

Зоологический журнал, 2022, T. 101, № 12, стр. 1394-1415
Балтийская популяция белощекой казарки (Branta leucopsis (Bechstein 1803)) в бореальной зоне Северо-Запада России
С. А. Коузов a, *, Э. М. Зайнагутдинова a, А. В. Кравчук a
a Санкт-Петербургский государственный университет
199034 С.-Петербург, Россия
* E-mail: skouzov@mail.ru
Поступила в редакцию 20.01.2022
После доработки 18.02.2022
Принята к публикации 12.03.2022
- EDN: OOSAKE
- DOI: 10.31857/S0044513422120066
Аннотация
Белощекая казарка в середине прошлого столетия испытывала значительную депрессию численности. Усиление мер по охране вида на западноевропейских зимовках и потепление климата в Арктике вызвали в последние десятилетия 20-го века рост численности вида и расширение ареала в южном направлении от арктических островов в материковые приморские тундры. Изолированные очаги размножения белощекой казарки образовались в местах миграционных скоплений на побережьях Балтийского моря и в местах зимовки на побережьях Северного моря. Начиная с последнего десятилетия прошлого века, вид стал расселяться в бореальной зоне российского Северо-Запада. В статье на основании собственных данных авторов и анализа научной литературы подробно рассмотрены процесс появления вида на территории Ленинградской области, республики Карелия и Мурманской области, а также изменение его статуса от редкого залетного до мигрирующего и обычного, но локально распространенного гнездящегося вида. Рассматриваются особенности фенологии вида, ландшафтного и биотопического распределения и биологии размножения. Экспоненциальный рост численности вида происходит синхронно как в Арктике, так и в новых местах гнездования в бореальной и умеренной климатических зонах Европы. Предположительно на фоне воздействия изменения климата и улучшения охраны вида имеет место и активный микроэволюционный процесс, что позволяет рассматривать белощекую казарку как интересный модельный вид для различных популяционных исследований.
В последние десятилетия существенно активизировался процесс изменения природной среды. Это связано как с циклическими изменениями климата, так и с последствиями деятельности человека. Среди разных типов биотопов одними из наиболее восприимчивых к изменениям являются побережья крупных водоемов бореальной, субарктической и арктической зон. В связи с этим особый интерес вызывает изучение адаптаций водоплавающих птиц к изменениям их жизненной среды. К таким адаптациям относятся, в частности, изменения ареалов и переселения птиц в совершенно новые для них типы гнездовых ландшафтов.
Белощекая казарка на протяжении большей части 20-го века гнездилась исключительно на арктических островах: на северо-западном побережье Гренландии, на Шпицбергене, Лофотенских о-вах, Вайгаче и на южном острове архипелага Новая Земля (Птушенко, 1952; Alsos et al., 1998; Cramp, Simmons, 1977). При этом чаще всего птицы гнездились на береговых скальных уступах (Мензбир, 1895; Птушенко, 1952).
В середине прошлого века наблюдалась очень сильная депрессия численности этого вида – в начале 50-х годов численность российской популяции белощекой казарки оценивалась всего в 10 000 особей (Boyd, 1961). Затем в течение 80-х годов прошлого века произошло многократное расширение гнездовой области вида в российской части Арктики, когда поселения белощекой казарки появились в полосе континентальных морских побережий на Югорском п-ове (Минеев, 1984), в Малоземельских тундрах (Минеев, Минеев, 2004), на Тиманском берегу (Сыроечковский, 1995), п-ове Канин (Filchagov, Leonovich, 1992), а также на о-ве Колгуев (Гаврило, 1991; Пономарева, 1991, 1992) и архипелаге Земля Франца-Иосифа (Сыроечковский, 1995). В последние десятилетия отмечены находки гнездящихся пар на Таймыре (Головнюк и др., 2015; Харитонов, 2018), а также на баренцевоморском побережье Кольского п-ова (Мельников, 2005), где теперь размножается до 100 пар казарок (Мельников, Плотников, 2017). При этом происходил экспоненциальный рост численности вида (Сыроечковский, 1995; Filchagov, Leonovich, 1992; Fox, Madsen, 2017; Ganter et al., 1999), характерный не только для популяции в целом, но и для динамики большинства новых колоний, взятых по отдельности (Мельников, Плотников, 2017; Минеев, Минеев, 2004; Glazov et al., 2021).
С начала 70-х годов прошлого века отмечается интенсивный рост численности зимующих и мигрирующих белощеких казарок в западном секторе Балтики, небольшие группы птиц стали задерживаться на побережьях Балтийского моря в летний период (Feige et al., 2008). Первый случай гнездования в регионе отмечен на небольшом островке у о-ва Готланд в 1971 г. (Larsson et al., 1988). В дальнейшем белощекая казарка появилась на гнездовании на морских островах западной Эстонии и южной Финляндии (Leito, 1996), Датского архипелага (Mortensen, Hansen, 1999; Olsen, 1992). В начале 1980-х годов гнездовые колонии казарок были обнаружены в Голландии (Meinger, van Swelm, 1994; Ouweneel, 2001; Voslamber et al., 2007), а в конце 1980-х годов – на морском побережье Германии (Koop, 1998; Kruckenberg, Hasse, 2004). Численность колоний вида быстро росла, и в начале XXI века гнездящаяся популяция белощеких казарок в Западной Европе насчитывала 42 000–55 000 птиц (Voslamber et al., 2007).
Несмотря на ряд опубликованных статей по гнездованию и миграциям белощекой казарки в русской части Финского залива и на других территориях Северо-Запада России, мы полагаем, что этот вопрос освещен в литературе крайне фрагментарно. Цель данного сообщения – проследить процесс расселения вида на территории бореальной зоны Северо-Запада России, выявить основные особенности миграций, распределения и биологии размножающихся птиц.
МАТЕРИАЛЫ И МЕТОДИКА
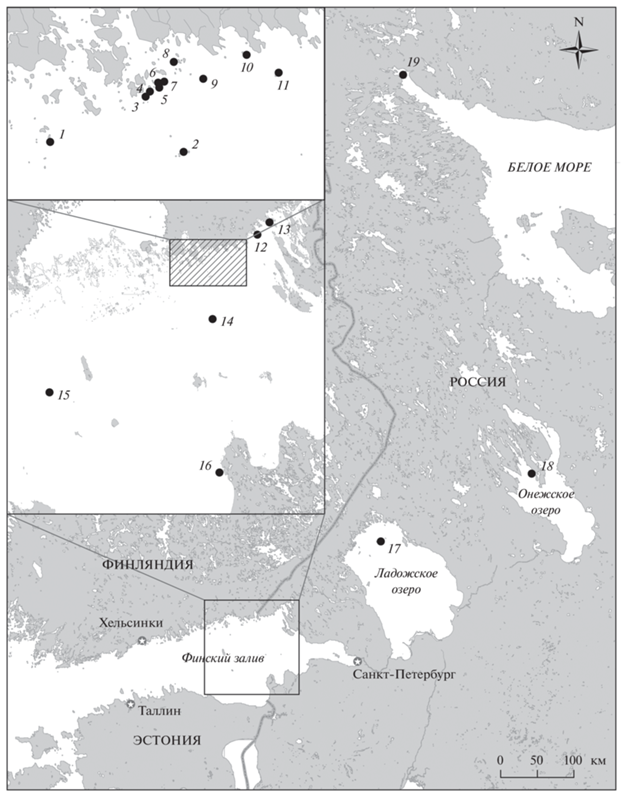
Данные, представленные в статье, получены в результате многолетних исследований морских птиц восточной части Финского залива, проводившихся авторами и другими сотрудниками Санкт-Петербургского университета, Зоологического института РАН и Балтийского Фонда Природы в период с 1987 по 2021 г. Исследовательскими экспедициями были охвачены Сойкинский и Кургальский полуострова, архипелаги Сескар, Виргины, Долгий Риф, Долгий Камень, Большой Фискар, острова Мощный, Малый, Вигрунд, Гогланд, Родшер, Малый и Большой Тютерс, Соммерс, Нерва, Рябинник, Малый Фискар, Гусиный, Стоглаз, банки Хитоматала, Кургальский риф, Тисколовский риф, Вигрунд, Вестгрунд и Виккала (рис. 1).
Рис. 1.
Карта района исследований с указанием мест гнездования белощеких казарок в регионе (черные кружки): 1 – о-в Долгий Риф, 2 – арх. Большой Фискар, 3 – о-в Западный Гребень, 4 – о-в Восточный Гребень, 5 – о-в Каменная Земля, 6 – о-в Галочий, 7 – Малая Отмель, 8 – арх. Толскери, 9 – о-в Рябинник, 10 – о-в Туман, 11 – о-в Малый Фискар, 12 – о-в Гусиный, 13 – о-в Стоглаз, 14 – о-в Нерва, 15 – о-в Родшер, 16 – о-в Реймосар, 17 – Валаамский арх., 18 – о-в Дедова Плешь в Кижских шхерах, 19 – луда Центральная Северного архипелага Кандалакшского залива Белого моря.
Следует отметить, что до конца 80-х годов прошлого века на всех этих территориях существовал строгий пограничный режим, препятствовавший проведению орнитологических исследований. Благодаря этому режиму, местные экосистемы не подвергались негативному антропогенному воздействию (отсутствовали фактор беспокойства и прямое преследование), т.е. условия гнездования птиц были близки к заповедным.
На Кургальском п-ове исследования проводились в 1987–1999 и 2005–2021 гг. В летнее время здесь велись подробные учеты гнездящихся птиц и выводков на прибрежных островах и модельных участках побережья (обычно не менее трех учетов за сезон). В периоды весенних и осенних миграций проводились наблюдения за пролетом: в 1994– 1996 и 2007–2008 гг.– стационарно в апреле–мае и в сентябре–октябре, в остальные годы – на кратких 3–5-дневных выездах 3–4 раза в месяц.
Первые сведения о птицах островов Финского залива получены в результате кратковременных экспедиций БИНИИ в 1991–1992 и 1994–1995 гг. (Бузун, 1997; Носков и др., 1993; Иовченко и др., 2002). В 2005–2008 гг. стационарные обследования гнездящихся и мигрирующих птиц проводились Рычковой (2010) на о-ве Сескар и на о-ве Большой Фискар (Kouzov et al., 2018).
На остальных участках акватории Финского залива исследования продолжались в рамках судовых экспедиций с кратковременными высадками на острова. В 2005–2006 гг. эти исследования проводились экспедициями БИНИИ. В 2010–2014 гг. судовые учеты гнездящихся птиц осуществлялись двумя экспедиционными отрядами, организованными Балтийским Фондом Природы и СПбГУ. В эти годы обычно проводилось по 3 экспедиционных учета. Каждый такой учет продолжался в течение 6–12 дней в период с конца мая до конца июля, при этом учетом были охвачены все островные районы восточной части Финского залива. В 2015 г. в период с 19 мая по 25 июня осуществлено 3 экспедиции длительностью от 3 до 14 дней, охватывавших острова у северного побережья Финского залива, о. Большой Тютерс и арх. Сескар. В 2016 г. в период с 28 мая по 31 июня осуществлено 2 экспедиции на острова Мощный, Вигрунд, Малый Тютерс, архипелаг Виргины, Родшер. В 2017–2021 гг. судовые учеты осуществлялись по аналогичной схеме. В репродуктивный сезон совершалось 2 повторных судовых учета длительностью 10–12 дней. Один из них проводился в конце мая–начале июня, второй – в конце июня–начале июля.
Во время высадок на острова проводили учеты гнезд, выводков и не размножающихся птиц. На небольших островах площадью от 1 до 12 га осуществляли тотальный учет при движении плотным зигзагом, на крупных островах (Большой и Малый Тютерс, Мощный, Малый) подробно обследовали только береговую полосу. Найденные гнезда с кладками описывали по стандартным методикам, производили замеры яиц, сроки откладки яиц определяли по результатам водного теста.
На открытой акватории и около островов миграции изучали во время 4–8-дневных судовых учетов в сентябре-октябре в 2010 и в 2012–2016 гг. Всего проведено 17 таких учетов. Весенние миграции над открытым морем изучали на маршрутах во второй половине мая одновременно с учетами гнездящихся птиц на островах.
В работе использованы 25-кратный бинокль, 100-кратная подзорная труба, фотоаппараты Nikon D90 и Nikon D7200 с объективами Nikkor 300:4 AF-S и Sigma 170-500:5.6-6.4.
Для сравнения величин кладок и яиц казарок, гнездящихся в восточной части Финского залива (поздние и ранние кладки), использовался t-критерий Стьюдента, вычисленный в программе Statsoft Statistica (Statsoft Statistica, 2011). Для сравнения величин кладок, обследованных нами, и кладок казарок в Арктике (литературные данные) использовался t-критерий, вычисленный не на основе выборок, а по формуле (Лакин, 1990) на основе средних и стандартных отклонений (поскольку более подробные данные по Арктике нам недоступны).
РЕЗУЛЬТАТЫ
История расселения и динамика численности вида в регионе
На протяжении всего периода орнитологических исследований вплоть до конца 80-х годов прошлого века белощекая казарка была крайне редким пролетным видом на территории Ленинградской обл. и Карелии (Мальчевский, Пукинский, 1983; Нейфельдт, 1970; Putkonen, 1940). В Архангельской обл. в середине 20-го века белощекая казарка на пролете также почти не наблюдалась (Белопольский, 1956). Однако уже с 1960-х годов интенсивные осенние миграции вида стали отмечать на различных участках у побережья Белого моря: в горле Белого моря, на восточном берегу Онежского п-ова (Кищинский, 1979) и на юго-западном берегу Онежского залива (Кумари, 1963).
Значительное увеличение численности белощекой казарки на пролете в восточной части Финского залива началось в конце 80-х годов прошлого века (Бубырева и др., 1993; Коузов, 1995). В следующем десятилетии она стала здесь многочисленным мигрантом (Бузун, 1998; Коузов, 1995; Kontiokorpi, Rusanen, 2014; Васильева, 2001). В эти годы массовые весенние миграционные стоянки белощекой казарки стали образовываться в окрестностях г. Олонец в Южной Карелии (Лапшин и др., 2016; Zimin et al., 2002) и в устье р. Северная Двина на Белом море (Андреев, 2005). В последние годы белощекая казарка стала одним из самых массовых мигрантов в восточной части Финского залива (Коузов, 2009, 2010, 2011; Коузов, Кравчук, 2010; Коузов, Лосева, 2014, 2014а), в Южной Карелии (Артемьев и др., 2009, 2011) и на Онежском п-ове Белого моря (Волков и др., 2015).
22 июня 1995 г. гнездо белощекой казарки с кладкой из 6 яиц было найдено на о-ве Долгий Риф (Гагинская и др., 1997) у северного побережья Финского залива на границе с Финляндией. К 2006 г. численность размножающейся группировки этого вида у северного побережья Финского залива возросла до 31 пары, гнезда найдены на островах Малый Фискар, Долгий Риф (А.Л. Рычкова, личное сообщение) и архипелаге Большой Фискар (табл. 1). Здесь держалось и несколько десятков неразмножающихся особей. В этом же году отмечен случай размножения одной пары белощеких казарок на о-ве Реймосар у западного побережья Кургальского п-ова на южном берегу Финского залива (Коузов, Кравчук, 2008).
Таблица 1.
Динамика находок гнезд белощекой казарки на островах восточной части Финского залива в 1995–2021 гг.
| Остров | 1995 | 2005 | 2006 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | Всего | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Родшер | – | – | – | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Долгий Риф | 1 | – | 22 | 1 | 5 | 4 | 1 | 5 | 8 | 9 | 7 | 9 | 8 | 6 | 5 | 91 | |
| Западный Гребень | 0 | 0 | – | – | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 4 | |
| Восточный Гребень | 0 | 0 | – | – | 0 | 0 | 0 | 5 | 6 | 4 | 2 | 3 | 1 | 2 | 1 | 24 | |
| Каменная Земля | 0 | 0 | – | – | 0 | 0 | 0 | 2 | 3 | 2 | 0 | 0 | 0 | 1 | 0 | 8 | |
| Галочий | 0 | 0 | – | – | 3 | – | 1 | 3 | 5 | 3 | 0 | 0 | 0 | 1 | 0 | 16 | |
| Малая Отмель | 0 | 0 | – | – | 6 | – | 3 | 10 | 14 | 8 | 1 | 3 | 1 | 1 | 2 | 49 | |
| Рябинник | 0 | 0 | – | – | 4 | 1 | 2 | 6 | 10 | 11 | 8 | 10 | 8 | 7 | 4 | 71 | |
| арх. Большой Фискар | Фискар | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 6 |
| Кивимаа | 0 | 0 | 2 | 0 | 0 | 1 | 1 | 1 | 1 | 2 | 4 | 3 | 4 | 1 | 0 | 20 | |
| Маннонен | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 2 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 10 | |
| Большой Западный | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 0 | 0 | 0 | 8 | |
| Туман | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 3 | 2 | 1 | 0 | 2 | 1 | 13 | |
| Малый Фискар | 0 | 3 | 5 | 2 | 6 | 2 | 3 | 4 | 12 | 10 | 10 | 9 | 6 | 8 | 7 | 87 | |
| Нерва | 0 | – | – | – | 0 | 2 | 2 | 2 | 3 | 2 | – | 1 | 0 | 1 | 1 | 14 | |
| Стоглаз | – | – | – | – | 0 | 0 | – | 0 | 3 | 10 | 9 | 12 | 7 | 8 | 6 | 55 | |
| Гусиный | – | – | – | – | – | – | – | – | – | 0 | 1 | 1 | 1 | 2 | 0 | 5 | |
| арх. Толскери | – | – | – | – | – | – | – | – | – | – | – | – | – | – | 1 | 1 | |
| Реймосар | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Всего | 1 | 3 | 32 | 5 | 25 | 13 | 15 | 42 | 76 | 67 | 46 | 55 | 36 | 40 | 28 | 484 | |
К 2010–2012 гг. численность белощеких казарок, гнездящихся в российской части Финского залива, варьировала от 5 до 25 пар. В 2010 г. гнездование этого вида отмечено на о-ве Родшер в центральной части Финского залива в 16 км к западу от о-ва Гогланд, в 2011 г. гнездящиеся пары белощеких казарок впервые найдены на островах Рябинник, Малая Отмель и Галочий у северного побережья Финского залива, в 2012 г. – размножение 2 пар наблюдалось на о-ве Нерва в открытой части Финского залива (табл. 1).
В 2014 и 2015 гг. зарегистрирован взрывной рост численности гнездовой группировки белощеких казарок в восточной части Финского залива – до 42 и 76 найденных гнезд за сезон, соответственно (табл. 1). В эти годы наблюдалось дальнейшее расширение зоны гнездования в северной части Финского залива. В 2014 г. белощекие казарки стали размножаться на островах Восточный Гребень, Каменная земля, в 2015 г. – на островах Западный Гребень, Туман и Стоглаз (Храбрый, Байбекова, 2016; наши данные).
В 2016–2021 гг. отмечена тенденция к снижению численности вида, однако в этот период также зарегистрированы новые места гнездования. Начиная с 2017 г. 1–2 пары белощеких казарок гнездились на о-ве Гусиный в Выборгском заливе, в 2021 г. гнездо белощекой казарки найдено на небольшом островке на архипелаге Толскери.
Восточнее Финского залива гнездование двух пар белощекой казарки впервые отмечено на островах Валаамского архипелага на Ладожском оз. в 2010 г., в последующие годы число гнездящихся птиц здесь постоянно росло – в 2013 г. уже гнездилось 5 пар (Агафонова и др., 2016). В 2015 г. первый случай гнездования одной пары зарегистрирован на Онежском оз. на о-ве Дедова Плешь в юго-западной части Кижских Шхер (Хохлова, Артемьев, 2015). В 2017 г. одна пара белощеких казарок загнездилась на луде Центральная Северного архипелага в шхерах Кандалакшского залива Белого моря (Шутова, Кожин, 2018).
Весенние миграции и предгнездовой период
Появление первых белощеких казарок на Финском заливе в годы с ранним развитием весенних событий отмечается в середине апреля (Коузов, 2010), в годы с поздней весной – в последних числах апреля (Рымкевич и др., 2012). До середины мая наблюдается пролет отдельных стай, часть из которых делает остановки на низкотравных луговинах островов и побережий. Такие стоянки известны на западном побережье Кургальского п-ова в угодье Кирьямо – до 1250 птиц (Коузов, 2010) и на о-ве Реймосар – до 350–400 птиц (Коузов, 2015), на островах Кургальского рифа – до 2000 птиц (Коузов, 2015), о-ве Мощный – до 500 птиц (Коузов, Лосева, 2014), на архипелаге Большой Фискар – до 250 птиц (Коузов, Лосева, 2014), Большом Тютерсе – 80–100 особей и Малом Тютерсе – 150–200 птиц. Небольшие группы отдыхающих птиц отмечаются на банках Лужской губы (Коузов, Лосева, 2014), на архипелаге Сескар, островах Малый Фискар и Рябинник, на мысах Конек, Портовый и Крестовый на северном побережье Финского залива.
Однако бо́льшая часть птиц в это время наблюдается в восточном Приладожье, где с третьей декады апреля начинают образовываться массовые долговременные стоянки белощеких казарок на полях около города Олонец и пос. Шуя (Артемьев и др., 2009, 2010, 2011, 2013).
Миграции белощеких казарок продолжаются до конца мая. Валовый пролет на Финском заливе происходит в течение нескольких дней в четвертой–пятой пятидневках мая. В это время за сутки через наблюдательный пункт может пролететь от 10 000–20 000 до 80 000 и в некоторых случаях даже до 114 000 птиц (Коузов, 2010; Коузов, Кравчук, 2010; Kontiokorpi, Rusanen, 2014). Практически одновременно происходит массовый отлет птиц со стоянок в восточном Приладожье (Артемьев и др., 2009, 2010, 2011, 2013; Лапшин и др., 2016) и валовая миграция на Белом море (Андреев, 2005; Волков, 2013; Волков и др., 2015).
Птицы балтийской популяции в местах гнездования на Финском заливе появляются существенно раньше начала валового пролета арктических популяций. Так, в 2015 г. небольшие группы из 10–15 птиц у островов Стоглаз, Малый Фискар, Туман и Рябинник наблюдались уже 24 апреля. В группах были хорошо заметны пары, которые активно обследовали наиболее возвышенные центральные части островов в поисках мест, подходящих для устройства гнезд. Периодически пары и группы из 2–3 пар делали круговые облеты над акваторией вокруг островов.
Гнездовой период
Сроки размножения. В 2005–2006 гг. откладка яиц в гнездах белощеких казарок в восточной части Финского залива начиналась с 4-й декады мая (Kouzov et al., 2019). По данным 2010–2021 гг., в самые ранние кладки могли начинаться уже в 1–2-й декаде мая (табл. 2). Период начала откладки яиц в эти годы продолжался и в течение июня. В 6 сезонах из 11 он продолжался до 5-й пятидневки июня, в остальные годы мог заканчиваться в течение первой половины месяца. Основная масса кладок (73%, n = 398), была начата в 3–6-й пятидневках мая, с пиком в 3–4-й пятидневках.
Таблица 2.
Сроки начала откладки яиц в гнездах белощекой казарки (Branta leucopsis) в восточной части Финского залива в 2010–2021 гг.
| Год | Май (пятидневки) | Июнь (пятидневки) | Всего | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | ||
| 2010 | 2 | 1 | 1 | 4 | |||||||||
| 2011 | 2 | 5 | 4 | 2 | 1 | 2 | 16 | ||||||
| 2012 | 1 | 2 | 3 | 1 | 1 | 8 | |||||||
| 2013 | 1 | 2 | 4 | 2 | 1 | 1 | 11 | ||||||
| 2014 | 3 | 6 | 5 | 3 | 4 | 1 | 1 | 2 | 3 | 28 | |||
| 2015 | 6 | 15 | 14 | 9 | 11 | 4 | 1 | 1 | 6 | 3 | 70 | ||
| 2016 | 4 | 14 | 11 | 10 | 12 | 2 | 2 | 4 | 4 | 63 | |||
| 2017 | 3 | 5 | 5 | 4 | 19 | 2 | 1 | 39 | |||||
| 2018 | 2 | 12 | 14 | 9 | 6 | 3 | 1 | 5 | 1 | 2 | 55 | ||
| 2019 | 4 | 9 | 8 | 7 | 4 | 1 | 2 | 1 | 36 | ||||
| 2020 | 2 | 8 | 12 | 10 | 5 | 1 | 1 | 1 | 40 | ||||
| 2021 | 4 | 11 | 7 | 3 | 2 | 1 | 27 | ||||||
| Всего | 5 | 38 | 92 | 86 | 72 | 42 | 17 | 5 | 10 | 15 | 15 | 398 | |
Откладка яиц в гнездах белощеких казарок на Валаамском архипелаге Ладожского оз. происходит во 2-й половине мая (Агафонова и др., 2016). Кладка, найденная в северной части Онежского озера в 2015 г., была начата 10 июня (Хохлова, Артемьев, 2015). В гнезде, найденном в Кандалакшском заливе Белого моря в 2017 г., судя по данным авторов, кладка была начата приблизительно 20 июня (Шутова, Кожин, 2018). Столь поздние сроки размножения авторы находки связывают с высокой вероятностью повторного размножения после потери первой кладки (там же).
Пространственное и ландшафтное распространение размножающихся птиц. Все известные к настоящему моменту случаи гнездования балтийской популяции белощекой казарки в бореальной зоне Северо-Запада России приурочены к четырем наиболее крупным водоемам региона. Это – восточная часть Финского залива, Ладожское и Онежское озера, а также Кандалакшский залив Белого моря (Агафонова и др., 2016; Хохлова, Артемьев, 2015; Kouzov et al., 2018). Подавляющее большинство гнезд вида найдено в северных частях этих водоемов на границе Балтийского кристаллического щита. Это районы, где доминирует так называемый сельговый ландшафт, – сглаженные ледником гранитные скалы. Для них характерны сильная изрезанность береговой линии и наличие многочисленных прибрежных островов.
В этих районах белощекие казарки гнездились исключительно на скалистых островах, площадью от 0.7 до 9.4 га, высотой от 3 до 12 м, и удаленных от берега на расстояние от 0.4 до 28.3 км (табл. 3). Из-за резкого свала глубин мелководная зона вокруг этих островов практически отсутствует. Преобладающие глубины на Финском заливе около этих островов варьировали от 5–10 до 10–40 м (табл. 3).
Таблица 3.
Характеристика островов в восточной части Финского залива, где отмечено размножение белощеких казарок в 1995–2021
| Остров | Всего случаев размножения | Ландшафтно-биотопическая характеристика островов | Площадь, га | Высота, м | Расстояние от берега или от крупного лесного острова, км | Доминирующие глубины около островов | |
|---|---|---|---|---|---|---|---|
| Родшер | 1 | Моренный валунно-галечниковый остров | 2.1 | 3.5 | 17.7 | 30–40 | |
| Долгий Риф | 91 | Сельговая скала с низкотравными луговинами и отдельными кустами в центре острова | 7.7 | 6.0 | 4.8 | 5–15 | |
| Западный Гребень | 4 | То же | 1.2 | 3.5 | 0.8 | 5–10 | |
| Восточный Гребень | 24 | То же | 2.3 | 3.5 | 1 | 5–10 | |
| Каменная Земля | 8 | То же | 0.9 | 3.5 | 1.6 | 5–10 | |
| Галочий | 16 | То же | 1.6 | 4.0 | 1.3 | 5–10 | |
| Малая Отмель | 49 | То же | 3.7 | 5.5 | 1.8 | 5–10 | |
| Рябинник | 71 | Высокая сельговая скала с низкотравными луговинами и отдельными кустами в центре острова | 4.0 | 4.5 | 2.62 | 5–20 | |
| арх. Большой Фискар | Фискар | 6 | Сельговая скала с небольшими низкотравными и среднетравными дерновинами в центре острова | 1.4 | 5.5 | 10.1 | 10–40 |
| Кивимаа | 20 | То же | 1.6 | 5.5 | 10.1 | 10–40 | |
| Маннонен | 10 | Высокая сельговая скала с низкотравными и среднетравными луговинами и отдельными деревьями и кустами в центре острова | 3.6 | 12.0 | 10.1 | 10–40 | |
| Западный | 8 | Сельговая скала с редким травяным покровом в скальных трещинах | 2.7 | 6.0 | 10.1 | 10–40 | |
| Туман | 13 | Сельговая скала с редким травяным покровом в скальных трещинах и древесно-кустарниковой растительностью в центре | 0.7 | 6.5 | 0.7 | 5–15 | |
| Малый Фискар | 87 | Сельговая скала с низкотравными и среднетравными луговинами, а также с отдельными кустами и деревьями в центре острова | 5.1 | 6.5 | 2.43 | 10–35 | |
| Нерва | 14 | Сельговая скала с незначительным травяным покровом в скальных трещинах | 9.4 | 7.5 | 28.3 | 10–35 | |
| Стоглаз | 55 | Сельговая скала с незначительным травяным покровом в скальных трещинах и древесно-кустарниковой растительностью в центре | 4.9 | 5.2 | 0.4 | 5–10 | |
| Гусиный | 5 | Высокая сельговая скала с участками низкотравных и среднетравных луговин, а также с валунно-галечниковыми отложениями в понижении в центре острова | 0.8 | 4.1 | 1.9 | 8–10 | |
| Безымянный островок на арх. Толскери | 1 | Сельговая скала с отдельными валунами, поросшая злаково-ландышевым сосново-мелколиственным лесом | 0.8 | 2.3 | 1.5 | 4–5 | |
| Реймосар | 1 | Моренный валунно-песчаный остров с луговинами различного типа и тростниковыми зарослями | 12.5 | 1.5 | 1.34 | 1–5 | |
Наибольшее количество случаев размножения белощеких казарок в восточной части Финского залива отмечено на островах Долгий Риф и Малый Фискар, архипелаге Большой Фискар, а также на островах Рябинник и Стоглаз, где суммарно найдено 71.9% всех гнезд (n = 484) (табл. 1 и 3).
За последние 25 лет на Финском заливе известно только два случая гнездования белощекой казарки вне вышеописанной ландшафтной зоны, которые можно трактовать как исключение из правила. Так, в 2006 г. одна пара птиц размножалась на моренном валунно-песчаном о-ве Реймосар у южного побережья Финского залива, а в 2010 г. одна пара птиц гнездилась на моренном галечниково-валунном о-ве Родшер в центре Финского залива в 17.7 км к юго-западу от о-ва Гогланд. В последующие годы птицы здесь достоверно не размножались.
На Валаамском архипелаге Ладожского оз. и Кижском архипелаге Онежского оз. все гнезда белощекой казарки также были найдены на небольших высоких скалистых островах (Агафонова и др., 2016; Хохлова, Артемьев, 2015). При этом на Валаамском архипелаге птицы занимали исключительно луды без древесной растительности (Агафонова и др., 2016), а на о-ве Дедова Плешь на Онежском оз., где в 2015 г. было отмечено размножение белощекой казарки, росли несколько сосен (Хохлова, Артемьев, 2015). В Кандалакшском заливе Белого моря в 2017 г. птицы также выбрали для размножения безлесный островок с тундроподобной растительностью и отдельными кустами можжевельника (Шутова, Кожин, 2018).
Как видно из табл. 1 и 3, до 2010 г. в восточной части Финского залива белощекие казарки гнездились преимущественно на островах, удаленных от берега на расстояние от 2.4 до 10.1 км, – 97.6% гнезд, n = 41. В последующие годы рост численности вида шел в первую очередь за счет птиц, заселявших самые близкие к побережью острова не далее 2 км – в период 2011–2021 гг. здесь гнездилось 39.5% всех пар (n = 443), а в сезоны с максимальной численностью гнездящихся пар в 2014–2016 гг. этот показатель возрастал соответственно до 47.6 (n = 42), 48.7 (n = 76) и 46.3% (n = 67) (рис. 2). Отмечена сильная значимая корреляция между долей птиц, гнездящихся на близких к берегу островах (от общего числа найденных в течение сезона гнезд), и общей численностью гнездовой группировки (r = 0.79, p < 0.05).
Рис. 2.
Динамика численности гнездящейся группировки белощеких казарок в северной части Финского залива и динамика численности казарок, гнездящихся на островах ближе 2 км в 1995–2021 гг.
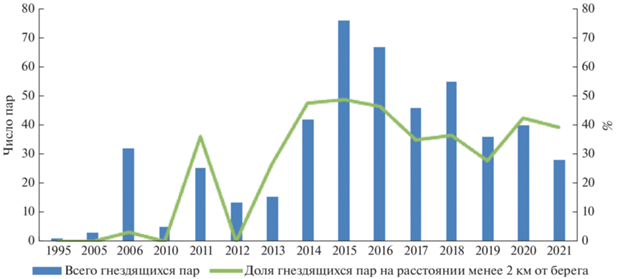
О том, что казарки в годы подъемов численности предпочитают гнездиться на самых близких к берегу островах, свидетельствует и более высокая плотность их гнездования здесь по сравнению с более удаленными островами. Так, в 2015, 2016 и 2020 гг. выявлена значимая отрицательная корреляция между удаленностью островов от берега и плотностью гнездования белощеких казарок на этих островах (r = –0.59, –0.60 и –0.53 соответственно, p < 0.05).
Микробиотопическое распределение гнезд. Почти все подробно описанные в 2010–2021 гг. гнезда белощеких казарок (n = 411) на островах Финского залива находились во внутренних частях островов, далеко от зоны заплеска. В зависимости от величины острова расстояние от гнезда до уреза воды варьировало от 12 до 39 м, в среднем составило 22.62 ± 7.37 м. Высота их расположения над водой варьировала от 1.7 до 11 м, в среднем 3.43 ± 1.94 м.
За все годы исследований в нижнем ярусе у подножия скал найдены всего 11 гнезд (2.7% всех гнезд, n = 411). Подавляющее большинство гнезд располагалось в среднем ярусе скал – 65.0% всех находок (n = 411), на верхнем ярусе скал – 32.4% всех находок (n = 411).
Все гнезда в верхнем ярусе скал (133 гнезда) находились на выровненных плато. Среди гнезд, найденных в среднем ярусе скал (267 гнезд), большинство (79.8%) располагалось на широких уступах или в широких долинах между грядами и только 12.7% гнезд здесь были устроены в глубоких узких скальных расщелинах. Все гнезда, обнаруженные в нижнем ярусе скал (11 гнезд), находились на широких скальных уступах. При этом 6 из них располагались на микроподнятиях рельефа – дерновинах, сформированных на завалах валунов.
Пары белощеких казарок занимали достаточно широкий спектр микробиотопов (9 разновидностей) от полностью лишенной растительности скальной поверхности до кустарников и лесных сосново-мелколиственных участков (табл. 4). Чаще всего мы находили гнезда на высокотравных луговинах (29.7%, n = 411) и под прикрытием отдельно стоящих кустарников или низких деревьев (21.7%, n = 411). В 10.2% случаев (n = 411) гнезда располагались на сосново-мелколиственных лесных участках. При этом в начальные периоды колонизации казарки выбирали для размножения исключительно безлесные острова (Kouzov et al., 2018; Zaynagutdinova et al., 2019), а освоение лесных биотопов началось только с 2015 г. при вселении птиц на острова Стоглаз и Толскери (табл. 1).
Таблица 4.
Микробиотопическое распределение гнезд белощекой казарки на островах восточной части Финского залива в 2010–2021 г. по данным о подробно описанных гнездах (n = 411)
| Ярус скал | Тип микрорельефа | Биотоп | Число гнезд |
|---|---|---|---|
| Верхний ярус скал | Выровненное плато | Открыто в микропонижении на голой скале | 21 |
| Открыто на тонком наносе песка на скале с редкой низкотравной растительностью | 29 | ||
| Низкотравная луговина на скальной подложке | 34 | ||
| Под отдельными кустами ивы, рябины, альпийской смородины или под низкой скальной сосной | 26 | ||
| Участки злаково-ландышевого сосново-мелколиственного леса с кустарниками | 23 | ||
| Средний ярус скал и широкие долины между валами | Широкие уступы и долины между валами | Низкотравная луговина на скальной подложке | 41 |
| Среднетравная луговина на скальной подложке | 33 | ||
| Высокотравная луговина в небольшом скальном понижении на плато | 80 | ||
| Под отдельными кустами ивы, рябины, альпийской смородины или под низкой скальной сосной | 40 | ||
| Участки злаково-ландышевого сосново-мелколиственного леса с кустарниками | 19 | ||
| Глубокие узкие скальные расщелины | Высокотравная луговина в глубокой скальной ложбине | 31 | |
| Под отдельными кустами ивы, рябины, альпийской смородины или под низкой скальной сосной | 23 | ||
| Нижний ярус скал | Широкие уступы | Высокотравные луговины и дерновины густых злаков на скоплениях валунов, лежащих на скальной поверхности | 11 |
| Всего | 411 | ||
При этом чем выше был ярус скал, тем чаще гнезда располагались открыто. Так, в верхнем ярусе доля гнезд, помещенных открыто на голых скалах, среди тонких наносов песка с отдельными куртинками низкой травы или на низкотравных луговинах совокупно составила 63.2% (133 гнезда). На среднем ярусе скал открытое расположение гнезд было отмечено только на низкотравных луговинах – 15.4% (267 гнезд). В отличие от верхнего яруса здесь появляются более закрытые гнезда в высокотравных и среднетравных луговинах – соответственно 30.0 и 12.4% случаев. В нижнем ярусе все гнезда (100% случаев, 11 гнезд) располагались в высокотравных луговинах и куртинах.
Более четверти всех найденных гнезд (26.3%, n = 411) располагалось под прикрытием древесно-кустарниковой растительности (на лесных участках или под отдельно стоящими кустами). Но эти биотопы использовались птицами только в верхнем и среднем ярусах скал.
Межвидовые территориальные отношения. Белощекие казарки в восточной части Финского залива гнездились почти исключительно на островах, занятых массовыми колониальными поселениями морских птиц (табл. 5). Всего на этих островах гнездилось 16 видов крупных водоплавающих птиц (Kouzov et al., 2019). Самыми массовыми видами были большой баклан и серебристая чайка. Наиболее широкое распространение по островам было у гаги и серебристой чайки (табл. 5). Распространены по большинству островов были также хохлатая чернеть, большой и средний крохали, клуша. Сизая чайка отмечалась на самых близких к побережью островках. Остальные виды встречались только эпизодически.
Таблица 5.
Численность и распространение массовых колониальных птиц на 17 островах, занятых поселениями белощекой казарки, в северной зоне восточной части Финского залива
| Вид | Количество островов, занятых поселениями вида | Количество гнездящихся пар вида | Гнезда белощекой казарки
по соседству с данным видом |
Минимальное расстояние между гнездами | Среднее расстояние между гнездами | |
|---|---|---|---|---|---|---|
| количество | доля гнезд, % | |||||
| Большой баклан (Phalacrocorax carbo) |
6 | 1140–2240 | 71 | 17.27 | 12 | 15.31 ± 1.85 |
| Лебедь-шипун (Cygnus olor) |
7 | 1–8 | 8 | 1.95 | 15 | 18.43 ± 2.01 |
| Серый гусь (Anser anser) |
8 | 1–9 | 16 | 3.89 | 7 | 11.55 ± 3.02 |
| Хохлатая чернеть (Aythya fuligula) |
10 | 11–37 | 77 | 18.73 | 2.8 | 3.93 ± 1.11 |
| Большой крохаль (Mergus merganser) |
11 | 11–19 | 31 | 7.54 | 2.5 | 3.77 ± 1.03 |
| Средний крохаль (Mergus serrator) |
11 | 9–17 | 40 | 9.73 | 1.8 | 4.94 ± 1.73 |
| Турпан (Melanitta fusca) |
3 | 1–7 | 1 | 0.24 | 11.5 | 11.5 |
| Обыкновенная гага (Somateria mollissima) |
14 | 101–188 | 331 | 80.54 | 1.2 | 2.91 ± 1.32 |
| Серебристая чайка (Larus argentatus) |
13 | 1340–2240 | 399 | 97.08 | 1.5 | 2.74 ± 1.43 |
| Клуша (Larus fuscus) |
11 | 70–150 | 36 | 8.76 | 1.5 | 3.82 ± 2.12 |
| Морская чайка (Larus marinus) |
8 | 6–10 | 6 | 1.46 | 2.5 | 4.45 ± 2.75 |
| Сизая чайка (Larus canus) |
4 | 21–32 | 20 | 4.87 | 4.5 | 6.71 ± 1.68 |
| Речная крачка (Sterna hirundo) |
2 | 32–105 | 6 | 1.46 | 5.5 | 7.33 ± 2.68 |
| Чеграва (Hydroprogne caspia) |
2 | 0–37 | 11 | 2.68 | 12 | 12 |
| Гагарка (Alca torda) |
6 | 44–85 | 2 | 0.49 | 11 | 11.5 |
| Атлантический чистик (Cepphus grylle) |
6 | 2–8 | 1 | 0.24 | 12 | 12 |
Наиболее часто белощекие казарки размножались в поселениях серебристой чайки и гаги (табл. 5), дистанции между гнездами белощекой казарки и гнездами этих видов были существенно меньше, чем в случаях с другими видами. Наибольшие дистанции отмечены между гнездами белощекой казарки и гнездами больших бакланов, лебедей-шипунов, речных крачек, чеграв и чистиковых птиц.
На Валаамском архипелаге Ладожского оз. и в Кижских шхерах все известные гнезда белощеких казарок также были найдены на островах с колониями серебристой, сизой чаек и клуши (Агафонова и др., 2016; Хохлова, Артемьев, 2015).
Среди казарок, гнездящихся в восточной части Финского залива на верхнем ярусе голых и слабозарастающих низкой травянистой растительностью скал (n = 29), выявлено 7 случаев занятия казарками гнезд обыкновенных гаг. В двух случаях насиженные яйца гаги оказались впоследствии за бортиком гнезда, в пяти случаях образовались смешанные кладки из одного–двух яиц гаги и 4–6 яиц белощекой казарки. В двух таких случаях удалось наблюдать успешное одновременное вылупление птенцов обоих видов (рис. 3), вероятно, казарки занимали гнезда гаг в период откладки последними яиц или в самом начале инкубации. В остальных случаях, судя по степени насиженности яиц, изгнание гаг с гнезд происходило на 6–10-й день инкубации. Возможно, конкуренция за место гнездования была связана с дефицитом микроуглублений в скальной поверхности, подходящих для размещения кладок.
Рис. 3.
Вылупление в смешанной кладке белощекой казарки и обыкновенной гаги. О-в Рябинник, 6 июня 2019 г.

На Кургальском п-ове в 2005 и 2008 гг. наблюдались одиночные белощекие казарки, которые в качестве третьей птицы-няньки находились неподалеку от выводков серых гусей (Коузов, Кравчук, 2008). В таких случаях белощекая казарка держалась либо последней непосредственно за замыкающим выводок одним из родителей (чаще самцом), либо сбоку от выводка. Такие казарки отличались несколько гипертрофированным охранным поведением, чаще родителей поднимая тревоги и демонстрируя агрессию при приближении крупных чаек. В 2006 г. такая птица наблюдалась в составе неразмножающегося трио с серыми гусями (Коузов, Кравчук, 2008). Судя по поведению, это была самка. Во время полетов она все время стремилась оказаться между двумя гусями, сразу за самцом. На воде и суше самец серого гуся обычно “дежурил” на камне, в то время как две другие птицы кормились или отдыхали. В 2014 г. в Кижских шхерах Онежского оз. наблюдали белощекую казарку, державшуюся постоянно в паре с серебристой чайкой (Хохлова, Артемьев, 2015). Подобные поведенческие аномалии, на наш взгляд, связаны с дефицитом партнеров, возникающим при расселении птиц на новые территории.
Внутривидовые территориальные отношения. Подавляющее большинство пар (80.3% всех гнезд, n = 486) в восточной части Финского залива гнездилось в 1995–2021 гг. в составе моновидовых колоний (табл. 6).
Таблица 6.
Распределение гнезд белощекой казарки в моновидовых колониях в восточной части Финского залива в 1995–2021 гг.
| Остров | 1995 | 2005 | 2006 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | Всего гнезд в колониях | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Родшер | – | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Долгий Риф | – | 22 | 0 | 5 | 4 | 0 | 5 | 8 | 7 | 6 | 8 | 7 | 6 | 5 | 83 | ||
| Западный Гребень | 0 | 0 | – | – | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | |
| Восточный Гребень | 0 | 0 | – | – | 0 | 0 | 0 | 5 | 6 | 4 | 2 | 3 | 0 | 2 | 0 | 22 | |
| Каменная Земля | 0 | 0 | – | – | 0 | 0 | 0 | 2 | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 7 | |
| Галочий | 0 | 0 | – | – | 3 | – | 0 | 3 | 5 | 3 | 0 | 0 | 0 | 0 | 0 | 14 | |
| Малая Отмель | 0 | 0 | – | – | 6 | – | 0 | 10 | 14 | 7 | 0 | 2 | 0 | 0 | 0 | 39 | |
| Рябинник | 0 | 0 | – | – | 4 | 0 | 0 | 6 | 10 | 11 | 8 | 10 | 8 | 7 | 4 | 68 | |
| арх. Большой Фискар | Фискар | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Кивимаа | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 4 | 0 | 0 | 10 | |
| Маннонен | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Большой Западный | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Туман | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 3 | 2 | 0 | 0 | 0 | 0 | 9 | |
| Малый Фискар | 0 | 3 | 5 | 0 | 6 | 0 | 3 | 4 | 12 | 10 | 10 | 8 | 6 | 8 | 6 | 81 | |
| Нерва | 0 | – | – | – | 0 | 0 | 0 | 0 | 0 | 0 | – | 0 | 0 | 0 | 0 | 0 | |
| Стоглаз | – | – | – | – | 0 | 0 | – | 0 | 3 | 9 | 9 | 10 | 9 | 7 | 6 | 53 | |
| Гусиный | – | – | – | – | – | – | – | – | – | 0 | 0 | 0 | 0 | 2 | 0 | 2 | |
| Толскери | – | – | – | – | – | – | – | – | – | – | – | – | – | – | 0 | 0 | |
| Реймосар | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Всего | 0 | 3 | 27 | 0 | 24 | 4 | 3 | 35 | 67 | 56 | 40 | 44 | 34 | 32 | 21 | 390 | |
Анализ имеющихся данных выявил значимую корреляцию доли колониальных пар белощеких казарок от общей численности гнездовой группировки (r = 0.59, p < 0.05). Кроме того, доля колониальных пар увеличивалась по мере уменьшения расстояния от острова до берега или крупного лесного острова. Корреляция между долей (%) колониальных пар на острове от расстояния (км) от острова до берега составила r = –0.58, p < 0.05. На островах, удаленных от берега на более чем 10 км, гнездились только одиночные пары казарок.
Описание гнезд. Отмечено два типа гнезд. Почти все подробно описанные гнезда (75.9%, n = 29), которые располагались в верхнем ярусе скал в микропонижениях субстрата, были примитивными лунками с незначительном слоем подстилки из мелкого растительного мусора с диаметром лотка 21–25 см. Бортики построек состояли исключительно из обильного пуха. Гнезда в среднем и нижнем ярусах скал, помещавшиеся среди высокотравных и низкотравных луговин и под кустами, являлись крупными постройками из измельченных злаков: диаметр 45–55 см, высота валика над субстратом 5–10 см, диаметр лотка 21–26 см, глубина лотка 5.0–6.5 см.
Величина кладки и размеры яиц. В кладках содержалось от двух до шести яиц, в среднем 4.79 ± ± 1.13 (n = 139). Размеры яиц 68.6–81.3 × 47.4–51.9 мм, в среднем 73.30 ± 3.12 × 49.99 ± 1.57 мм (n = 655). Отмечено существенное значимое уменьшение этих величин в кладках, начатых во вторую половину периода откладки яиц (табл. 7).
Таблица 7.
Средние величины кладок и яиц белощекой казарки, отложенных в первую (2–5-я пятидневки мая) и вторую (1–5-я пятидневки июня) половины периода откладки яиц и значения t-критерия для сравнения этих величин
| Показатель | Май | Июнь | Весь период | t-значение |
|---|---|---|---|---|
| Величина кладки | 5.17 ± 0.81 (n = 110) |
3.34 ± 1.01 (n = 29) |
4.79 ± 1.13 (n = 139) |
10.24 (p < 0.001) |
| Длина яиц | 73.64 ± 3.12 (n = 558) |
71.35 ± 2.33 (n = 97) |
73.30 ± 3.12 (n = 655) |
6.89 (p < 0.001) |
| Ширина яиц | 50.05 ± 0.98 (n = 558) |
49.68 ± 1.02 (n = 97) |
49.99 ± 1.57 (n = 655) |
2.15 (p = 0.03) |
Поведение выводков. Судя по прямым наблюдениям, а также возрасту встреченных выводков и срокам откладки яиц, вылупление в гнездах белощеких казарок в восточной части Финского залива происходило в период с начала 3-й пятидневки июня до конца 5-й пятидневки июля. Массовое появление птенцов в гнездах отмечалось во второй половине июня. Во время учетов выводки в возрасте до 5 дней встречались только у островов в местах гнездования птиц. Судя по учетам в конце июня и в июле, часть выводков более старшего возраста широко рассредоточивается вдоль побережья и по прибрежным островам. Так, в 2013–2015 гг. отдельные семьи белощеких казарок были встречены у северного побережья Финского залива и крупных прибрежных лесистых островов на расстоянии до 4–5 км от ближайшего известного места гнездования, а около многих прибрежных островов, где, судя по состоянию гнезд, произошло успешное вылупление, выводки старше пяти дней не отмечались. По нашим наблюдениям, выводки из гнезд на о-ве Стоглаз в возрасте около 5 дней перемещались через узкую протоку шириной 0.4 км к большому лесному о-ву Игривый и далее держались там на узких прибрежных луговинах и небольших островках вдоль берега этого острова. Обычно эти выводки держались группами из 2–5 семей. На островах, удаленных от берега на расстояние более 10 км (архипелаг Большой Фискар и Нерва), выводки держатся до подъема на крыло, что чаще всего происходит в первой декаде августа.
Линька
С 2013 г. стали отмечаться небольшие линочные скопления белощеких казарок у островов Нерва (10–12 особей), Долгий Риф (12–20 особей) и на архипелаге Большой Фискар (8–35 особей). Смена полетного оперения происходит в 3-й декаде июля–начале августа. Птицы держатся на наиболее выступающих в море плоских скалистых мысах. Здесь же отмечались и отдельные подросшие выводки.
ОБСУЖДЕНИЕ
Расселение белощеких казарок балтийской популяции в бореальной зоне северо-запада России началось значительно позже, чем в западных частях Балтийского региона. Впервые на Балтике казарки начали гнездиться на о-ве Готланд (Larsson et al., 1988; Black et al., 2014), затем расселение птиц шло в восточном и западном направлениях. В рамках этого процесса появление казарки в российской части Балтики после заселения финских и эстонских территорий кажется вполне закономерным, в то время как дальнейшее появление гнездящихся птиц на Онежском и Ладожском озерах выглядит продолжением процесса расселения вида. Мы полагаем, что этот процесс еще далек от завершения. Так, обнаружение размножающихся белощеких казарок на севере Ладожского оз. (Агафонова и др., 2016), в шхерах Онежского озера (Хохлова, Артемьев, 2015), в Кандалакшском заливе Белого моря (Шутова, Кожин, 2018) и на баренцевоморском побережье Кольского п-ова (Мельников, 2005; Мельников, Плотников, 2017), вполне возможно, демонстрирует происходящее прямо на наших глазах слияние балтийско-западноевропейского и арктического очагов размножения вида.
Если на Северном море казарки могут гнездиться не только на островах, но и на материке, то на Балтике птицы предпочитают исключительно острова (Feige et al., 2008). Такой выбор характерен и для востока Финского залива и связан с тем, что острова – это территории, как правило, свободные от наземных хищников. В России казарки гнездятся на небольших островках площадью 0.7–12.5 га. Сходного размера острова казарки заселяют и в других районах Балтики. Так, в Эстонии казарки гнездятся на островах площадью 2–5.5 га (Leito, 1996). В Германии казарки заселяют особенно маленькие острова площадью менее 0.02 га (Feige et al., 2008). В Швеции колонии располагаются на относительно плоских островах площадью не более 300 га (Ganter et al., 1999).
Примечательна особенность гнездования белощеких казарок в бореальной зоне Северо-Запада России почти исключительно в ландшафте сглаженных скал. Почти все известные случаи гнездования были на скалистых шхерных островах у северных побережий Финского залива, Ладожского и Онежского озер в местах выхода Балтийского кристаллического щита. Большинство птиц заселяло выровненные площадки и ложбины среднего яруса скал в локальных участках с хорошо развитой травянистой растительностью или под кустами. Ландшафты районов гнездования на Онежском и Ладожском озерах также весьма схожи с этими ландшафтами. Подобный тип гнездования белощеких казарок был доминирующим в Арктике в период сильного сокращения численности и гнездового ареала в середине 20-го века (Сыроечковский, 1995; Успенский, 1951; Bauer, Glutz, 1968; Cramp, Simmons, 1977), выровненные приморские тундры и плоские прибрежные острова вид стал заселять лишь по мере роста популяции в течение последних сорока лет (Волков, Чуприн, 1995; Гаврило, 1991; Калякин, 1986; Cыpoeчкoвский, 1995; Filchagov, Leonovich, 1992; Kondratyev et al., 2013).
Предпочтение скального ландшафта находится в существенном противоречии с распределением кормовых ресурсов. На скальных островах травянистая растительность представлена небольшими пятнами в понижениях субстрата и скальных трещинах. Из-за резкого свала глубин обильные водорослевые обрастания присутствуют обычно в полосе 2–5 м вокруг островов. Из водоплавающих птиц здесь предпочитают гнездиться преимущественно глубоководные ныряльщики: большой баклан, гага, гагарка и чистик (Коузов, Кравчук, 2020; Коузов, Шилин, 2016; Cherenkov et al., 2016; Shilin et al., 2014). В то же время у южного побережья Финского залива и в его центральной части присутствует большое количество небольших плоских моренных островов с обильной травянистой растительностью и примыкающими к ним обширными мелководными зонами с богатой водной растительностью (Kouzov et al., 2021; Zaynagutdinova et al., 2019). Эти острова активно используются для гнездования другими растительноядными гусеобразными птицами – лебедем-шипуном (Коузов, 2005, 2016; Коузов, Кравчук, 2014), серым гусем (Коузов, 2005а; Коузов, Кравчук, 2013) и серой уткой (Коузов, Кравчук, 2010а, 2012), однако казарки на таких островах не встречаются.
Сходная картина наблюдается и в более западных частях Финского залива, и у его горла. Так, на шхерных скалистых архипелагах вдоль побережья Финляндии в настоящее время размножается около 3000–3500 пар белощеких казарок (Valkama et al., 2011), а на низких моренных островах в прибрежной зоне Эстонии – только 113 пар (Feige et al., 2008). Возможно, выбор скальных островов для гнездования является результатом предпочтений вида, а не дефицита других гнездопригодных территорий.
При этом белощекие казарки демонстрируют большую пластичность в выборе микробиотопов на скальных островах. В последние годы казарки заселяют даже лесные участки, что на ранних этапах колонизации восточной части Финского залива не отмечалось, и птицы гнездились только на безлесных скальных уступах и в расщелинах (Kouzov et al., 2018; Zaynagutdinova et al., 2019). Подобные ландшафтные предпочтения можно объяснить тем, что островной скальный ландшафт был изначальным местом существования вида (Птушенко, 1952; Cыpoeчкoвский, 1995; Успенский, 1951; Bauer, Glutz, 1968; Cramp, Simmons, 1977), наиболее безопасным для сохранения кладок и птенцов от наземных хищников. Вероятно, при заселении новых территорий птицы сперва стремятся использовать именно этот наиболее безопасный для них ландшафт.
Из адаптаций белощекой казарки, позволяющих размножаться в условиях малой кормовой емкости скальных биотопов в восточной части Финского залива, можно указать явное предпочтение казарками при заселении наиболее близких к берегу островков. При этом выводки после вылупления широко распределяются вдоль побережья, что снижает кормовую конкуренцию. Казарки явно тяготеют к островам, наиболее приближенным к берегу, чтобы выводки перемещались на небольшие расстояния. В России половина гнезд казарок расположена на островах, находящихся на расстоянии не более 2 км от берега. Похожее расстояние до берега преодолевают казарки в Голландии – 70–1500 м (Feige et al., 2008). В Германии это расстояние еще меньше – 10–400 м до побережья (Feige et al., 2008). При этом в некоторых районах на Балтике половина выводков удаляется на расстояние более 3000 м от колонии (Feige et al., 2008).
Казарки, гнездящиеся на Северном и Балтийском море, покидая гнездовые территории после вылупления птенцов, в течение выводкового периода, как правило, кормятся в сельскохозяйственных ландшафтах или в городских парках (Feige et al., 2008; Väänänen et al., 2011). В России казарки с выводками после вылупления также покидают гнездовые территории и перемещаются на побережье, однако вынуждены проводить послегнездовой период на естественных приморских лугах (с ограниченным количеством кормовых участков), поскольку сельское хозяйство на северном берегу Финского залива развито очень слабо. Поэтому, вероятно, рост численности казарок на российской территории балтийского побережья незначителен. Так как сельскохозяйственные луга и пастбища с высококалорийными кормовыми ресурсами казаркам недоступны на территории Ленинградской обл., то и рост численности гнездовой популяции в восточной части Финского залива должен быть менее существенным, чем в более западных районах.
Средняя величина кладки в восточной части Финского залива была достоверно больше этого показателя в 9 из 10 выборок из различных районов Русской Арктики, полученных в разные годы (табл. 8). Существенное увеличение величин кладок у птиц Финского залива по сравнению с птицами, гнездящимися в Арктике, вероятно, связано с многократным сокращением весеннего миграционного пути и, соответственно, с сокращением энергетических затрат на перелет, величина которых оказывает ключевое влияние на репродуктивные кондиции арктических гусей (Drent et al., 2007; Hahn, 2011).
Таблица 8.
Средние величины кладок по данным из разных районов русской Арктики и величины t-критерия при их сравнении со средней величиной кладки в восточной части Финского залива (4.79 ± 1.13, n = 137)
| Район исследований | Источник данных | Годы | Величина кладки | Число кладок | Значения t-критерия при сравнении с нашими данными | Критическое значение t (p = 0.05) |
|---|---|---|---|---|---|---|
| п-ов Канин | Filchagov, Leonovich, 1992 | 1990–1991 | 4.61 ± 0.25 | 30 | 1.69 | 1.96 |
| о. Колгуев | Пономарева, 1992 | 1989–1990 | 3.99 ± 0.09 | 195 | 8.27 | 1.96 |
| о. Колгуев | Кондратьев и др., 2009 | 2006 | 3.61 ± 0.03 | 893 | 12.22 | 1.96 |
| о. Колгуев | Кондратьев и др., 2009 | 2007 | 3.36 ± 0.21 | 87 | 14.43 | 1.96 |
| о. Колгуев | Кондратьев и др., 2009 | 2008 | 3.42 ± 0.22 | 54 | 13.55 | 1.96 |
| Югорский п-ов | Filchagov, Leonovich, 1992 (Morozov В.В., устное сообщение) | 1990 | 4.08 ± 0.26 | 38 | 6.74 | 1.96 |
| о. Вайгач | Filchagov, Leonovich, 1992 (Morozov В.В., устное сообщение) | 1991 | 4.32 ± 0.12 | 109 | 4.83 | 1.96 |
| о. Вайгач | Filchagov, Leonovich, 1992 (Syroechkovsky E.E., устное сообщение) | 1988 | 4.0 2 ± 0.2 | 20 | 7.24 | 1.96 |
| о. Вайгач | Гуртовая, 1997 | 1995–1996 | 3.93 ± 0.19 | 59 | 8.63 | 1.96 |
| Колоколковская губа | Van der Jeugd et al., 2009 | 2002 | 2.77 ± 0.10 | 252 | 20.88 | 1.96 |
Динамические изменения в популяциях белощекой казарки, происходящие в последние десятилетия, свидетельствуют о том, что, несмотря на экспоненциальный рост численности вида и многократное расширение его гнездовой области, вышедшей далеко за пределы Арктики и Субарктики, процесс этот не является инвазивным, т.е. он не связан напрямую с непреднамеренной или преднамеренной интродукцией со стороны человека. Прямым доказательством тому является хорошо задокументированный естественный процесс вселения этого вида на о-в Готланд. Схема такова: задержка молодых птиц на весенней миграции – летование и линька неполовозрелых птиц в регионе – появление первых гнездящихся пар – рост гнездящейся группировки на островках восточного Готланда – расселение и появление новых колоний на других участках балтийских побережий (Forslund, Larsson, 1991; Larsson et al., 1988). Все случаи выпуска белощеких казарок из зоопарков Финляндии происходили гораздо позже начала этого естественного процесса и послужили “приятным дополнением” к нему, а не точкой его инициации (Väsänen et al., 1998; Väänänen et al., 2011).
Таким образом, появление на Балтике и на побережьях Северного моря вида, характерного для Субарктики, экстраординарным событием не является. На побережьях Балтийского моря известны места размножения морской чернети, синьги, турпана, морянки, камнешарки, белохвостого песочника, полярной крачки и короткохвостого поморника (Cramp, Simmons, 1977, 1983; Curry-Lindahl, 1964; Hilden, 1987; Rutschke, 1989). Ареалы обыкновенной гаги, гагарки, чистика и тонкоклювой кайры от Балтики доходят до Ла-Манша и Бретани (Cramp, Simmons, 1977, 1983; Rutschke, 1989). А такие тундровые виды как галстучник, золотистая ржанка и чернозобик в регионе Балтики образуют даже устойчивые обособленные “южные подвиды” (Cramp, Simmons, 1983). Все эти случаи также являются следствиями не только смягчающего действия приморского климата и наличия большого количества прибрежных азональных и интразональных местообитаний, имеющих довольно сходный облик в различных климатических зонах, но и следствием долговременной динамики климата, обусловливавшей существенную пульсацию ареалов этих видов птиц в Голоцене (Кривенко, 1991). И единственным отличием в ситуации с белощекой казаркой является то обстоятельство, что процесс происходит прямо на наших глазах. А это, в свою очередь, и представляет огромный интерес для науки как живая модель для изучения механизмов расселения (Guo, 2014).
В настоящее время процесс расселения и популяционного роста белощекой казарки вызывает многочисленные вопросы, на которые пока нет однозначных ответов. Несомненно, драйверами этого процесса послужили развитие теплой фазы климата и усиление мер по охране вида на зимовках, а также увеличение использования удобрений в сельском хозяйстве (Розенфельд, Шереметьев, 2016; Розенфельд и др., 2021; Cыpoeчкoвский, 1995; Madsen et al., 1999; Prop, de Vries, 1993). Однако расширение ареала вида при развитии теплой фазы климата шло преимущественно с северо-востока на юго-запад, т.е. в направлении, противоположном направлению изменений основных климатических факторов (сроков наступления весны, средних летних температур и длительности безморозного периода) (Розенфельд, Шереметьев, 2016; Розенфельд и др., 2021; Cыpoeчкoвский, 1995; Madsen et al., 1999). Так, в конце многовекового холодного периода – в первой половине 20-го века – рефугиум российской популяции белощекой казарки находился в самом холодном восточном углу Баренцевоморского бассейна на островах Вайгач и Новая Земля (Розенфельд и др., 2021; Cыpoeчкoвский, 1995). Отсюда птицы стали расселяться на более западные участки прибрежной зоны Баренцева моря, где даже при климатическом пессимуме условия были существенно лучше благодаря действию Гольфстрима (по данным исследований тех же авторов). При этом появление новых мест гнездования в районах весенних миграционных остановок на Балтике началось практически одновременно с событиями в Арктике (Cыpoeчкoвский, 1995; Filchagov, Leonovich, 1992; Forslund, Larsson, 1991; Larsson et al., 1988).
Рядом авторов высказывается вполне обоснованная гипотеза, что подобный странный рефугиум образовался в результате прямого преследования вида со стороны человека на морских побережьях и островах более западных частей Баренцева моря и что нынешнее расселение птиц является лишь восстановлением старого ареала в результате усиления мер по охране вида (Розенфельд и др., 2021; Cыpoeчкoвский, 1995). Однако появление балтийско-западноевропейского очага размножения вида эта гипотеза не объясняет. Кроме того, в таком случае непонятно, почему в первой половине и в середине 20-го века не было даже попыток гнездования вида на островах заповедников восточного и северного побережья Кольского п-ова (Гавриловские острова, архипелаг Семь Островов и Айновы о-ва), где климатические условия еще мягче, чем на п-ове Канин и о-ве Колгуев. Заселять Восточный Мурман белощекие казарки стали в самую последнюю очередь, и до сих пор численность их здесь относительно невысока (Мельников, 2005; Мельников, Плотников, 2017).
Не меньший интерес представляет масштабная смена биотопических предпочтений белощекой казарки при расселении в новые районы гнездования – вместо скальных уступов и речных обрывов Вайгача и Новой Земли (Кaлякин, 1986; Мензбир, 1895; Птушенко, 1952; Cramp, Simmons, 1977; Prop et al., 1984) вид стал активно заселять низменные островки, приморские косы и лайды, а также участки сырых тундр (Гаврило, 1991; Карагичева, 2011; Минеев, Минеев, 2004; Пономарева, 1992; Розенфельд и др., 2021; Cыpoeчкoвский, 1995; Фильчагов, 1997; Filchagov, Leonovich, 1992; Glazov et al., 2021; Gurtovaya, Litvin, 2001; Karagicheva et al., 2011). Ранее считалось, что скальное гнездование является изначальной характерной чертой вида, связанной с адаптациями к сохранению кладок от наземных разорителей (Гуртовая, 1997; Мензбир, 1895; Птушенко, 1952; Cыpoeчкoвский, 1995; Cramp, Simmons, 1977). В рамках гипотезы об образовании рефугиума в восточном секторе Баренцева моря в результате антропогенного преследования высказывается обоснованное предположение, что гнездование на скалах и речных обрывах было вынужденным. Такой выбор происходил из-за того, что в условиях очень позднего развития весенних событий именно эти участки освобождаются из-под снега первыми, и что при потеплении климата и восстановлении прежнего западно-баренцевоморского участка ареала этот лимит был снят (Розенфельд и др., 2021).
При вселении белощекой казарки на побережья Балтийского моря мы видим, что первые ее поселения на Готланде и на островах Западно-Эстонского архипелага также появились в сходных биотопах – на низменных выровненных островках с низкотравной растительностью (Cыpoeчкoвский, 1995; Black et al., 2014; Forslund, Larsson, 1991; Larsson et al., 1988; Leito, 1996; van der Graaf et al., 2007). При дальнейшем расселении на юг и юго-запад в Северной Германии и Голландии птицы продолжали гнездиться в низменных приморских местообитаниях (Cыpoeчкoвский, 1995; Feige et al., 2008). Однако при распространении на северо-восток, достигнув краев Балтийского кристаллического щита на Финском заливе, птицы начинают размножаться почти исключительно в скальном возвышенном ландшафте, избегая низких выровненных моренных островов (Kouzov et al., 2018; Väänänen et al., 2011). Исходя из вышесказанного видно, что выходы скал остаются предпочтительным для вида гнездовым биотопом, но бурный популяционный рост как бы выталкивает избыток птиц в новые, более распространенные местообитания.
Потепление климата в Арктике привело к более раннему прилету на места гнездования белощеких казарок и более позднему отлету с зимовок (Eichhorn et al., 2006; Lameris et al., 2018). Таким образом значительно сократилось время пребывания на промежуточных остановках (Розенфельд и др., 2021; Eichhorn et al., 2006). Однако на Северо-Западе России изменения миграционных стратегий белощеких казарок имеют противоположную направленность. Миграции птиц начинаются приблизительно на месяц раньше, чем 30 лет назад (Мальчевский, Пукинский, 1983). При этом идет образование новых миграционных стоянок на островах Финского залива (Коузов, 2010, 2015; Коузов, Лосева, 2014), на полях восточного Приладожья в районе г. Олонец (Артемьев и др., 2009, 2011, 2013; Лапшин и др., 2016), Прионежья у г. Шуя (Артемьев и др., 2010) и в дельте Северной Двины (Андреев, 2005).
Изменения миграционного поведения птиц можно объяснить тем, что емкость старых миграционных стоянок уже недостаточна для вмещения растущей популяции, и часть птиц вынуждена искать новые места остановок вдоль миграционного пути (Eichhorn et al., 2009). В результате миграционная стратегия вида меняется: часть птиц по-прежнему совершает дальний поздний безостановочный бросок на п-ов Канин после длительной стоянки в Западной Эстонии (Leito, 2008; Van der Graaf et al., 2006), в то время как другая часть перемещается короткими бросками после непродолжительных остановок, двигаясь “за зеленой волной” (Najafabadi et al., 2014, 2015), подобно гусям рода Anser (Drent et al., 1978; Van der Graaf et al., 2006). Этому способствует переход белощеких казарок на кормление на полях совместно с гусями (Артемьев и др., 2009, 2011, 2013; Лапшин и др., 2016; Розенфельд и др., 2021). Для сравнения, черная казарка, на зимовках и миграционных остановках питающаяся преимущественно морским растением зостерой Zostera marina (Розенфельд, Шереметьев, 2016; Cramp, Simmons, 1977; Hassall et al., 2001), до настоящего времени вынуждена мигрировать одним дальним броском из Западной Эстонии в дельту Северной Двины в конце мая – начале июня (Мальчевский, Пукинский, 1983; Коузов, 2010; Андреев, 2005) во многом потому, что зостера между данными местами стоянок нигде более не произрастает.
На наш взгляд, вышеуказанные особенности экспансии белощекой казарки из арктических районов в места с более мягким климатом, практически синхронное с расселением в Арктике ее расселение в регионе Балтийского и Северного морей, а также существенная смена биотопического распределения и миграционной стратегии не могут быть объяснены только воздействием на птиц динамики климата и усиления мер по ее охране на местах зимовки и миграционных остановок. Подобные события, возможно, связаны с проявлением активно идущего микроэволюционного процесса (Бигон и др., 1989; Guo, 2014). Так, в конце 19-го века–первой половине 20-го столетия, под воздействием климатического пессимума и усиления антропогенного пресса вид, вероятно, проходил через эволюционное “бутылочное горлышко”. В результате в популяции накопилось определенное количество особей с широкой нормой реакции на фотопериодические и ландшафтно-климатические условия среды. И именно за счет этих птиц, при улучшении жизненных условий начались взрывной рост численности вида и практически синхронное расселение его в местах прежних миграционных остановок с весьма различными условиями обитания.
Следует обратить внимание, что большинство районов размножения казарок на Балтийском и Северном море являются ООПТ (Feige et al., 2008). В Швеции половина мест гнездования казарок представляет собой ООПТ, другая половина расположена в районах, которые использовались ранее в военных целях, и фактор беспокойства здесь до сих пор невысок (Feige et al., 2008), в Финляндии половина гнезд также находится на ООПТ (Valkama et al., 2011; Väänänen et al., 2011).
В российской части Финского залива казарки также гнездятся преимущественно в местах, которые до недавнего времени были закрыты для посещения (приграничная зона) и где уровень беспокойства был невысок. Однако после снятия ряда ограничений пограничного режима в 2017 г. здесь значительно чаще появляются туристы и возрастает уровень беспокойства для птиц.
В настоящее время существуют серьезные проблемы с охраной мест гнездования белощекой казарки в регионе, несмотря на то что большая часть из них формально входит в состав ООПТ. Вне состава ООПТ находятся только острова Нерва, Гусиный, Малый Фискар и Туман. Остров Стоглаз входит в состав регионального природного парка “Кивипарк”, однако штата охраны здесь нет. Бо́льшая часть других островов у северного побережья Финского залива входит в состав организованного в 2017 г. федерального заповедника “Восток Финского залива”, однако до сих пор у заповедника отсутствуют штат охраны и инфраструктура. Таким образом, первостепенными задачами сохранения вида в регионе являются организация реального режима охраны на всех уже существующих ООПТ и организация региональных ООПТ на островах Нерва, Гусиный, Малый Фискар и Туман.
Список литературы
Агафонова Е.В., Михалёва Е.В., Соколовская М.В., 2016. Гнездование канадской Branta canadensis и белощёкой B. leucopsis казарок на островах Валаамского архипелага Ладожского озера // Русский орнитологический журнал. Т. 25. Экспресс-выпуск 1257. С. 801‒ 802.
Андреев В.А., 2005. Экологические особенности миграций гусеобразных в устьевой части Северной Двины. Автореф. дис. … канд. биол. наук. М. 20 с.
Артемьев А.В., Зимин В.Б., Лапшин Н.В., Симонов С.А., 2009. Особенности динамики весенних скоплений гусеобразных птиц Anseriformes на Олонецких полях Карелии в 2009 году // Русский орнитологический журнал. Т. 18. Экспресс-выпуск 522. С. 1891‒ 1899.
Артемьев А.В., Зимин В.Б., Лапшин Н.В., Симонов С.А., Логинов И.И., 2011. Особенности динамики весенних скоплений гусеобразных Anseriformes на Олонецких полях Карелии в 2010 и 2011 годах // Русский орнитологический журнал. Т. 20. Экспресс-выпуск 706. С. 2293‒2300.
Артемьев А.В., Симонов С.А., Лапшин Н.В., Логинов И.И., 2013. Особенности динамики весенних скоплений гусеобразных птиц Anseriformes на Олонецких полях Карелии в 2013 году // Русский орнитологический журнал. Т. 22. Экспресс-выпуск 933. С. 2948‒ 2953.
Артемьев А.В., Зимин В.Б., Лапшин Н.В., Хохлова Т.Ю., 2010. Весенняя орнитофауна агроценозов поселка Шуя // Изучение динамики популяций мигрирующих птиц и тенденций их изменений на Северо-Западе России. СПб: Тускарора. Вып. 8. С. 30‒38.
Белопольский Л.О., 1956. Некоторые данные о полете птиц осенью 1954 года на побережье Белого моря и перспективах организации наблюдений за миграциями птиц в Карело-Финской ССР. Ежегодник Общества естествоиспытателей при Академии наук Эстонской ССР. Т. 49. С. 65‒73.
Бигон М., Харпер Дж., Таунсенд К., 1989. Экология: особи, популяции и сообщества. Т. 2. М.: Мир. 450 с.
Бубырева В.А., Бузун В.А., Волкович Н.М., Коузов С.А., Шаповалова О.В., Щукин А.К., 1993. Отчет Кургальской экспедиции Санкт-Петербургского Общества Естествоиспытателей в полевой сезон 1992 г. // Вестник СПбГУ. Сер. 3. Вып. 2(10). С. 111‒117.
Бузун В.А., 1997. Остров Сескар (восточная часть Финского залива). Материалы первого семинара по программе “Изучение состояния популяций мигрирующих птиц и тенденций их изменений в России”. СПб.: Тускарора. С. 42‒49.
Бузун В.А., 1998. Данные о миграции птиц на острове Сескар (Финский залив, Балтийское море) весной 1997 г. // Материалы второго семинара по программе “Изучение состояния популяций мигрирующих птиц и тенденций их изменений в России” (18–20 февраля 1998 г., Москва). М.: ГУП ВТИИ. С. 47‒69.
Волков А.Е., 2013. Наблюдения за весенним пролетом птиц на Онежском полуострове в 2013 году // Русский орнитологический журнал. Т. 22. Экспресс-выпуск 931. С. 2875‒2892.
Волков А.Е., Брагин А.В., Покровская И.В., Волкова Е.В., 2015. Весенний пролёт гусей и лебедя-кликуна на Онежском полуострове в 2014 г. // Казарка – Бюллетень Рабочей Группы по Гусеобразным Северной Евразии. Вып. 18. С. 91‒103.
Волков А.Е., Чуприн И.И., 1995. Новые сведения о гнездовании белощекой казарки (Branta leucopsis) на острове Колгуев // Бюллетень РГГ. № 1. С. 47‒50.
Гаврило М.В.,1991. Белощекая казарка на острове Колгуеве // Материалы 10-й всесоюз. орнитологической конференции. Минск. Ч. 2. С. 128.
Головнюк В.В., Поповкина А.Б., Соловьёв М.Ю., 2015. Первый случай гнездования белощёкой казарки на Таймыре // Казарка. Вып. 18. С. 25‒29.
Гагинская А.Р., Носков Г.А., Резвый С.П., 1997. О находке гнезда белощёкой казарки на Финском заливе // Казарка. Вып. 3. С. 111‒112.
Гуртовая Е.Н., 1997. Условия гнездования белощеких казарок на ocтpoвe Вайгач в 1996 году // Казарка. Вып. 3. С. 109‒110.
Иовченко Н.П., Гагинская А.Р., Носков Г.А., Резвый С.П., 2002. Результаты орнитологического обследования островов Финского залива в 1994‒1995 годах // Птицы и млекопитающие Северо-Запада России. Труды Биологического НИИ. Т. 48. С. 100‒120.
Kaлякин B.H., 1986. O pacпpocтpaнeнии и экoлoгии бeлoщeкoй кaзapки нa Ocтpoвe Baйгaч и Югopcкoм пoлyocтpoвe // Aктyaльныe пpo6лeмы opнитoлoгии. M.: Hayкa. C. 93‒103.
Карагичева Ю.В., 2011. Особенности экологии белощёкой казарки (Branta leucopsis) при освоении новых местообитаний в Субарктике. Автореф. дис. … канд. биол. наук. М. 24 с.
Кищинский А.А., 1979. Миграции белощекой казарки – Branta leucopsis (Bechst) // Миграции птиц Восточной Европы и Северной Азии (Аистообразные – Пластинчатоклювые). М.: Наука. 229 с.
Кондратьев А.В., Зайнагутдинова Э.М., Крукенберг Х., 2009. Современный статус и биология гусей на острове Колгуеве // Казарка. Т. 15. Вып. 2. С. 31‒70.
Коузов С.А., 1995. Новые сведения о некоторых видах казарок в Восточной части Финского залива // Бюллетень рабочей группы по гусям Восточной Европы и Северной Азии. № 1. С. 174‒177.
Коузов С.А., 2005. Адаптации к открытым морским мелководьям у лебедей-шипунов, гнездящихся на Кургальском полуострове (восточная часть Финского залива). Материалы 111 Междунар. симпозиума “Гусеобразные Северной Евразии”. Санкт-Петербург. С. 160‒162.
Коузов С.А., 2005а. Адаптации к открытым морским мелководьям у серых гусей, гнездящихся на Кургальском полуострове (восточная часть Финского залива). Материалы 111 Междунар. симпозиума “Гусеобразные Северной Евразии”. Санкт-Петербург. С. 162‒163.
Коузов С.А., 2009. Летне-осенние скопления и транзитные миграции водно-болотных птиц на Кургальском полуострове в 2007 г. // Изучение динамики популяций мигрирующих птиц и тенденций их изменений на Северо-Западе России. Вып. 6. С. 71‒87.
Коузов С.А., 2010. Весенняя миграция водно-болотных птиц на Кургальском полуострове в 2008 г. // Изучение динамики популяций мигрирующих птиц и тенденций их изменений на Северо-западе России. Вып. 7. С. 42‒59.
Коузов С.А., 2011. Миграции белощекой и черной казарок через Кургальский полуостров и сопредельные участки восточной части Финского залива. Тезисы 4-го совещания рабочей группы по гусеобразным Северной Евразии “Гусеобразные Северной Евразии”. Элиста 24–29 марта 2011 г.
Коузов С.А., 2015. О массовых стоянках белощёкой казарки Branta leucopsis на Кургальском полуострове весной 2008 и 2015 годов // Русский орнитологический журнал. Т. 24. № 1194. С. 3463‒3467.
Коузов С.А., 2016. Лебедь-шипун (Cygnus olor Gmelin 1789) в восточной части Финского залива: история расселения, распределение размножающихся птиц и биология размножения // Вестник Санкт-Петербургского университета. Серия 3. Биология. № 2. С. 38‒69.
Коузов С.А., Кравчук А.В., 2008. Первый случай гнездования белощёкой казарки Branta leucopsis на Кургальском полуострове // Русский орнитологический журнал. Т. 17. № 423. С. 908‒910.
Коузов С.А., Кравчук А.В., 2010. Наблюдения за миграциями водно-болотных птиц и миграционными стоянками 16.05‒04.06.2009 г. на Березовых островах // Изучение динамики популяций мигрирующих птиц и тенденций их изменений на Северо-западе России. Вып. 8. С. 83‒88.
Коузов С.А., Кравчук А.В., 2010а. Гнездование серой утки в Ленинградской области // Динамика популяций охотничьих животных Северной Европы. V Международный симпозиум 1‒5 сентября 2010 г. Рабочеостровск, Карелия, Россия. Петрозаводск. С. 134‒135.
Коузов С.А., Кравчук А.В., 2012. Серая утка в восточной части Финского залива: история заселения, биология и миграции // Бюллетень РГГ “Казарка”. Вып. 15. № 2. С. 106‒139.
Коузов С.А., Кравчук А.В., 2013. Серый гусь (Anser anser) в Ленинградской области: основные тенденции многолетних изменений численности, экология, миграции и перспективы реинтродукции // Вестник охотоведения. Т. 10. № 1. С. 5–16.
Коузов С.А., Кравчук А.В., 2014. Биология лебедя-шипуна (Cygnus olor) в восточной части Финского залива // Вестник охотоведения. Т. 11. № 2. С. 199‒ 203.
Коузов С.А., Кравчук А.В., 2020. Большой баклан Phalacrocorax carbo в восточной части Финского залива: долговременная динамика численности, распределение и роль в местных экосистемах // Русский орнитологический журнал. Т. 29. № 2014. С. 6037‒ 6038.
Коузов С.А., Лосева А.В., 2014. О локальных путях пролета и миграционных стоянках морских птиц в восточной части Финского залива по результатам судовых учетов 2010–2013 гг. // Вестник охотоведения. Т. 11. № 2. С. 204‒210.
Коузов С.А., Лосева А.В., 2014а. О локальных путях пролета и миграционных стоянках морских птиц в восточной части Финского залива по результатам судовых учетов 2010–2013 гг. // Динамика популяций охотничьих животных Северной Европы. Тезисы докладов VI Международного симпозиума. 31 марта–4 апреля 2014 г. Черные камни, Карелия, Россия, Петрозаводск. С. 55‒56.
Коузов С.А., Шилин М.В., 2016. Основные тенденции многолетней динамики сообществ гидрофильных птиц островной зоны восточной части Финского залива // Русский орнитологический журнал. Т. 25. Экспресс-выпуск 1257. С. 799‒801.
Кривенко В.Г., 1991. Водоплавающие птицы и их охрана. М.: Агропромиздат. 270 с.
Кумари Э., 1963. Динамика численности некоторых мигрирующих морских птиц на Белом море и в восточной части Балтийского моря. Сообщения Прибалтийской Комиссии по изучению миграции птиц. Т. 2. С. 67‒80.
Лакин Г.Ф., 1990. Биометрия. М.: Высшая Школа. 350 с.
Лапшин Н.В., Артемьев А.В., Симонов С.А., 2016. Численность белощекой казарки Branta leucopsis (Bechst) на весенних стоянках и ее статус в южной Карелии // Труды Карельского научного центра РАН. № 7. С. 85–92.
Мальчевский А.С., Пукинский Ю.Б., 1983. Птицы Ленинградской области и сопредельных территорий: История, биология, охрана. Т. 1. Л.: Изд. ЛГУ. 480 с.
Мельников М.В., 2005. Первый случай гнездования белощёких казарок Branta leucopsis на Восточном Мурмане // Русский орнитологический журнал. Т. 14. № 304. С. 1022–1023.
Мельников М.В., Плотников Р.В., 2017. Летопись природы по району архипелага Семь островов, Баренцево море, за 2016 год // Летопись природы Кандалакшского заповедника. Кн. 62. С. 7–161.
Мензбир М.А., 1895. Птицы России, 1. М. 836 с.
Минеев Ю.Н., 1984. О гнездовании белощекой казарки на Югорском полуострове // Oрнитология. Вып. 19. С. 183.
Минеев Ю.Н., Минеев О.Ю., 2004. Современное состояние малоземельской популяции белощекой казарки Branta leucopsis // Русский орнитологический журнал. Т. 13. Экспресс-выпуск 267. С. 655–661.
Нейфельдт И.А., 1970. Обзор орнитологических исследований в Карелии. Труды Зоологического института Академии Наук СССР. Т. 47. С. 67‒110.
Носков Г.А., Федоров В.А., Гагинская А.Р., Сагитов Р.А., Бузун В.А., 1993. Орнитофауна островов восточной части Финского залива. Русский орнитологический журнал. Вып. 2 (2). С. 163‒173.
Пономарева Т.С., 1991. Белощекая казарка в окрестностях острова Колгуева // Материалы 10-й Всесоюзной орнитологической конференции. Минск: Наука и техника. Ч. 2, кн. 2. С. 157–158.
Пономарева Т.С., 1992. Гнездование белощекой казарки в районе острова Колгуев // Бюллетень московского общества испытателей природы. Вып. 98. № 2. С. 39–44.
Птушенко Е.С., 1952. Отряд Гусеобразные. Подсемейство Гусиные. В кн.: Дементьев Г.П., Гладков Н.А. Птицы Советского Союза. М.: Советская Наука. С. 247–344.
Розенфельд С.Б., Шереметьев И.С., 2016. Арктические гуси (Anser) и казарки (Branta) Евразии: анализ факторов динамики численности и ареалов // Журнал общей биологии. Т. 77. № 1. С. 16–37.
Розенфельд С.Б., Волков С.В., Рогова Н.В., Киртаев Г.В., Соловьев М.Ю., 2021. Влияние изменений условий гнездования в Арктике на экспансию российской популяции белощекой казарки (Branta leucopsis) // Зоологический журнал. Т. 100. № 5. С. 510–523.
Рымкевич Т.А., Носков Г.А., Коузов С.А., Уфимцева А.А., Зайнагутдинова Э.М., Стариков С.А., Рычкова А.Л., Иовченко Н.П., 2012. Результаты синхронных учетов мигрирующих птиц в Невской губе и на прилежащих акваториях весной 2012 года // Изучение динамики популяций мигрирующих птиц и тенденций их изменений на Северо-Западе России. Вып. 9. С. 68–79.
Cыpoeчкoвский Е.Е. мл., 1995. Hoвое в гнездовом распространении белощеких казарок в России // Казарка. Бюллетень РГГ. № 1. С. 39–45.
Успенский С.М., 1951. Гнездовья белощекой казарки на Новой Земле // Охрана природы. М. С. 124–127.
Фильчагов А.В., 1997. Массовое повторное гнездование белощёких казарок Branta leucopsis на полуострове Канин, Россия // Казарка. № 3. С. 101–197.
Харитонов С.П., 2018. Материалы мониторинга орнитофауны участка “Бухта Медуза” // Летопись природы ФГБУ “Заповедники Таймыра”, 2018 г. Кн. 6. Норильск. С. 187–213.
Хохлова Т.Ю., Артемьев А.В., 2015. Первая регистрация гнездования белощекой казарки Branta leucopsis на Онежском озере // Русский орнитологический журнал. Т. 24. Вып. 1152. С. 2021–2024.
Храбрый В.М., Байбекова С.А., 2016. Гнездование белощёкой казарки Branta leucopsis в Выборгском заливе (Ленинградская область) // Русский орнитологический журнал. Т. 25. Экспресс-выпуск 1235. С. 80–81.
Шутова Е.В., Кожин М.Н., 2018. Белощёкая казарка в Кандалакшском заливе Белого моря // Казарка. № 20. С. 127–129.
Alsos I.G., Elvebakk A., Gabrielsen G.W., 1998. Vegetation exploitation by barnacle geese Branta leucopsis during incubation on Svalbard // Polar Research. V. 17. P. 1–14.
Black J., Prop J., Larsson K., 2014. The Barnacle Goose. London: T & AD Poyser. 287 p.
Bauer K., Glutz U.-N., 1968. Handbuchder Vogel Mittel Europas. Hrsg. VonG. Neuthammer. 2. Anseriformes. FrankfurtunMain: Akad. Verl.-Ges. P. 5–356.
Boyd H., 1961. The number of Barnacle geese in Europe in 1959–60 // Wildfowl Trust Ann. Rep. 12. P. 116–124.
Cherenkov A.E., Kouzov S.A., Semashko V.Y., Tertitski G.M., Semashko E.V., 2016. Present status of Razorbills Alca torda in Russia: occurrence, population and migrations // Marine Ornithology. V. 44. № 2. P. 207–213.
Cramp S., Simmons K.E.L. (eds), 1977. The Birds of the Western Palearctic. Vol. 1: Ostrich to Ducks. Oxford–London–New-York: Oxford University Press. P. 1–722.
Cramp S., Simmons K.E.L. (eds), 1983. Birds of Western Palearctic. Vol. 3. Oxford–London–New York: Oxford University Press. 913 p.
Curry-Lindahl K., 1964. The situation of ducks, geese and swans in Norway, Sweden and Finland. Proc. Est. Europ. Meet. Wildfowl Conservat., St. Andrews, 1963. London: Nature Conservancy. P. 62‒73.
Drent R., Ebbinge B., Weijand B., 1978. Balancing the energy budget of arctic-breeding geese throughout the annual cycle: a progress report // Verhandlungen der Ornithologische Gesellschaft in Bayern. V. 23. P. 239‒264.
Drent R.H., Eichhorn G., Flagstad A., Van der Graaf A.J., Litvin K.E., Stahl J., 2007. Migratory connectivity in Arctic geese: spring stopovers are the weak links in meeting targets for breeding // Journal of Ornithology. V. 148. № 2. P. 501–514.
Eichhorn G., Afanasyev V., Drent R.H., van der Jeugd H.P., 2006. Spring stopover routines in Russian Barnacle Geese Branta leucopsis tracked by resightings and geolocation // Ardea. Vol. 94. P. 667‒678.
Ganter B., Larsson K., Syroechkovsky E.V., Litvin K.E., Leito A., Madsen J., 1999. Barnacle Goose Branta leucopsis: Russia // Baltic. In: Madsen J., Cracknell G., Fox A.D. (eds) Goose populations of Western Paleartic. A review of status and distribution. Wetlands International Publishing. Vol. 48. Wetlands International, Wageningen, The Netherlands. National Environmental Reseach Institute, Rönde, Denmark. P. 270–283.
Glazov P.M., Loshchagina J.A., Kondratyev A.V., Zaynagutdinova E.M., Kruckenberg H., Pokrovsky I.G., 2021. The long-term monitoring of bird populations on Kolguev island in the Barents sea // Arctic. V. 74 (Suppl. 1). P. 23–40.
van der Graaf S.A.J., Stahl J., Klimkowska A., Bakker J.P., Drent R.H., 2006. Surfing on a green wave – how plant growth drives spring migration in the Barnacle goose Branta leucopsis // Ardea. V. 94. P. 567–577.
van der Graaf S.A.J., Stahl J., Veeneklaas R.M.A., Bakker J.P., 2007. Vegetation characteristic of brackish marsh on Gotland and foraging choises of migraring and brood rearing geese // Ann. Bot Fennici. V. 44. P. 33‒41.
Gurtovaya E.N., Litvin K.E., 2001. Changes in Barnacle Goose nest distribution on Vaygach Island (1986–1997). Proc. 6th Annual Meeting of the Goose Specialist Group of Wetlands International. P. 28‒29.
van der Jeugd H.P., Eichhorn G., Litvin K.E., Stahl J., Larsson K., van der Graaf S.A.J., Drent R.H., 2009. Keeping up with early springs: rapid range expansion in an avian herbivore incurs a mismatch between reproductive timing and food supply // Global Change Biology. V. 15. Issue 5. P. 1057‒1071.
Feige N., van der Jeugd H.P., van der Graaf S.A.J., Larsson K., Leito A., Stahl J., 2008. Newly established breeding sites of the Barnacle Goose Branta leucopsis in North-western Europe – an overview of breeding habitats and colony development // Vogelwelt. № 129. P. 244–252.
Filchagov A.V., Leonovich V.V., 1992. Breeding range expansion of Barnacle and Brent Geese in the Russian European North // Polar Research. V. 11. № 2. P. 41‒46.
Forslund P., Larsson K., 1991. Breeding range expansion of the Barnacle Goose Branta leucopsis in the Baltic area // Ardea. V. 79. P. 343‒346.
Fox A.D., Madsen J., 2017. Threatened species to superabundance: The unexpected international implications of successful goose conservation // Ambio. V. 46 (Suppl. 2). P. 179–187.
Hahn S., Loonen M.J.J.E., Klaassen M., 2011. The reliance on distant resources for egg formation in high Arctic breeding barnacle geese Branta leucopsis // J. Avian Biol. V. 42. P. 159‒168.
Hassall M., Riddington R., Helden A., 2001. Foraging behaviour of brent geese, Branta b. bernicla, on grasslands: effects of sward length and nitrogen content // Oecologia. V. 127. P. 97–104.
Hilden O., 1987. Recent changes in the sea-bird populations of Finland // Водно-болотные угодья и водоплавающие птицы. Таллин. С. 74–83.
Karagicheva J., Rozenfeld S., van der Jeugd H., Rakhimberdiev E., 2011. A pilot analysis of diet composition of Barnacle Geese Branta leucopsis at a sub-arctic salt marsh on the Kanin peninsula, Russia // Goose Bulletin. V. 13. P. 3–7.
Kondratyev A., Zaynagutdinova E., Kruckenberg H., 2013. Barnacle Goose Branta leucopsis abundance on Kolguev Island – current status and history of population growth // Wildfowl. V. 63. P. 56–71
Kontiokorpi J., Rusanen P., 2014. Survey of springtime arctic bird migration in Vyborg in 1988–2008 and in Kurortny District in 1992–2001 // Reports of the Finnish Environment institute. P. 39.
Koop B., 1998. Die Brutansiedlung und Bestandentwicklung der Weißwangengans Branta leucopsis in Schleswig-Holstein // Limicola. V. 12. P. 72–76.
Kouzov S., Zaynagutdinova E., Sagitov R., Rychkova A., 2018. Nesting of Barnacle Goose (Branta leucopsis) in the Russian part of the Gulf of Finland // Arctic. V. 71. № 1. P. 76–88.
Kouzov S., Zaynagutdinova E., Kravchuk A.,2019. Late nesting makes Barnacle Geese Branta leucopsis sensitive to anthropogenic disturbance in the Russian part of the Baltic Sea // Wildfowl. V. 69. P. 160‒175.
Kouzov S.A., Gubelit, Y.I., Kravchuk A.V., Koptseva E.M., Zaynagutdinova E.M., Nikitina V.N., 2021. Seasonal changes in the diet of Mute Swans Cygnus olor in the recently colonised eastern Gulf of Finland // Wildfowl. V. 71. P. 83‒107.
Kruckenberg H., Hasse T., 2004. Nonnenganse (Branta leucopsis) als Brutvogel an der Unterems // Vogelkdl. Ber. Niedersachsen. V. 36. P. 83–88 [In German].
Lameris T.K., van der Jeugd H.P., Eichhorn G., Dokter A.M., Bouten W., Boom M.P., Litvin K.E., Ens B.J., Nolet B.A., 2018. Arctic geese tune migration to a warming climate but still suffer from a phenological mismatch // Current Biology. V. 28. P. 1‒7.
Larsson K., Forslund P., Gustafsson L., Ebbinge B., 1988. From the high Arctic to the Baltic: the successful establishment of a Barnacle Goose Branta leucopsis population on Gotland, Sweden // Ornis Scandinavica. V. 19. P. 182–189.
Leito A.,1996. The Barnacle Goose in Estonia // Estonia Maritime. V. 1. P. 1–103.
Leito A., 2008. Internationally important staging areas for geese in Estonia // Vogelwelt. V. 129. P. 191‒194.
Madsen J., Cracknell G., Fox A.D., 1999. Goose populations of the Western Palearctic. A review of status and distribution // Wetlands International Publication. V. 48. P. 1‒344.
Meinger P.L., van Swelm N.D., 1994. Brandganzen Branta leucopsis als broedvogel in het Deltagebied // Limosa. V. 67. P. 1–5.
Mortensen E., Hansen M., 1999. Ynglefugle på Saltholm 1998–1999. In: Købenshavns Amt og Købenshavns Lufthavne: Natur for waltnings rapport nr. 32, Kopenhagen. P. 49.
Najafabadi S.M., Wang T., Skidmore A.K., Toxopeus A.G., Kölzsch A., et al., 2014. Migratory Herbivorous Waterfowl Track Satellite-Derived Green Wave Index // PLoS ONE. V. 9(9). P. e108331.
Najafabadi S.M., Darvishzadeh R., Skidmore A.K., Kölzsch A., Vrieling A., Nolet B.A., Exo K.-M., Meratnia N., Havinga P.J.M., Stahl J., Toxopeus Al.G., 2015. Satellite-versus temperature-derived green wave indices for predicting the timing of spring migration of avian herbivores // Ecological Indicators. V. 58. P. 322–331.
Olsen K.M., 1992. Danmarks fugle – en oversigt. Dansk Ornithol. Forening, Kobenhaven.
Ouweneel G.L., 2001. Snelle groei van de broedpopulatie Brandganzen Branta leucopsis in het deltagebied // Limosa. V. 74. P. 137–146.
Putkonen T.A., 1940. Tiedonantoja (Chlidonias n. nigra, Locustella naevia, Branta leucopsis, Tadorna tadorna) // Ornis Fennica. V. 17 (1). P. 8–9.
Prop J., de Vries J., 1993. Impact of snow and food conditions on the reproductive performance of barnacle geese Branta leucopsis // Ornis Scandinavica. V. 24. P. 110–121.
Prop J., van Eerden M.R., Drent R.H., 1984. Reproductive success of the barnacle goose in relation to food exploitation on the breeding grounds, western Spitsbergen // Norsk Polarinstitutt Skrifter. V. 181. P. 87–117.
Guo Q., 2014. Central-marginal population dynamics in species invasions // Frontiers in Ecology and Evolution. V. 2. № 23. 17 p. https://doi.org/10.3389/fevo.2014.00023
Rutschke E., 1989. Die Wildenten Europas: Biologie, Ökologie, Verhalten. Berlin. 368 p.
Shilin M., Chusov A., Lednova J., Kouzov S., 2014. Variety and vulnerability of waterbird community in the eastern part of the Gulf of Finland in the zone of “Nordstream” marine gas pipeline // Baltic International Symposium (BALTIC). IEEE/OESDOI: 10.1109. BALTIC. 2014.6887863. P. 1‒6.
Valkama J., Vepsäläinen V., Lehikoinen A., 2011. 3rd Finnish Bird Atlas. Finnish Museum of Natural History and The Ministry of the Environment. URL: http://atlas3.lintuatlas (Дата обращения: 20.09.2015).
Väänänen V.-M., Nummi P., Lehtiniemi T., Luostarinen V.-M., Mikkola-Roos M., 2011. Habitat complementation in urban barnacle geese: from safe nesting islands to productive foraging lawns // Boreal Environment Research. V. 16 (suppl. B). P. 26–34.
Väsänen R.A., Lammi E., Koskimies P., 1998. Muuttuva pesimälinnusto. Otava, Keuruu. 567 p.
Voslamber B., van der Jeugd H.P., Koffijberg K., 2007. Numbers, trends and distribution of breeding goose populations in the Netherlands // Limosa. V. 80. P. 1–17.
Zaynagutdinova E., Kouzov S., Batova P.R., Michailov Y.M., Kravchuk A.V., 2019. Spring migration stopovers of swans Cygnus sp. in the Russian part of the Gulf of Finland. Wildfowl. Special Issue 5. P. 123–138.
Zimin V.B., Artemyev A.V., Lapshin N.V., 2002. Survey of spring bird migrations and stopovers in the Olonits fields in Karelia // Study of the Status and Trends of Migratory Bird Populations in Russia. 4-rd issue. St.-Petersburg. P. 18–28.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал