, превышающих таковые у зубра B. bonasus, однако оценка значений этих границ зачастую проводится субъективно. Поэтому видовая принадлежность костных находок крупных быков в ряде случаев может быть дискуссионной. Так, костные остатки из археологических памятников SB периода в Казахстане, которые в настоящей работе рассматриваются как принадлежащие туру, другие авторы считают принадлежащими домашнему скоту (Гайдученко, 1998; 2013).
Дикая и домашняя формы лошади не различаются по морфологическим признакам костей скелета, вследствие этого относительно принадлежности лошадей из энеолитических (SB) стоянок Казахстана (ботайско-терсекская историко-культурная общность) имеются противоположные точки зрения. Одни исследователи относят костные остатки лошадей из этих памятников к дикой форме (Ермолова, 1993; Levine, 1999; Benecke, von den Driesch, 2003; Kosintsev, Kuznetsov, 2013), другие – к домашней (Ахинжанов и др., 1992; Гайдученко, 1998; Anthony, Brown, 2000; Outram et al., 2009). В этой связи нами исключены из анализа материалы функционировавших 5000– 4000 л. н. древних поселений с территории Казахстана, в которых кости лошадей найдены совместно с костями домашних животных. Рассматривается материал этого периода только из кратковременных стоянок, оставленных охотниками: здесь количество костных остатков невелико и сопутствующие виды представлены только дикими животными.
Находки костей лошади из энеолитических поселений лесной зоны Полесья и Подвинья (Бибикова, 1972; Кузьмина, 2003), а также лесостепной полосы Урало-Поволжья (Моргунова и др., 2017) отнесены к дикой форме на основании анализа типа хозяйства, культуры, радиоуглеродного возраста и видового состава сопутствующей фауны.
Костные остатки E. lenensis (Russanov 1968) и Equus sp. с территории северо-востока Сибири (Боескоров и др., 2013, 2016) в настоящей работе указаны как принадлежащие E. ferus.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Дикая лошадь (Equus ferus (Boddaert 1785)) пережила переход от плейстоцена к голоцену, хотя ее численность и ареал уменьшились, о чем свидетельствует малочисленность находок (рис. 1A). Некогда единый ареал вида в начале голоцена стал фрагментированным, а в ряде регионов дикая лошадь вымерла.
Рис. 1.
Места находок костных остатков копытных в PB–BO (A) и AT (B) периодах: 1 – европейский осел (Equus hydruntinus), 2 – дикая лошадь (E. ferus), 3 – первобытный бизон (Bison priscus), 4 – гигантский олень (Megaloceros giganteus), 5 – тур (Bos primigenius), 6 – овцебык (Ovibos moschatus). Закрашенные значки обозначают находки, имеющие абсолютный возраст; значки в виде контуров – находки, датированные на основании радиоуглеродных дат слоя и археологическими методами.
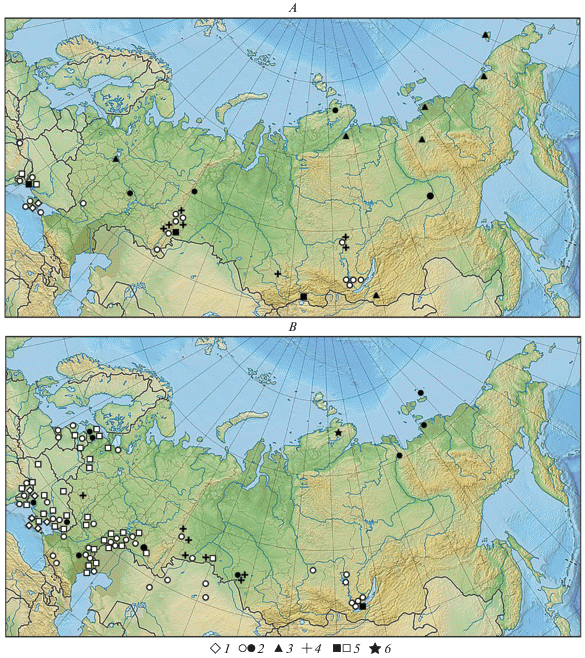
На севере Восточной Европы (Lozovski, 1996; Жилин, 2001, 2002, 2004; Ponomarev et al., 2013; Huftchammer et al., 2018), кроме Прибалтики (Паавер, 1965), Скандинавии (Groves, 1986) и Приморья (Гасилин, 2013), дикая лошадь вымерла на рубеже плейстоцена и голоцена. На большей части Сибири лошадь также исчезла в это время. Исчезновение вида в ряде регионов происходило быстро. Так, в Верхнем Поволжье лошадь уже отсутствует в многочисленных фаунах PB периода (Жилин, 2001, 2002, 2004).
Причиной исчезновения лошади на вышеуказанных территориях, вероятно, стало быстрое формирование лесов, прежде всего хвойных и смешанных (Хотинский, Климанов, 2002; Blyakharchuk, 2003; Väliranta et al., 2006; Novenko et al., 2009; Tarasov et al., 2009). Напротив, в тех районах, где сформировался мозаичный ландшафт, дикая лошадь продолжала обитать. Так, спорово-пыльцевые спектры из вмещающих отложений местонахождений Урала, в которых найдены кости лошади, показывают наличие лесных массивов, но со значительными остепненными участками (Жилин и др., 2012). Радиоуглеродные даты из этих местонахождений получены непосредственно для кости лошади (табл. 1) и для вмещающего слоя (Жилин и др., 2012), что является надежным свидетельством сохранения дикой лошади в некоторых районах лесной зоны.
В раннем голоцене (PB-BO) остатки лошади найдены (рис. 1A) в северном Причерноморье (Бибикова, 1978), Крыму (Мацкевой, 1977; Бибикова, 1984; Benecke, 1999), Карпатах (Мацкевой и др., 1983), Подонье (Трегуб, Федюнин, 2012), Среднем Поволжье (Гугалинская и др., 2010), на Урале (Улитко, 2006; Bachura, Kosintsev, 2007; Жилин и др., 2012), Таймыре (MacPhee et al., 2002), в Приангарье и западном Прибайкалье (Номоконова, Горюнова, 2012; Клементьев, 2014), Северо-Восточной Сибири (Боескоров и др., 2016). К концу раннего голоцена лошадь исчезает в Крыму (Benecke, 1999).
Начиная с AT периода, основная часть ареала дикой лошади занимала территорию современной степной и лесостепной зон (рис. 1B). Подавляющее большинство костных остатков происходит из местонахождений на Нижнем Днестре (Давид, 1982), Нижнем Днепре (Манько и др., 2001; Котова, 2002; Pruvost et al., 2011), Нижнем Дону (Крижевская, 1974; Кузьмина, Каспаров, 1987), в Предкавказье (Спасовский, 2008), в Среднем и Нижнем Поволжье, Прикаспии (Кузьмина, 1988; Косинцев, Варов, 1996; Гасилин и др., 2008; Шишлина и др., 2015; Vybornov et al., 2018), южном Приуралье и Зауралье (Мосин, Страхов, 2010; Моргунова и др., 2017), Западной Сибири (Ахинжанов и др., 1992; Marchenko et al., 2015), Приенисейской Сибири (Мандрыка, Оводов, 2005) и западном Прибайкалье (Ермолова, 1978; Оводов, Панычев, 1982; Номоконова и др., 2009).
Изолированные популяции дикой лошади в этот период сохранялись в лесной зоне (рис. 1B), о чем свидетельствуют находки в Прибалтике (Паавер, 1965; Тимофеев, 1996; Sommer et al., 2011), в Приладожье (Громова, 1949) и на Среднем Урале (Чаиркина, 2005). Другой изолированный участок ареала находился на крайнем северо-востоке Сибири. Костные остатки вида происходят с северной оконечности Якутии и с о-ва Вилькицкого (Kuznetsova et al., 2001; Анисимов и др., 2009; Лазарев и др., 2011; Боескоров и др., 2013). Отсутствие костных находок лошади в центральной части Западной, Средней и Северо-Восточной Сибири может быть следствием слабой изученности района, для которого известно мало местонахождений. Однако надежных свидетельств обитания дикой лошади в таежной зоне нет, поэтому отсутствие находок вида на большей части Сибири наиболее вероятно отражает отсутствие дикой лошади на этой территории в течение голоцена и, следовательно, указывает на изолированность северного фрагмента ее ареала.
К концу AT периода лошадь вымирает на всей территории Сибири, за исключением районов крайнего севера. Самая поздняя находка происходит со Среднего Енисея из отложений стоянки Усть-Шилка 2 возрастом 5800–5220 л. н. (Мандрыка, Оводов, 2005).
Географическое распределение костных находок, датируемых SB периодом (рис. 2A), показало, что основной ареал дикой лошади сохранялся в пределах современных зон степи и лесостепи, простираясь сплошной полосой от северного Причерноморья (Журавлев, 2001), Среднего и Нижнего Поволжья (Кузьмина, 1988), Южного Урала и Зауралья (Мосин, 1996; Косинцев, Пластеева, 2013; Косинцев и др., 2013), Казахстана (Ахинжанов и др., 1992) до Новосибирского Приобья и Предалтайских степей (Ситников и др., 2007; Косинцев и др., 2013). Из местонахождений, расположенных в лесной зоне Русской равнины (Бибикова, 1972; Кузьмина, 2003) и в Прибалтике (Паавер, 1965) находки костей лошади немногочисленны.
Рис. 2.
Места находок костных остатков копытных в SB (A) и SA (B) периодах: 1 – дикая лошадь (Equus ferus), 2 – первобытный бизон (Bison priscus), 3 – тур (Bos primigenius), 4 – овцебык (Ovibos moschatus). Закрашенные значки обозначают находки, имеющие абсолютный возраст; значки в виде контуров – находки, датированные на основании радиоуглеродных дат слоя и археологическими методами.
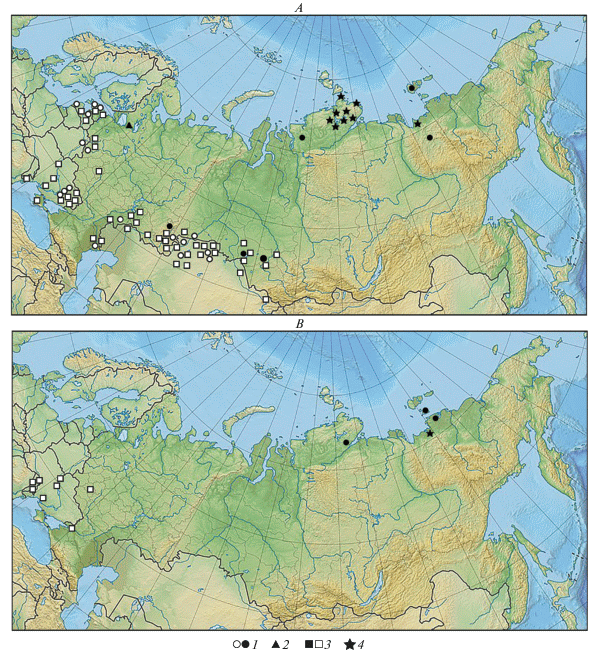
Изолированные популяции дикой лошади в SB период продолжали обитать на крайнем севере Сибири, о чем свидетельствуют прямые даты по костям с о-ва Котельный, северной оконечности Якутии и п-ова Таймыр (Сулержицкий, Романенко, 1997; Кузнецова, ван дер Плихт, 2009; Лазарев и др., 2011).
Точные границы распространения дикой лошади в SA периоде неизвестны по причине широкого распространения домашних лошадей. Письменные исторические источники свидетельствуют, что дикие лошади – тарпаны – еще обитали в евразийских степях до 19 в. (Гептнер и др., 1961; Кириков, 1979, 1983). Вместе с тем, тарпаны свободно скрещивались с домашними лошадьми, т.к. описаны случаи угона дикими жеребцами домашних кобыл (Анучин, 1896).
Восстановленный ареал тарпана занимал степные, лесостепные и полупустынные области Восточной Европы, частично заходя в лесную зону современной Литвы, Белоруссии и Калининградской области (Гептнер и др., 1961), и соответствовал ареалу дикой лошади в SB периоде, за исключением его северо-сибирского фрагмента. От причерноморских степей и Крыма ареал тарпана проходил на восток по низовьям Волги, нижнему и среднему течению Урала и до Барабинской степи и Предалтайских степей включительно (Гептнер и др., 1961). Южная его граница захватывала степи Предкавказья, доходила до Каспийского побережья и, вероятно, до северной оконечности Аральского моря и оз. Балхаш (Гептнер и др., 1961). В литературе известны голоценовые находки диких лошадей из районов, расположенных южнее восстановленной границы ареала тарпана (Кожамкулова, 1969), однако их точный возраст неизвестен.
На крайнем севере Сибири лошадь дожила вплоть до начала нашей эры (табл. 1). Костные остатки в местонахождениях на Хромской губе (Лазарев, 1980), о-ве Большой Ляховский (Kuznetsova et al., 2001) и п-ове Таймыр (Сулержицкий, Романенко, 1997) принадлежат именно дикой форме. Домашняя лошадь появляется в арктической зоне позднее, вместе с якутскими племенами, которые заселили регион между 1500 и 1000 л. н. (Crubézy et al., 2010).
Европейский плейстоценовый осел (Equus hydruntinus (Regalia 1907)). Таксономический статус E. hydruntinus остается дискуссионным (Burke et al., 2003; Orlando et al., 2006; Geigl, Grande, 2012). Молекулярные исследования (Orlando et al., 2006; Bennett et al., 2017) показывают родственные связи между E. hydruntinus и куланом (E. hemionus (Pallas 1775)).
В первой половине голоцена (рис. 1) вид известен из археологических памятников Крыма (Щепинский, Черепанова, 1969; Бибикова, 1975, 1984; Benecke, 1999), нижнего Приднестровья (Бибикова, 1978; Давид, 1982) и северного Приазовья (Котова, 2002), хотя ареал в позднем плейстоцене был шире и охватывал юг Европы до Кавказа (Верещагин, 1959; Громова, 1965; Orlando et al., 2006). Количество костных остатков в изученных местонахождениях голоцена невелико, что косвенно указывает на невысокую численность вида в прошлом.
К середине голоцена европейский осел полностью исчез из фауны региона. Наиболее поздние датированные находки со стоянки Семеновка 1 в Приазовье (рис. 1B) происходят из слоя периода AT 2, т.е. не позднее 6000 л. н. (Котова, 2002). Мало известно об особенностях экологии E. hydruntinus, однако можно предположить, что он имел общие черты с азиатскими ослами (Burke et al., 2003), следовательно, предпочитал открытые ландшафты с засушливым климатом. Вымирание E. hydruntinus, вероятно, определялось изменениями окружающей среды и ростом влажности климата, которые способствовали сокращению и фрагментации ареала в раннем голоцене (Crees, Turvey, 2014). Низкая численность вида, малая площадь ареала и воздействие промысловой деятельности человека привели к тому, что последние представители E. hydruntinus исчезают уже к середине атлантического периода.
Гигантский олень (Megaloceros giganteus (Blumenbach 1799)). В позднем плейстоцене вид обитал от Западной Европы до Забайкалья (Vislobokova, 2012; Lister, Stuart, 2019), однако с самого начала голоцена его распространение и численность значительно сократились.
Фактических данных о распространении гигантского оленя очень немного, поэтому пределы ареала вида на рассматриваемой территории не могут быть очерчены. Известные находки PB–BO периода (рис. 1A) присутствуют в местонахождениях Среднего и Южного Урала (Stuart et al., 2004; Lister, Stuart, 2019), юга Западной Сибири и Приангарья (van der Plicht et al., 2015). Вымирание вида в Приангарье произошло существенно раньше, чем в других регионах, и, вероятно, было связано с замещением степных и лесостепных участков таежной растительностью. Радиоуглеродный возраст находок из Приангарья соответствует 9450–9235 л. н. (van der Plicht et al., 2015), и в отложениях более позднего времени вид уже не встречается.
В начале AT периода рефугиумы гигантского оленя еще сохранялись в центральной части Среднерусской возвышенности (Lister, Stuart, 2019), на Южном Урале и в Зауралье (Stuart et al., 2004; Lister, Stuart, 2019), Барабинской лесостепи (van der Plicht et al., 2015), однако к середине периода вид исчезает из фауны региона. Все голоценовые находки M. giganteus приурочены к лесостепной зоне.
Тур (Bos primigenius (Bojanus 1825)). Для PB–BO периодов известны находки вида из южных областей (рис. 1A): в Приднестровье (Давид, 1982), Поднепровье (Пiдоплiчко, 1956; Котова, 2002; Biagi et al., 2008), южном Зауралье (Мосин, 2000) и в верховьях р. Енисей (Лавров, Забелин, 2007). Точный возраст имеют три находки: кость из древнего поселения Мирное в северном Причерноморье датируется 8280 ± 45 л. н. (Biagi et al., 2008), роговой стержень со стоянки Краснокаменка в южном Зауралье – 8040 ± 210 л. н (Мосин, 2000), скелет тура из отложений р. Барык (Тыва) – 9860 ± 160 л. н. (Лавров, Забелин, 2007). Редкость костных остатков в местонахождениях PB–BO периода косвенно свидетельствует о невысокой численности животного.
Наибольшей численности и обширности своего ареала тур достигает в среднем голоцене, что отражается в росте числа находок костей и местонахождений. В AT периоде (рис. 1B) вид населял лесостепи и лиственные леса, реже – степную зону Европы (Паавер, 1965; Крижевская, 1974; Бибикова, 1975, 1978; Давид, 1982; Кузьмина, Каспаров, 1987; Косинцев, Варов, 1996; Котова, 2002; Гасилин, Косинцев, 2009; Саблин и др., 2011; Галимова и др., 2015; Vybornov et al., 2018) и Сибири до Прибайкалья. Самая восточная находка в Прибайкалье датируется 4966–4940 л. н. (Лозей и др., 2014). В Приморье тур в голоцене не встречался (Гасилин, 2013).
Остеологические материалы свидетельствуют, что в течение SB периода (рис. 2A) тур был представлен на северо-западе Русской равнины (Паавер, 1965; Саблин и др., 2011) и Среднерусской возвышенности (Антипина, 2011), расположенных в подзоне смешанных лесов; в степной и лесостепной зонах северного Причерноморья (Пiдоплiчко, 1956; Журавлев, 2001; Котова, 2002; Секерская, 2010), Поволжья (Гасилин, Косинцев, 2009), Зауралья (Косинцев, 1988; Косинцев, Кисагулов, 2018) и Западной Сибири (Ахинжанов, 1992; Девяшин, Косинцев, 2013). С конца SB периода происходит сокращение ареала вида, в результате которого тур полностью исчезает в лесной (Громова, 1931; Паавер, 1965; Косинцев, Бачура, 2016) и лесостепной (Цалкин, 1963) зонах Восточной Европы, в Западной Сибири (Девяшин, Косинцев, 2013).
Немногочисленные костные остатки вида, относящиеся к началу SA, отмечены в археологических памятниках Причерноморья (Пiдоплiчко, 1956) и Приазовья (Благуш, 2013), тогда как на остальной территории тур уже исчез (рис. 2B). Для более позднего времени – от начала нашей эры до конца эпохи средневековья – находки тура описаны только из археологических поселений Приднестровья (Давид, 1982), Поднепровья (Пiдоплiчко, 1956), Полесья (Александрович, 1999) и Среднерусской возвышенности (Чубур, Радюш, 2013). Поскольку в слоях этих поселений предполагаемые кости скелета тура найдены вместе с костями зубра и крупного рогатого скота, результаты видовой идентификации нуждаются в специальной проверке. В соседних регионах южной Европы позднее 1500 л. н. тур не встречался (Bökönyi, 1974). Общеизвестное упоминание об охоте Владимира Мономаха на “туров” в черниговских землях (Повесть временны́х лет, 1996) в начале 2-го тысячелетия н.э. не может служить доказательством присутствия зверя, т.к. не ясно, какой из видов животных имелся в виду – зубр или тур.
На остальной территории Восточной Европы костных остатков тура, за исключением отдельных роговых чехлов (Цалкин, 1952; Паавер, 1965), которые могли использоваться в качестве изделий и сувениров, в археологических памятниках не найдено. Отсутствие тура в изучаемом регионе в этот период подкрепляется также историческими свидетельствами, согласно которым на протяжении 10–19 вв. в лесостепной и степной зоне Восточной Европы тур уже не обитал, а зубр был представлен (Кириков, 1979, 1983). Последние туры сохранялись в малонаселенных человеком лесах севера Европы до конца средневековья. Письменные источники указывают на обитание вида в средние века в Литве (Кириков, 1966) и выживании зверя на границе Литвы и Польши вплоть до 17 в. (van Vuure, 2005).
Первобытный бизон (Bison priscus (Bojanus 1825)) был одним из наиболее многочисленных копытных животных Северной Евразии в позднем плейстоцене, однако к концу BO периода голоцена вымер на большей части прежнего ареала (рис. 1A). Изолированные популяции бизонов в PB-BO сохранялись на северо-западе европейской части (Hartz et al., 2010), на крайнем севере Сибири (MacPhee et al., 2002; Vartanyan et al., 2008; Боескоров и др., 2013; Kirillova et al., 2015; Григорьев и др., 2017), а также в Забайкалье (Зарецкая и др., 2015). Все известные кости бизона имеют абсолютный возраст (табл. 1). Представленная в литературе радиоуглеродная дата трубчатой кости B. priscus с р. Орда (Новосибирское Приобье), 9320 ± 95 л. н., СОАН–4568 (Васильев и др., 2007), должна рассматриваться с осторожностью, т.к. достоверность видового определения этой находки невозможно проверить.
Необходимо отметить, что малочисленность находок костей голоценового бизона может быть связана с трудностью морфологического разграничения остатков Bos и Bison, а также с немногочисленностью костного материала периода раннего голоцена в целом.
Для азиатской части региона достоверные находки остатков бизона более позднего возраста неизвестны, а для европейской – имеется сделанная на северо-западе региона, на р. Оять (рис. 2A), единственная находка черепа бизона SB периода, абсолютный возраст которой составил 3045 ± ± 35 л. н. (Markova et al., 2015). Фрагменты роговых стержней бизона из неолитических стоянок и одного средневекового поселения в Прибайкалье (Ермолова, 1978), фрагменты черепа, рогового стержня и зубов из неолитической стоянки в северном Приангарье (Васильев и др., 2013) в настоящей работе не учтены. Абсолютный возраст находок из Приангарья неизвестен, а поздний возраст костных остатков из Прибайкалья не может быть подтвержден (Пластеева, Клементьев, 2018).
Овцебык (Ovibos moschatus (Zimmermann 1780)). Все известные костные находки овцебыка датируются периодами AT, SB, SA и происходят с территории, в настоящее время расположенной за пределами северного полярного круга: на п-ове Таймыр (Сулержицкий, Романенко, 1997; Kuznetsova et al., 2001; MacPhee et al., 2002; Campos et al., 2010; Lorenzen et al., 2011) и на севере Якутии (Kuznetsova et al., 2001; Boeskorov et al., 2017). Следует предположить, что животные обитали здесь непрерывно с позднего плейстоцена. На остальной части исследуемого региона вид вымер еще в конце плейстоцена, и последними географическими областями, оставленными видом, были Западная Европа и Урал (Markova et al., 2015).
Главным фактором в вымирании овцебыка является изменение климата (Lorenzen et al., 2011). Значительное сокращение его ареала в Евразии началось еще в конце плейстоцена (Markova et al., 2015), а в голоцене последние представители вида сохранялись лишь на территории современной арктической зоны благодаря наличию оптимальных условий обитания.
ЗАКЛЮЧЕНИЕ
Начавшиеся в конце плейстоцена–начале голоцена изменения природной среды привели к сокращению ареалов дикой лошади, первобытного бизона, овцебыка, европейского осла и гигантского оленя в Северной Евразии. Снижение численности и распространения у разных копытных животных произошло в раннем голоцене, а у некоторых видов началось еще в конце позднего плейстоцена. Реакция изученных видов на изменения внешней среды была неодинаковой.
Плейстоценовые ареалы широко распространенных видов – дикой лошади и первобытного бизона – в раннем голоцене распались на отдельные фрагменты. Южная граница ареала бизона достаточно быстро отступила к северу, а к концу бореального периода бизоны исчезли в азиатской части субконтинента, сохранившись лишь в его европейской части. Локальное вымирание дикой лошади в раннем голоцене выразилось в разрыве некогда единого ареала. В последующие периоды дикая лошадь обитала в двух географически разделенных районах: на территории современной Арктики, где долгое время сохранялись перигляциальные ландшафты, и в зоне евразийских степей и лесостепей. Части ареала были географически изолированы таежной зоной, сформировавшейся в центральной Сибири с началом голоцена (Хотинский, Климанов, 2002).
Таким образом, наиболее экологически пластичные виды копытных животных, как дикая лошадь и первобытный бизон, в начале голоцена сократили свою численность и распространение, однако впоследствии дикая лошадь сохранилась в степной зоне, а первобытный бизон вымер в Северной Евразии. В течение первой половины голоцена дикая лошадь обитала не только в открытых степных пространствах, но и на территории, где произрастали широколиственные или светлые смешанные леса, обеспечивавшие необходимый мозаичный ландшафт. Причинами постепенного сокращения численности последних диких лошадей, вплоть до их полного исчезновения, явилось прямое истребление человеком и сельскохозяйственное освоение основных биотопов – степей (Кириков, 1979, 1983). Последние тарпаны продержались в лесостепной и степной зонах Причерноморья и Урало-Поволжья до второй половины 19 в. (Кириков, 1983).
Из бизонов в голоцене вымер лишь первобытный бизон, имевший к его началу наибольший ареал. Родственные бизонам зубры дожили до исторического времени в закрытых ландшафтах Центральной Европы и Кавказа (Гептнер и др., 1961; Benecke, 2005).
Существенное уменьшение и фрагментацию ареалов претерпели также относительно специализированные виды открытых и полуоткрытых биотопов, которые имели узкое распространение на юге региона – европейский осел и гигантский олень. Их исчезновение произошло уже в первой половине голоцена. Оба вида были сравнительно немногочисленны в голоцене, имели ограниченное распространение и являлись объектами добычи древнего человека, что делало их более уязвимыми.
Единственным видом, который расширил свой ареал в голоцене, является тур. К середине периода тур достигает наибольшей численности и географического распространения в регионе, встречаясь в лесной и лесостепной зонах Европы и Сибири. Однако уже в начале нашей эры тур исчезает с большей части своего ареала. Решающим фактором вымирания тура в природе, вероятно, стал антропогенный, влияние которого заметно усилилось с переходом человеческих коллективов умеренной полосы Северной Евразии к производящему типу хозяйства в эпоху неолита. Воздействие на оставшиеся популяции животных проявлялось как в непосредственном истреблении животных, так и в изменении мест их обитания посредством разрушения привычных биотопов и конкуренции с домашним скотом (Гептнер и др., 1961).
Можно заключить, что реакция изученных копытных животных на комплекс ландшафтно-климатических изменений возникала в разное время и протекала с различной интенсивностью – вероятно, в соответствии с влиянием конкретных лимитирующих факторов. Каждый из видов в изученной группе имеет свою историю вымирания: от начала снижения численности, характера сокращения и фрагментации ареала до географического положения и биотопического характера рефугиумов, где вымирали последние популяции. Выяснение причин исчезновения крупных травоядных млекопитающих требует не только детального изучения нового и имеющегося фаунистического материала, но и привлечения широкого спектра палеоэкологических данных.
Список литературы
Александрович Н.П., 1999. Остеологическая оценка географической и хронологической изменчивости охотничьих млекопитающих средневековой Беларуси. Автореф. дис. … канд. биол. наук. Минск. 19 с.
Анисимов М.А., Иванова В.В., Пушина З.В., Питулько В.В., 2009. Лагунные отложения острова Жохова: возраст, условия формирования и значение для палеогеографических реконструкций региона Новосибирских островов // Известия АН. Серия географическая. № 5. С. 107–119.
Антипина Е.Е., 2011. Археозоологические коллекции из поселений поздней бронзы на территории Тульской области. Приложение 3 // Гак Е.И. Поселения эпохи бронзы на северной окраине донской лесостепи. М.: ГИМ. С. 166–201.
Анучин Д.Н., 1896. К вопросу о диких лошадях и об их приручении в России // Журнал Министерства народного просвещения. С. 224–225.
Ахинжанов С.М., Макарова С.М., Нурумов Т.Н., 1992. К истории скотоводства и охоты в Казахстане. Алма-Ата: Гылым. 218 с.
Бибикова В.И. 1958. О некоторых отличительных чертах в костях конечностей зубра и тура // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 63. №. 6. С. 23–35.
Бибикова В.И., 1972. Фауна из неолитических поселений северо-восточной Белоруссии // Природная обстановка и фауны прошлого. № 6. Киев: Наукова Думка. С. 51–70.
Бибикова В.И., 1975. О смене некоторых компонентов фауны копытных на Украине в голоцене // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 80. № 6. С. 67–72.
Бибикова В.И., 1978. Фауна из мезолитических поселений Белолесье и Гиржево (Нижнее Поднестровье) // Археологические исследования северо-западного Причерноморья. Киев. С. 17–29.
Бибикова В.И., 1984. Копытные Северного Причерноморья в раннем голоцене (по материалам археологических раскопок) // Проблемы изучения истории современных биогеоценозов. М.: ИЭМЭЖ. С. 171–203.
Благуш А.А., 2013. Некоторые данные к изучению видового разнообразия млекопитающих Елизаветинского городища (раскопки 2008–2011 гг.) // Историко-археологические исследования в Азове и на Нижнем Дону в 2011 г. С. 146–148.
Боескоров Г.Г., Протопопов А.В., Мащенко Е.Н., Потапова О.Р., Кузнецова Т.В. и др., 2013. Новые находки ископаемых млекопитающих уникальной сохранности в многолетней мерзлоте Якутии // Доклады Академии Наук. Т. 452. № 4. С. 461–465.
Боескоров Г.Г., Ноговицын П.Р., Мащенко Е.Н., Белолюбский И.Н., Степанов А.Д., и др., 2016. Новые данные о млекопитающих мамонтовой фауны бассейна Средней Лены (Якутия; Национальный природный парк “Ленские Столбы” и прилегающие территории) // Доклады Академии Наук. Т. 469. № 2. С. 190–194.
Васильев С.К., Орлова Л.А., Кузьмин Я.В., 2007. Местонахождение фауны крупных млекопитающих сартанского времени на реке Орда (Ордынский р-н Новосибирской области) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. XIII. С. 29–32.
Васильев С.К., Славинский В.С., Постнов А.В., 2013. Гигантский олень (Megaloceros giganteus Blumenbach, 1803) в палеофауне голоценовых стоянок Северного Приангарья (Усть-Тушама-1, Остров Сосновый Тушамский, Усть-Талая) // Вестник Новосибирского университета. Серия: История, филология. Т. 12. № 7. С. 177–185.
Верещагин Н.К., 1959. Млекопитающие Кавказа: история формирования фауны. М., Л.: Изд-во АН СССР. 703 стр.
Гайдученко Л.Л., 1998. Домашняя лошадь и крупный рогатый скот поселения Кожай I // Поселение Кожай I. Алматы: ИА РК. С. 234–254.
Гайдученко Л.Л., 2013. Древнейшие формы крупного рогатого скота Урало-Казахстанских степей // Этнические взаимодействия на Южном Урале: сборник науч. трудов. Челябинск: Рифей. С. 261–268.
Галимова Д.Н., Аськеев И.В., Аськеев О.В., Гусенцова Т.М., 2015. Археозоологический анализ неолитической стоянки Подолье I в южном Приладожье // Неолитические культуры Восточной Европы: хронология, палеоэкология, традиции. СПб. С. 163–166.
Гасилин В.В., 2013. Крупные млекопитающие Приморья в голоцене // Зоологический журнал. Т. 92. № 9. С. 1055–1063.
Гасилин В.В., Косинцев П.А., 2009. Динамика структуры фауны копытных Урало-Поволжья в голоцене // Вестник Оренбургского университета. № 6. С. 108–110.
Гасилин В.В., Косинцев П.А., Саблин М.В., 2008. Фауна неолитической стоянки Варфоломеевская в степном Поволжье // Фауны и флоры Северной Евразии в позднем кайнозое. Екатеринбург–Челябинск: Рифей. № 6. С. 25–100.
Гептнер В.Г., Насимович А.А., Банников А.Г., 1961. Млекопитающие Советского Союза. Т. 1. Парнокопытные и непарнокопытные. М.: Высшая школа. С. 700–707.
Григорьев С.Е., Чепрасов М.Ю., Саввинов Г.Н., Тихонов А.Н., Новгородов Г.П. и др., 2017. Палеонтологические и археозоологические исследования в бассейне р. Яна // Вестник Северо-Восточного Федерального университета. Т. 1 (57). С. 20–35.
Громова В.И., 1931. Первобытный бык или тур (Bos primigenius Воj.) в СССР // Ежегодник Зоологического музея АН СССР. Т. XXXII. С. 293–364.
Громова В.И., 1949. История лошадей (рода Equus) в Старом Свете // Труды Палеонтологического института. Т. XVII. № 1. С. 191–192.
Громова В.И., 1965. Краткий обзор четвертичных млекопитающих Европы (опыт сопоставления). М.: Наука. 143 с.
Гугалинская Л.А., Алифанов В.М., Березина Н.С., Березин А.Ю., Хисяметдинова А.А. и др., 2010. Шолмская погребенная почва на финальнопалеолитическом поселении Шолма-1 (Приволжская возвышенность, Чувашское плато) // Бюллетень комиссии по изучению четвертичного периода. № 70. С. 45–58.
Давид А.И., 1982. Формирование териофауны Молдавии в антропогене. Кишинев: Штиинца. 151 с.
Девяшин М.М., Косинцев П.А., 2013. Копытные млекопитающие среднего и позднего голоцена юго-востока Западной Сибири // Вестник Томского государственного университета. Биология. № 2 (22). С. 127–140.
Ермолова Н.М., 1978. Териофауна долины Ангары в позднем антропогене. Новосибирск: Наука. 222 с.
Ермолова Н.М., 1993. Остатки млекопитающих поселения Ботай (по раскопкам 1982 года) // Проблемы реконструкции хозяйства и технологий по данным археологии. Петропавловск: Отдел “Археология Северного Казахстана” ИА НАН РК. С. 87–89.
Жилин М.Г., 2001. Костяная индустрия мезолита лесной зоны Восточной Европы. М. 328 с.
Жилин М.Г., 2002. Охота и рыболовство в мезолите Волго-Окского междуречья (по материалам торфяниковых поселений) // Северный Археологический Конгресс. Ханты-Мансийск. С. 112–122.
Жилин М.Г., 2004. Природная среда и хозяйство мезолитического населения центра и северо-запада лесной зоны Восточной Европы. М.: Academia. 141 с.
Жилин М.Г., Савченко С.Н., Сериков Ю.Б., Косинская Л.Л., Косинцев П.А., 2012. Мезолитические памятники Кокшаровского торфяника. М. 167 с.
Журавлев О.П., 2001. Остеологические материалы из памятников эпохи бронзы лесостепного Днепро-Донского междуречья. Киев: ИА НАНУ. 200 с.
Зарецкая Н.Е., Кобылкин Д.В., Косинцев П.А., Максимов Ф.Е., Рыжов Ю.В. и др., 2015. Новые данные о голоценовом возрасте бизонов (Bison priscus) в Забайкалье // Фундаментальные проблемы квартера. Итоги изучения и основные направления дальнейших исследований: Материалы IX Всероссийского совещания по изучению четвертичного периода. Иркутск: ИГ СО РАН. С. 162.
Кириков С.В., 1966. Промысловые животные, природная среда и человек. М.: Наука. 348 с.
Кириков С.В., 1979. Человек и природа восточно-европейской лесостепи в X–начале XIX в. М.: Наука. 183 с.
Кириков С.В., 1983. Человек и природа степной зоны. Конец X–середина XIX в. M.: Наука. 126 с.
Клементьев А.М., 2014. Раннеголоценовая фауна северной Ангары (материалы археологических объектов) // Известия Иркутского Государственного Университета. Серия Геоархеология. Этнология. Антропология. Т. 8. С. 31–44.
Кожамкулова Б.С., 1969. Антропогеновая ископаемая териофауна Казахстана. Алма-Ата: Институт зоологии АН Казахской ССР. 149 с.
Косинцев П.А., 1988. Голоценовые остатки крупных млекопитающих Западной Сибири // Современное состояние и история животного мира Западносибирской низменности. Свердловск: Изд-во УрО АН СССР. С. 32–51.
Косинцев П.А., Бачура О.П., 2016. Фауна крупных млекопитающих центра Восточно-Европейской равнины в голоцене // Динамика современных экосистем в голоцене: Материалы Всероссийской конференции. М.: КМК. С. 99–102.
Косинцев П.А., Варов А.И., 1996. Ранние этапы животноводства в Волго-Уральском регионе // Взаимодействие человека и природы на границе Европы и Азии. Самара. С. 29–31.
Косинцев П.А., Кисагулов А.В., 2018. Костные остатки тура и крупного рогатого скота из голоценовых местонахождений южного Зауралья // Вестник Оренбургского Государственного педагогического университета. № 1 (25). С. 44–55.
Косинцев П.А., Пластеева Н.А., 2013. Ареалы голоценовой дикой лошади (Equus ferus Boddaert, 1785) в Северной Евразии // Динамика современных экосистем в голоцене: Материалы 3-й Всероссийской научной конференции (с международным участием). Казань: Отечество. С. 199–202.
Косинцев П.А., Пластеева Н.А., Васильев С.К., 2013. Дикие лошади (Equus (Equus) s.l.) Западной Сибири в голоцене // Зоологический журнал. Т. 92. № 9. 2013. С. 1107–1116.
Котова Н.С., 2002. Неолитизация Украины. Луганск: Шлях. 268 с.
Крижевская Л.Я., 1974. К вопросу о формах хозяйства неолитического населения северо-восточного Приазовья // Первобытный человек и природная среда. М. С. 263–268.
Кузнецова Т.В., ван дер Плихт Х., 2009. Позднеплейстоценовые и голоценовые лошади северной Якутии // 200 лет отечественной палеонтологии: материалы конференции. М. С. 65–66.
Кузьмина И.Е., 1988. Млекопитающие Северного Прикаспия в голоцене // Археологические культуры Северного Прикаспия: сборник научных трудов. Куйбышев. С. 173–188.
Кузьмина С.А., 2003. Новые фаунистические данные по результатам раскопок неолитических памятников Смоленской и Псковской областей // Древности Подвинья: исторический аспект. СПб: Изд-во Государственного Эрмитажа. С. 305.
Кузьмина И.Е., Каспаров А.К., 1987. Остатки животных их неолитических стоянок Копанище и Черкасская в Воронежской области // Труды Зоологического института АН СССР. Т. 168. С. 87–99.
Лавров А.В., Забелин В.И., 2007. Первобытный тур (Bos primigenius, Bovidae) в раннем голоцене республики Тывы // Фундаментальные проблемы квартера: итоги изучения и основные направления дальнейших исследований. Материалы V Всероссийского совещания по изучению четвертичного периода. М.: ГЕОС. С. 208–209.
Лазарев П.А., 1980. Антропогеновые лошади Якутии. М.: Наука. 189 с.
Лазарев П.А., Григорьев С.Е., Плотников В.В., Саввинов Г.Н., 2011. Находки уникальных останков туш лошади и бизона в Верхоянском районе Якутии // Проблемы региональной экологии. Т. 4. С. 13–18.
Лозей Р.Дж., Номоконова Т.Ю., Савельев Н.А., 2014. Радиоуглеродное датирование и фауна многослойной стоянки Бугульдейка II на Байкале (по материалам раскопок 2006–2008 гг.) // Известия Иркутского государственного университета. Серия Геоархеология. Этнология. Антропология. Т. 7. С. 18–36.
Мандрыка П.В., Оводов Н.Д., 2005. Археология и фауна мезолитического комплекса Усть-Шилка-2 на Среднем Енисее // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. XI. Ч. I. С. 157–162.
Манько В.А., Телиженко С.А., Журавлев О.П., Ковалюх Н.Н., 2001. Предварительные итоги исследования узла неолитических памятников у озера Туба // Древности Северского Донца. Луганск: Шлях. С. 27–43.
Мацкевой Л.Г., 1977. Мезолит и неолит Восточного Крыма. Киев: Наукова Думка. 180 с.
Мацкевой Л.Г., Адаменко О.М., Пашкевич Г.А., Татаринов К.А., 1983. Природная среда и мезолит запада Украины // Советская археология. № 1. С. 5–29.
Моргунова Н.Л., Васильева И.Н., Кулькова М.А., Рослякова Н.В., Салугина Н.П. и др., 2017. Турганикское поселение в Оренбургской области. Оренбург: Издательский центр ОГАУ. С. 221–232.
Мосин В.С., 1996. Стоянка Бурли II и некоторые вопросы энеолита Южного Зауралья // Новое в археологии Южного Урала. Челябинск: Рифей. 265 с.
Мосин В.С., 2000. Зауральская культурно-историческая область в мезолите–энеолите // Древняя история Южного Зауралья. Т. 1. Челябинск: Изд-во ЮУрГУ. С. 235.
Мосин В.С., Страхов А.Н., 2010. Нео-энеолитические комплексы поселения Кочегарово I // Уральское археологическое совещание. Т. XVIII. Уфа: Изд-во БГПУ. С. 111–112.
Номоконова Т.Ю., Горюнова О.И., 2012. Фаунистические остатки из многослойного поселения Саган-Нугэ (озеро Байкал) // Проблемы археологии, антропологии и этнографии Сибири и сопредельных территорий. Т. 18. С. 123–127.
Номоконова Т.Ю., Лозей Р.Дж., Горюнова О.И., 2009. Фауна многослойного поселения Берлога (Малое море, озеро Байкал) // Проблемы археологии, антропологии и этнографии Сибири и сопредельных территорий. Т. 15. С. 117–181.
Оводов Н.Д., Панычев В.А., 1982. Фауна и радиоуглеродное датирование неолитической стоянки Саган-Заба на Байкале // Проблемы археологии и этнографии Сибири: Тезисы докладов региональной конференции. Иркутск: Изд-во ИГУ. С. 66–68.
Паавер К.Л., 1965. Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту. 449 с.
Пiдоплiчко I.Г., 1956. Матерiали до вивчення минулих фаун УРСР. Вип. 2 Киiв. 189 с.
Пластеева Н.А., Клементьев А.М., 2018. Крупные млекопитающие Прибайкалья в голоцене // Зоологический журнал. Т. 97. № 5. С. 599–612.
Повесть временны́х лет, 1996. Подгот. текста, пер., ст. и коммент. Д.С. Лихачева; Под ред. Адриановой-Перетц В.И.; [Доп. М.Б. Свердлова]. 2-е изд., испр. и доп. СПб.: Наука. 667 с.
Саблин М.В., Пантелеев А.В., Сыромятникова Е.В., 2011. Археозоологический анализ остеологического материала из неолитических свайных поселений Подвинья: хозяйство и экология // Труды Зоологического института РАН. Т. 315. № 2. С. 143–153.
Секерская Е.П., 2010. Особенности палеоэкономической стратегии населения Нижнего Дуная в эпоху позднего энеолита – ранней бронзы // Материалы по археологии Северного Причерноморья. Вып. 11. С. 141–162.
Ситников С.М., Васильев С.К., Кирюшин К.Ю., 2007. Анализ фаунистических остатков с поселения Новоильинка III // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 11. С. 363–366.
Спасовский Ю.Н., 2008. Результаты определения остеологических сборов из поселений майкопской культуры “Новосвободненское” и “Пхагуапе” // Археология Кавказа и Ближнего Востока. М.: ТАУС. С. 256–258.
Сулержицкий Л.Д., Романенко Ф.А., 1997. Возраст и расселение мамонтовой фауны азиатского Заполярья // Криосфера Земли. № 1. С. 12–19.
Тимофеев В.И., 1996. Памятники типа Цедмар // Археология. Неолит Северной Евразии. Отв. ред. Ошибкина С.В. М.: Наука. С. 162–166.
Трегуб Т.Ф., Федюнин И.В., 2012. Плаутино 2: Некоторые вопросы геоморфологии и палеогеографии / Первобытные древности Евразии. К 60-летию А.Н. Сорокина. М.: ИА РАН. С. 275.
Улитко А.И., 2006. Голоценовые млекопитающие из карстовых полостей Среднего Урала // Динамика современных экосистем в голоцене: материалы Всероссийской научной конференции. М.: Товарищество научных изданий КМК. С. 243–247.
Хотинский Н.А., Алешинская З.В., Гуман М.А., Климанов В.А., Черкинский А.Е., 1991. Новая схема периодизации ландшафтно-климатических изменений в голоцене // Известия АН СССР. Серия географическая. № 3. С. 30–42.
Хотинский Н.А., Климанов В.А., 2002. Растительность голоцена // Динамика ландшафтных компонентов и внутренних морских бассейнов Северной Евразии за последние 130 000 лет. Атлас-монография. Отв. ред. Величко А.А. М.: ГЕОС. С. 89–104.
Цалкин В.И., 1952. Новая находка остатков первобытного быка в СССР // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. LXVII. № 2. С. 81–83.
Цалкин В.И., 1963. К истории млекопитающих восточноевропейской лесостепи // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. LXVIII. № 2. С. 48–62.
Цалкин В.И., 1970. Древнейшие домашние животные Восточной Европы. М.: Наука. 280 с.
Чаиркина Н.М., 2005. Энеолит Среднего Зауралья. Екатеринбург: ИИА УрО РАН. 313 с.
Чубур А.А., Радюш О.А. К зооархеологической характеристике Новосильского городища // Вопросы археологии, истории, культуры и природы Верхнего Поочья. Материалы XV Всеросс. науч. конф. Калуга: Фридгельм, 2013. С. 53–57.
Шишлина Н.И., Борисов А.В., Клещенко А.А., Идрисов И.А., Чернышева Е.В., ван дер Плихт Й., 2015. Стоянка Большая Элиста 1. Теория и практика поиска и изучения поселений древних скотоводов в пустынно-степной зоне // Известия Самарского научного центра РАН. Т. 17. № 3. С. 252–261.
Щепинский А.А., Черепанова Е.Н., 1969. Северное Присивашье в V–I тысячелетии до нашей эры. Симферополь. С. 36–38.
Anthony D., Brown D., 2000. Eneolithic horse exploitation in the Eurasian steppes: diet, ritual and riding // Antiquity. V. 74. P. 75–86.
Bachura O., Kosintsev P., 2007. Late Pleistocene and Holocene small- and large-mammal faunas from the Northern Urals // Quaternary International. V. 160. P. 121–128.
Benecke N., 1999. The evolution of the vertebrate fauna in the Crimean Mountains from the Late Pleistocene to the mid-Holocene // The Holocene history of the European vertebrate fauna: Modern aspects of research. Leidorf, Berlin. P. 43–57.
Benecke N., 2005. The Holocene distribution of European bison – the archaeozoological record // Munibe (Antropologia-Arkeologia). V. 57. P. 421–428.
Benecke N., von den Dreisch A., 2003. Horse exploitation in the Kazakh steppes during the Eneolithic and Bronze Age // Prehistoric steppe adaptation and the horse. Cambridge: McDonald Institute for archaeological research. P. 69–82.
Bennett E., Champlot S., Peters J., Arbuckle B., Guimaraes S. et al., 2017. Taming the late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA // PLOS One. V. 22 (4). P. e0174216. https://doi.org/10.1371/journal.pone.0174216
Biagi P., Stanko V.N., Kiosak D.V., 2008. New radiocarbon dates for Myrne settlement // Naukovi praci. V. 96. № 83. P. 33–37.
Binney H.A., Willis K.J., Edwards M.E., Bhagwat Sh.A., Anderson P.M., 2009. The distribution of late-Quaternary woody taxa in northern Eurasia: evidence from a new macrofossil database // Quaternary Science Reviews. V. 28. P. 2445–2464.
Blyakharchuk T.A., 2003. Four new pollen sections tracing the Holocene of the southern part of the West Siberian Lowland // The Holocene. V. 13. № 5. P. 715–731.
Boeskorov G., Plotnikov V., Protopopov A., Klimovskiy A., Kolesov S., van der Plicht J., 2017. New finding of a Holocene muskox (Ovibos pallantis H. Smith, 1827) In Yakutia (East Siberia, Russia) // VII International conference on mammoths and their relatives. P. 14.
Bökönyi S., 1974. History of domestic mammals in central and eastern Europe. Budapest: Akademiai Kiadó. 597 p.
Burke A., Eisenmann V., Ambler G.K., 2003. The systematic position of Equus hydruntinus, an extinct species of Pleistocene equid // Quaternary Research. V. 59. P. 459–469.
Campos P.F., Willerslev E., Sher A., Orlando L., Axelsson E. et al., 2010. Ancient DNA analyses exclude humans as the driving force behind late Pleistocene musk ox (Ovibos moschatus) population dynamics // Proceedings of the National academy of sciences. V. 107. P. 5675–5680.
Cooper A., Turney Ch., Hughen K.A., Brook B.W., McDonald H.G., Bradshaw C.J.A., 2015. Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover // Science. V. 349. № 6248. P. 602–606.
Crees J., Carbone C., Sommer R., Benecke N., Turvey S., 2016. Millennial-scale faunal record reveals differential resilience of European large mammals to human impacts across the Holocene // Proceedings of the Royal Society. Biological Sciences. V. 283. P. 2015–2152.
Crees J., Turvey S., 2014. Holocene extinction dynamics of Equus hydruntinus, a late-surviving European megafaunal mammal // Quaternary Science Review. V. 91. P. 16–29.
Crubézy E., Amory S., Keyser C., Bouakaze C., Bodner M. et al., 2010. Human evolution in Siberia: from frozen bodies to ancient DNA // BMC Evolutionary Biology. V. 10. № 25. https://doi.org/10.1186/1471-2148-10-25
Geigl E.-M., Grange T., 2012. Eurasian wild asses in time and space: morphological versus genetic diversity // Annals of anatomy. V. 194. P. 88–102.
Groves C.P., 1986. The taxonomy, distribution and adaptations of recent Equids // Equids in the Ancient World. Wiesbaden: Dr Ludwig Reichert Verlag. P. 11–65.
Guthrie D.R., 2006. New carbon dates link climatic change with human colonization and Pleistocene extinctions // Nature. V. 441. P. 207–209.
Hartz S., Terberger T., Zhilin M., 2010. New AMS-dates for the Upper Volga Mesolithic and the origin of microblade technology in Europe // Quartar. V. 57. P. 155–169.
Hufthammer A.K., Svendsen J.I., Pavlov P., 2018. Animals and humans in the European Russian Arctic towards the end of the last Ice Age and during the mid-Holocene time // Boreas. https://doi.org/10.1111/bor.12343
Kirillova I.V., Zanina O.G., Chernova O.F., Lapteva E.G., Trofimova S.S. et al., 2015. An ancient bison from the mouth of the Rauchua river (Chukotka, Russia) // Quaternary Research. V. 84. P. 232–245.
Kosintsev P., Kuznetsov P., 2013. Comment on “The earliest horse harnessing and milking” // Tyragetia. V. 7. P. 405–408.
Kuznetsova T.V., Sulerzhitsky L.D., Siegert Ch., 2001. New data on the “mammoth” fauna of the Laptev Shelf Land (Arctic Siberia) // Proceedings of the First International Congress “The World of Elephants”. Rome: Consiglio Nazionale delle Ricerche. P. 289–292.
Levine M.A., 1999. Botai and the origins of horse domestication // Journal of Anthropology and Archaeology. V. 18. P. 29–78.
Lister A.M., Stuart A.J., 2008. The impact of climate change on large mammal distribution and extinction: evidence from the last glacial / interglacial transition // C. R. Geoscience. V. 340. P. 615–620.
Lister A.M., Stuart A.J., 2019. The extinction of the giant deer Megaloceros giganteus (Blumenbach): New radiocarbon evidence // Quaternary International. V. 500. P. 185–203.
Lorenzen E.D., Nogue’s-Bravo D., Orlando L., Weinstock J., Binladen J. et al., 2011. Species-specific responses of Late Quaternary megafauna to climate and humans // Nature. V. 479. P. 359–365.
Lozovski V.M., 1996. Zamostje 2: les derniers chasseurs-pêcheurs préhistoriques de la plaine russe: the last prehistoric hunter-fishers of the Russian plain. Cedarc, Treignes. 96 p.
MacDonald G.M., Velichko A.A., Kremenetski C.V., Borisova O.K., Goleva A.A. et al., 2000. Holocene treeline history and climate change across Northern Eurasia // Quaternary Research. V. 53. P. 302–311.
MacPhee R.D.E., Tikhonov A.N., Mol D., de Marliave C., van der Plicht H. et al., 2002. Radiocarbon chronologies and extinction dynamics of the Late Quaternary mammalian megafauna of the Taimyr Peninsula, Russian Federation // Journal of Archaeological Science. V. 29. P. 1017–1042.
Marchenko Z.V., Orlova L.A., Panov V.S., Zubova A.V., Molodin V.I. et al., 2015. Paleodiet, radiocarbon chronology, and the possibility of fresh-water reservoir effect for Preobrazhenka 6 burial ground, Western Siberia: preliminary results // Radiocarbon. V. 57. № 4. P. 595–610.
Markova A.K., Puzachenko A.Yu., van Kolfschoten T., Kosintsev P.A., Kuznetsova T.V. et al., 2015. Changes in the Eurasian distribution of the musk ox (Ovibos moschatus) and the extinct bison (Bison priscus) during the last 50 ka BP // Quaternary International. V. 378. P. 99–110.
Novenko E.Yu., Volkova E.M., Nosova N.B., Zuganova I.S., 2009. Late Glacial and Holocene landscape dynamics in the southern taiga zone of East European Plain according to pollen and macrofossil records from the Central Forest State Reserve (Valdai Hills, Russia) // Quaternary International. V. 207. P. 93–103.
Olsen S.J., 1960. Postcranial skeletal characters of Bison and Bos // Papers of the Peabody Museum of Archaeology and Ethnology, Harvard University. V. 35. № 4. P. 1–15.
Orlando L., Mashkour M., Burke A., Douady C.J., Eisenmann V., Hänni C., 2006. Geographic distribution of an extinct equid (Equus hydruntinus: Mammalia, Equidae) revealed by morphological and genetical analyses of fossils // Molecular Ecology. V. 15. P. 2083–2093.
Outram A.K., Stear N.A., Bendrey R., Olsen S., Kasparov A. et al., 2009. The earliest horse harnessing and milking // Science. V. 323. P. 1332–1335.
Plicht J. van der, Molodin V.I., Kuzmin Y.V., Vasiliev S.K., Postnov A.V., Slavinsky V.S., 2015. New Holocene refugia of giant deer (Megaloceros giganteus Blum.) in Siberia: updated extinction patterns // Quaternary Science Reviews. V. 114. P. 182–188.
Ponomarev D., Puzachenko A., Bachura O., Kosintsev P., van der Plicht J., 2013. Mammal fauna during the Late Pleistocene and Holocene in the far northeast of Europe // Boreas. V. 42. P. 779–797.
Pruvost M., Bellone R., Benecke N., Sandoval-Castellanos E., Cieslak M. et al., 2011. Genotypes of predomestic horses match phenotypes painted in Paleolithic works of cave art // Proceedings of the National academy of sciences. V. 108. P. 18 626–18 630.
Rabanus-Wallace M.T., Wooller M.J., Zazula G.D., Shute E., Jahren A.H. et al., 2017. Megafaunal isotopes reveal role of increased moisture on rangeland during late Pleistocene extinctions // Nature Ecology and Evolution. № 1. Article 0125.
Sommer R.S., Benecke N., Lõugas L., Nelle O., Schmölcke U., 2011. Holocene survival of the wild horse in Europe: a matter of open landscape? // Journal of Quaternary Science. V. 26. P. 805–812.
Stampfli H.R., 1963. Wisent, Bison bonasus (Linne) 1758, Ur Bos primigenius Bojanus, 1827, und Hausrind. Bos taurus (Linne), 1758. In: Boessneck J., Jéquier J.P. et Stampfli H.R. (eds): Seeberg, Burgäschisee-Sud. Acta Bernensia, 2, fasc. P. 117–196.
Stuart A.J., Kosintsev P.A., Higham T.F.G., Lister A.M., 2004. Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth // Nature. V. 431. P. 684–689.
Tarasov P.E., Bezrukova E.V., Krivonogov S.K., 2009. Late Glacial and Holocene changes in vegetation cover and climate in southern Siberia derived from a 15 kyr long pollen record from Lake Kotokel // Climate of the Past. V. 5. P. 285–295.
Väliranta M., Kultti S., Seppä H., 2006. Vegetation dynamics during the Younger Dryas-Holocene transition in the extreme northern taiga zone, north-eastern European Russia // Boreas. V. 35. P. 201–212.
Vartanyan S.L., Arslanov K.A., Karhu J.A., Possnert G., Sulerzhitsky L.D., 2008. Collection of radiocarbon dates on the mammoths (Mammuthus primigenius) and other genera of Wrangel Island, northeast Siberia, Russia // Quaternary Research. V. 70. № 1. P. 51–59.
Vislobokova I.A., 2012. Giant deer: origin, evolution, role in the biosphere // Paleontological Journal. V. 46. № 7. P. 643–775.
Vuure C. van, 2005. Retracing the aurochs: history, morphology and ecology of an extinct wild ox. Sofia-Moscow. 431 p.
Vybornov A., Kulkova M., Kosintsev P., Platonov V., Phillipsen B., Nesterov E.M., 2018. Diet and chronology of Neolithic-Eneolithic cultures (from 6500 to 4700 cal BC) in the Low Volga basin // Radiocarbon. V. 60. № 5. P. 1597–1610.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал