

Лесоведение, 2019, № 3, стр. 215-227
Биотопическая и экологическая приуроченность черники, брусники и голубики в Пинежском заповеднике
С. Ю. Попов *
Московский государственный университет имени М.В. Ломоносова, биологический факультет
119992 Москва, Ленинские горы, l, стр. 12, Россия
* E-mail: s_yu_popov@rambler.ru
Поступила в редакцию 03.04.2018
После доработки 25.04.2018
Принята к публикации 04.02.2019
Аннотация
Vaccinium myrtillus, V. vitis-idaea, V. uliginosum в лесной зоне на территории Европейской России являются широко распространенными видами. На основе обработки 584 геоботанических описаний, собранных в Пинежском заповеднике, составлено представление о всем спектре биотопов и местообитаний данной территории. Проведен анализ биотопического распределения этих трех видов и изучена их экологическая приуроченность к факторам увлажненности, трофности, кислотности, богатства почв азотом и освещенности, оцененных по шкалам Д.Н. Цыганова. Биотопическая приуроченность трех видов рода Vaccinium включает относительно широкий спектр местообитаний – от влажнотравных до черничных лесов. В небольших количествах они могут быть встречены на болотах всех типов генезиса. Фитоценотические оптимумы трех видов различаются довольно хорошо: для черники это нормально дренированные черничные типы леса, для брусники – чуть более дренированные лиственничники, для голубики – недостаточно дренированные сосняки долгомошные. Значения экологических факторов, рассчитанные по шкале Д.Н. Цыганова, отражают экологические предпочтения изучаемых видов. Почти по всем факторам виды имеют хорошие различия на высоком уровне значимости.
Vaccinium myrtillus, V. vitis-idaea и V. uliginosum – обычные виды разных типов хвойных лесов на территории Европейской России. Они являются, пожалуй, самыми распространенными ягодными растениями в таежной зоне. По преобладанию их в напочвенном покрове, выделяются некоторые типы леса (Сукачев, 1975). Другими словами, эти виды являются настолько обычными и сами собой разумеющимися, что изученность их экологии остается на качественном эмпирическом уровне. В основном, виды рода Vaccinium в настоящее время активно изучаются с биохимической точки зрения, и как ягодники (Jaakola et al., 2004; Демаков и др., 2010; Конюхова и др., 2013; Старицын, Беляев, 2014; Залесов, Панин, 2017; Лугинина и др., 2017).
Экологическая приуроченность трех рассматриваемых видов была уже довольно давно определена эмпирически (Ellenberg, 1974, Раменский и др., 1956; Цыганов, 1983), и с тех пор не подвергалась проверке количественными методами, за одним исключением. Для черники Л.А. Жукова с соавт. (2010) провела оценку экологической приуроченности по шкалам Д.Н. Цыганова для разных регионов Европейской России на статистической основе. Последнее время шкалам Д.Н. Цыганова в геоботанической литературе отдается все большее предпочтение, по сравнению со шкалами Л.Г. Раменского и Г. Элленберга, поскольку они дают более адекватные результаты, соответствующие как теоретическим ожиданиям, так и прямым измерениям факторов среды (Бабешина и др., 2004, 2011; Жукова, 2010; Тетерюк, 2000; Зубкова, 2011; Попов, Федосов, 2017; Беляева, Черненькова, 2018). Поэтому в настоящей работе оценка экологической приуроченности видов рода Vaccinium также оценивается по шкалам Д.Н. Цыганова.
Распространение видов рода Vaccinium по биотопам также хорошо известно, но эта информация разбросана в неявном виде среди многочисленных геоботанических работ (Сабуров, 1972; Пучнина, 2000; Василевич, 2004; Кучеров и др., 2010; Кутенков, Кузнецов, 2013; и мн. др.).
Целью данной работы является систематизация знаний по экологии Vaccinium myrtillus, V. vitis-idaea и V. uliginosum: выявление биотопического распределения и экологических предпочтений видов в северной тайге Европейской России с использованием современных количественных методов обработки экологических данных.
ОБЪЕКТЫ И МЕТОДИКА
В качестве ключевого участка для изучения биотопического распределения видов рода Vaccinium в северной тайге выбран Пинежский заповедник (его площадь 51 807.0 га). На этой территории сосредоточено высокое разнообразие ландшафтов и растительных сообществ, присущих северной тайге Европейской части России (Попов, 2016), что особенно благоприятно для изучения биотопического распределения видов.
Геоботанические описания травяно-кустарничкового и мохового ярусов во всех типах растительности (леса, луга, болота и т.д.) выполняли на пробных площадях размером 10 × 10 м. Такой размер площади является минимальным для выявления флористического разнообразия в лесных сообществах (Корчагин, 1964). Производили глазомерную оценку проективного покрытия (в % от общей площади) для каждого вида травяно-кустарничкового и мохового ярусов. При проведении полевых работ мы стремились описать всю растительность, а не только биотопы со сфагновыми мхами, для полного охвата все экологического пространства местообитаний и растительных сообществ. Описания составляли на полевых маршрутах, которые планировали с таким расчетом, чтобы они проходили основные ландшафтные урочища заповедника, занимающие разную площадь и характеризующиеся своими особенностями растительности. Таким образом, описаниями было охвачено практически полное разнообразие сообществ заповедника во всех ландшафтах, а число описаний каждого биотопа примерно пропорционально занимаемой им площади на изученной территории. Всего сделано 584 описания растительных сообществ, из них 497 выполнены С.Ю. Поповым в ходе маршрутных исследований, 87 – Л.В. Пучниной на постоянных пробных площадях и маршрутах.
По данным полевых описаний составлена классификация растительности на основе принципов, предложенных Д.Н. Сабуровым (1972) с применением современных методов многомерной статистики и ординации (Смирнов, Ханина, 2004). Проверку достоверности выделения групп ассоциаций осуществляли с помощью дискриминантного анализа. Ординацию сообществ проводили методом бестрендового анализа соответствий (DCA) в пакете PС-ORD 5.0 (McCune, Grace, 2002). 584 геоботанических описания были классифицированы в 28 групп ассоциаций.
По преобладанию видов той или иной экологической группы выделяли группы ассоциаций (серии – по Д.Н. Сабурову (1972)). В кустарничково-зеленомошных лесах выделяли черничную группу ассоциаций по преобладанию бореальных кустарничков и мелкотравья. В сфагновых лесах выделены травяно-сфагновая (по преобладанию хвощей болотного и лесного и мезотрофных видов сфагнов – Sphagnum fallax и S. riparium) и сфагновая (по преобладанию видов олиготрофно-болотных сфагнов и вересковых кустарничков с пушицей) группы ассоциаций. В травяном классе выделены влажнотравная (по преобладанию бореальных гигрофитов), широкотравная (по преобладанию видов бореального и неморального широкотравья) группы. Долгомошную группу выделяли по преобладанию видов группы хвоща лесного (Кутенков, Кузнецов, 2013). Разнотравная группа, помимо бореальных кустарничков и мелкотравья, характеризуется значительной встречаемостью (по сравнению с черничной группой) видов широкотравья. Группы ассоциаций для болот выделяли по типу питания и генезису (верховые, переходные и т.д.).
Экологические характеристики каждой группы ассоциаций определяли по шкалам Д.Н. Цыганова (1983). Экологический индекс каждого описания определялся через средневзвешенные значения обилия видов по факторам: увлажнения почв (HD), кислотности (RC), трофности (TR), обеспеченности почв азотом (NT) и освещенности (LC) по формуле:
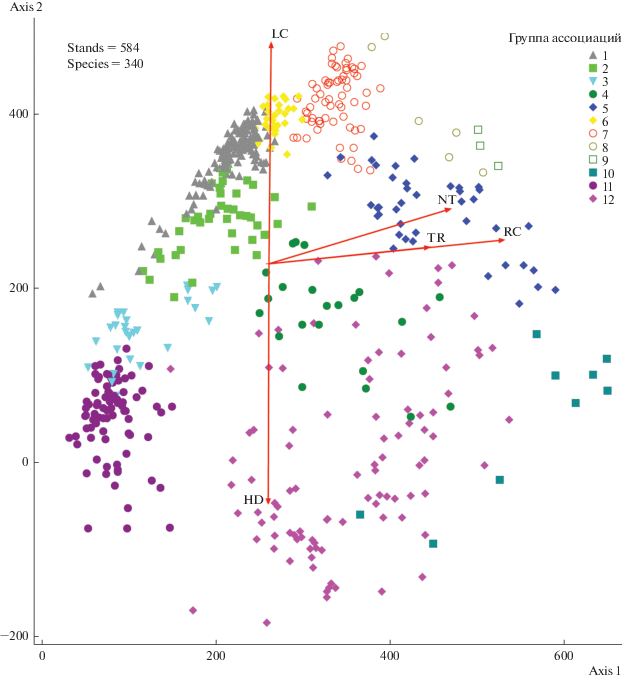
Рис. 1.
Ординация 584 геоботанических описаний в осях DCA. Группы по сходству условий местообитания: 1 – сосняки, ельники и березняки черничные; 2 – сосняки, ельники и березняки долгомошные; 3 – сосняки и ельники сфагновые; 4 – ельники, березняки, ивняки травяно-сфагновые; 5 – ельники, березняки, сероольшаники, ивняки, луга влажнотравные; 6 – ельники, лиственничники и березняки разнотравные; 7 – ельники и березняки широкотравные; 8 – луга высокотравные; 9 – луга разнотравные; 10 – болота низинные травяно-гипновые; 11 – болота верховые; 12 – болота переходные и болота переходные ключевые. Оси в центре соответствуют экологическим факторам Д.Н. Цыганова: LC – освещенность, RC – кислотность, NT – насыщенность азотом, TR – трофность, HD – увлажненность (длина и угол наклона осей соответствуют степени скоррелированности с осями варьирования DCA).
Для видов рода Vaccinium по балльным оценкам каждого описания рассчитаны основные статистические показатели: средняя, медиана, минимум, максимум и стандартное отклонение по каждому из пяти экологических факторов. Значимость различий между тремя видами по каждому экологическому фактору также проверялась по критерию Дункана.
На рис. 1 представлен результат ординации 584 описаний в первых двух осях варьирования DCA. Из рисунка видно, что значения осей имеют высокую корреляцию со значениями экологических факторов (оси в центре облака точек): ось 2 – с влажностью почвы и освещенностью, ось 1 – со значениями факторов богатства почвы. В табл. 1 приведены значения коэффициентов корреляции между осями DCA и факторами. Таким образом, рис. 1 можно рассматривать и как визуализацию распределения сообществ и местообитаний в экологическом пространстве.
Таблица 1.
Коэффициент корреляции Пирсона между значениями осей DCA и экологических факторов
| Экологический фактор | Axis 1 | Axis 2 |
|---|---|---|
| HD | –0.06 | –0.90 |
| TR | 0.83 | 0.21 |
| NT | 0.79 | 0.40 |
| RC | 0.88 | 0.31 |
| LC | 0.11 | 0.83 |
Примечание. Экологические факторы: LC – освещенность, RC – кислотность, NT – насыщенность азотом, TR – трофность, HD – увлажненность; Axis 1, Axis 2 – оси DCA (см. рис. 1).
Нагрузки на оси определяли по расстоянию Брэя–Кертиса (Съеренсена). Они составили 31.2% на первую ось и 33.6% – на вторую. Нагрузка на первую ось меньше, чем на вторую из-за того, что было применено вращение на 90°. В то же время обе оси примерно в равном соотношении отвечают за степень разброса данных. Собственные значения осей составили 0.81 и 0.56 для первой и второй осей соответственно. Длина градиента для первой оси составила 6.7 и 6.2 – для второй. Столь высокие значения перечисленных показателей говорят о высокой значимости осей ординации и отражении ими экологически верной информации (Джонгман и др., 1999).
По численным значениям экологических индексов для описаний, в которых были встречены изучаемые виды, оценивали их экологические предпочтения по каждому экологическому фактору. Биотопическое распределение оценивали на основе таблицы фитоценотической значимости видов, которая определяется через встречаемость и обилие (Работнов, 1974). Для установления сходства биотопического распределения видов использовали коэффициент Съеренсена, рассчитываемый по формуле:
где а – число биотопов для первого вида, b – число биотопов для второго вида, с – общее число биотопов для перового и второго видов (Шмидт, 1984). Статистический анализ данных производили в программе Statistica 6.0.РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Подробная характеристика синтаксонов, перечисленных в табл. 2, была дана ранее (Пучнина, 2000; Пучнина и др., 2008; Попов, Яковлева, 2008; Титова, Горячкин, 2008; Попов, 2017). Характеристика выделенных групп местообитаний (рис. 1) также была приведена в опубликованной ранее работе (Попов, Федосов, 2017), поэтому сразу перейдем к рассмотрению биотопических предпочтений изучаемых видов. Группы ассоциаций и группы местообитаний, выявленные для территории Пинежского заповедника, охватывают практически все экологическое пространство, возможное в северотаежной подзоне. Исключение, составляют только сосняки лишайниковые, которые не отмечены в заповеднике, но относительно широко распространены на севере Архангельской области (Сабуров, 1972; Кучеров, Зверев, 2012).
Таблица 2.
Биотопическое распределение и фитоценотическая значимость видов рода Vaccinium
| № | Группы ассоциаций | Vaccinium vitis-idaea | Vaccinium myrtillus | Vaccinium uliginosum | Число описаний |
|---|---|---|---|---|---|
| 1 | Ельники черничные | V2* | V4 | II2 | 69 |
| 2 | Сосняки черничные | V2 | V4 | III2 | 64 |
| 3 | Березняки черничные | V2 | V4 | III2 | 19 |
| 4 | Ельники долгомошные | V2 | V3 | III2 | 31 |
| 5 | Сосняки долгомошные | V2 | V2 | V3 | 2 |
| 6 | Березняки долгомошные | V2 | V2 | IV2 | 7 |
| 7 | Ельники сфагновые | V2 | IV2 | V2 | 12 |
| 8 | Сосняки сфагновые | IV1 | III2 | V2 | 17 |
| 9 | Ельники травяно-сфагновые | V1 | III2 | I2 | 10 |
| 10 | Березняки травяно-сфагновые | V1 | III2 | II1 | 8 |
| 11 | Ивняки травяно-сфагновые | 4 | |||
| 12 | Болота верховые | I1 | I2 | III2 | 83 |
| 13 | Болота переходные | I1 | I1 | I1 | 72 |
| 14 | Болота переходные ключевые | I1 | II2 | 25 | |
| 15 | Болота низинные травяно-гипновые | 9 | |||
| 16 | Ельники разнотравные | V2 | V2 | I1 | 12 |
| 17 | Березняки разнотравные | V2 | V2 | 12 | |
| 18 | Осинники разнотравные | V2 | V2 | 6 | |
| 19 | Лиственничники разнотравные | V3 | V2 | II2 | 7 |
| 20 | Ельники влажнотравные | IV1 | I1 | 17 | |
| 21 | Березняки влажнотравные | III1 | I1 | 6 | |
| 22 | Сероольшаники влажнотравные | 1 | |||
| 23 | Ивняки влажнотравные | II1 | 6 | ||
| 24 | Ельники широкотравные | II2 | II2 | 41 | |
| 25 | Березняки широкотравные | II1 | II2 | 24 | |
| 26 | Луга высокотравные | 6 | |||
| 27 | Луга разнотравные | 3 | |||
| 28 | Луга влажнотравные | 11 | |||
| Всего | 21 | 20 | 16 | 584 | |
| Всего групп ассоциаций, % | 75.0 | 71.4 | 57.1 | ||
Из табл. 2 видно, что черника является наиболее обильным видом в долгомошных и черничных лесах разного породного состава. Брусника хоть и встречается в этих лесах с высокой частотой, является менее обильным видом. Наибольшая частота встречаемости у голубики наблюдается в долгомошных и сфагновых лесах, что находится в полном соответствии с известными данными по экологии этих видов (Баландина, Вахрамеева, 1978, 1980; Баландина, 1983). В местообитаниях разнотравных лесов черника и брусника также встречаются более или менее постоянно, но с небольшим обилием. Голубика в таких лесах встречается значительно реже (табл. 2).
В целом из табл. 2 видно, что все три вида распространены в большинстве биотопов и отсутствуют только в наиболее богатых местообитаниях – на лугах, в сероольшаниках и в ивняках травяно-сфагновых (болотнотравных). Наиболее широкий спектр биотопов охватывает брусника – ее можно найти в 75.0% групп ассоциаций. Чуть уступает ей черника – 71.4%. Наиболее стенотопным видом из трех рассматриваемых является голубика, экологический ареал которой охватывает чуть более половины (57.1%) всего экологического пространства (табл. 2).
Несмотря на некоторую разницу в биотопическом распределении, в большинстве биотопов экологические ареалы всех трех видов перекрываются. Наибольшая степень сходства наблюдается у черники и брусники, наименьшая – у черники и голубики (табл. 3). Брусника, как более эвритопный вид встречается в наибольшем числе биотопов совместно с двумя другими видами. Биотопов, которые бы являлись уникальными для брусники и черники, нет. Везде, где встречается какой-то один из этих видов, всегда встречается хотя бы еще один из трех. Для голубики есть только один уникальный биотоп – ивняки влажнотравные, которые представляют из себя, в сущности, зарастающие ивами (Salix cinerea и S. phylicifolia) сообщества из Filipendula ulmaria или Calamagrostis langsdorffii в поймах небольших речек, где в очень незначительных количествах к евтрофному влажнотравью иногда может добавляться Vaccinium uliginosum.
Таблица 3.
Сходство между видами рода Vaccinium по биотопическому рспределению (перед дробью – число видов, после дроби – коэффициент Съеренсена)
| Vaccinium myrtillus | Vaccinium vitis-idaea | Vaccinium uliginosum | |
|---|---|---|---|
| Vaccinium myrtillus | $\frac{{20}}{1}$ | ||
| Vaccinium vitis-idaea | $\frac{{20}}{{0.98}}$ | $\frac{{21}}{1}$ | |
| Vaccinium uliginosum | $\frac{{14}}{{0.78}}$ | $\frac{{15}}{{0.81}}$ | $\frac{{16}}{1}$ |
На рис. 2–4 показано распределение видов рода Vaccinium в экологическом пространстве, образованном всем спектром биотопов, найденных в Пинежском заповеднике. Поскольку сильно дренированные местообитания (очень сухие) в заповеднике отсутствуют (Попов, 2017), верхняя часть графика рассеяния представлена точками, относящимися к нормально дренированным местообитаниям (Чертов, 1981). Это ельники, сосняки и березняки черничные, широкотравные ельники и березняки. Среди них черничные леса находятся в левой части горизонтальной оси, широкотравные – в правой. Поскольку эта ось имеет высокое сродство с факторами почвенного богатства (табл. 2), а изучаемые виды на ней “смещены” влево, можно говорить, что все три вида имеют экологический оптимум в олиготрофных или мезотрофных местообитаниях. Это так же находится в полном соответствии с тем, что нам известно об их экологии. Вообще, графики на рис. 2–4 можно рассматривать как визуализацию биотопического распространения видов Vaccinium, поскольку оси варьирования имеют высокий коэффициент корреляции с соответствующими экологическими факторами.
Рис. 2.
Фитоценотическая значимость Vaccinium mytrillus в осях непрямой ординации DCA. Диаметр значков точек увеличивается пропорционально увеличению обилия вида. Левый и нижний вспомогательные графики – диаграммы рассеяния по обилию вида относительно вертикальной и горизонтальной осей соответственно. Кривая на этих диаграммах очерчивает область рассеяния 95% точек. Обозначения те же, что на рис. 1.
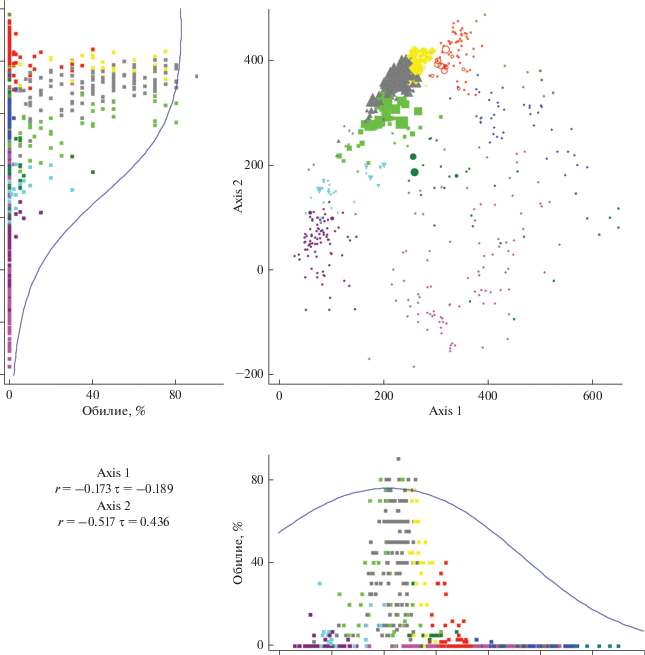
Рис. 3.
Фитоценотическая значимость Vaccinium vitis-idaea в осях непрямой ординации DCA (обозначения на рис. 1, 2).
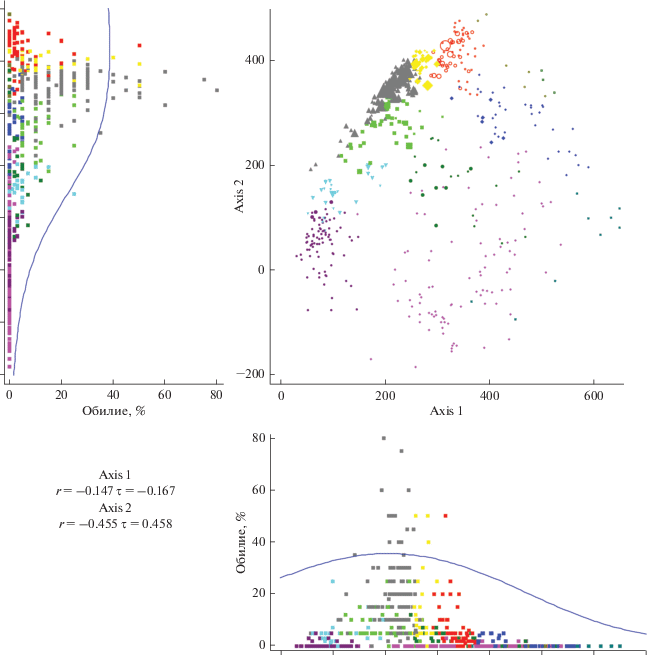
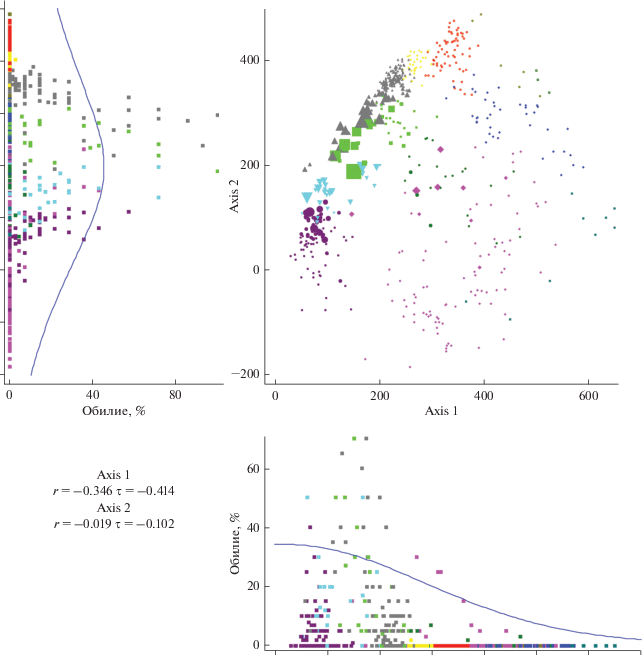
Рис. 4.
Фитоценотическая значимость Vaccinium uliginosum в осях непрямой ординации DCA (обозначения на рис. 1, 2).
Экологический (фитоценотический) оптимум вида принято определять как набор сообществ, в которых вид характеризуется наибольшими значениями встречаемости и обилия (Заугольнова, 1985). В настоящей работе в качестве фитоценотического оптимума рассматриваются сообщества, в которых класс постоянства кислицы не ниже IV, а класс обилия не ниже 3. Как видно из табл. 2, разнообразие таких сообществ в северной тайге невелико – это ельники, сосняки и березняки черничные и ельники долгомошные – для черники; лиственничники разнотравные – для брусники; сосняки долгомошные – для голубики. Леса черничного типа произрастают в нормально дренированных местообитаниях на основной поверхности водоразделов. Лиственничники разнотравные, хотя формально и причисляются к нормально дренированным, но находятся как бы на верхней границе этой градации, ближе к сильно дренированным местообитаниям. Они произрастают по вершинам всхолмлений на водоразделах и на крутых склонах коренного берега р. Сотки. Сосняки долгомошные произрастают по окрайкам болот, в депрессиях на водораздельной поверхности или на основной поверхности водоразделов, где уровень грунтовых вод повышен (Попов, 2017). Таким образом, по условиям местообитания фитоценотические оптимумы трех видов довольно сильно различаются.
Результаты обработки собранного нами массива данных показывают, что расчетные балльные оценки по пяти факторам несколько отличаются от приводимых Д.Н. Цыгановым (1983) (табл. 4). Так, например, по шкале увлажненности (Hd) для черники нижнее значение шкалы, данное этому виду Д.Н. Цыгановым, составляет 10 (лугово-степной/сухолесолуговой), верхнее – 19 (болотный). Рассчитанные нами величины таковы: нижнее значение (минимум) – 12.3 (округленно – 12, соответствует градации “сухолесолуговой/влажно-лесолуговой”), верхнее – 15.5 (округленно 15 или 16, что соответствует “сыро-лесо-луговой” или “сыро-лесолуговой/болотно-лесолуговой” соответственно). Если внимательно посмотреть на шкалу Д.Н. Цыганова, то оценка 10 – это слишком “сухо” для черники, так как 9 – это уже степные условия. В то же время “19” – слишком влажно, так как “20” относится уже к прибрежноводным местообитаниям. Аналогичные нестыковки можно найти и по другим факторам и для других видов (табл. 4). Поэтому, значения, полученные на основе обработки статистического материала, кажутся более адекватными.
Таблица 4.
Расчетные балловые значения для видов рода Vaccinium по пяти экологическим факторам и их значения по шкалам Д.Н. Цыганова (1983)
| Вид | Основные статистики | Экологические шкалы | ||||
|---|---|---|---|---|---|---|
| HD | TR | NT | RC | LC | ||
| Диапазон шкал (по Д.Н. Цыганову, 1983) | ||||||
| 1–23 | 1–19 | 1–11 | 1–13 | 1–9 | ||
| Расчетные значения | ||||||
| Vaccinium myrtillus | Mean | 14.1 | 4.3 | 3.8 | 4.6 | 4.8 |
| Median | 14.0 | 4.1 | 3.6 | 4.5 | 5.0 | |
| Minimum | 12.3 | 3.6 | 2.2 | 3.2 | 3.1 | |
| Maximum | 15.5 | 5.9 | 6.4 | 7.0 | 5.8 | |
| Std.Dev. | 0.6 | 0.4 | 0.6 | 0.6 | 0.5 | |
| Vaccinium vitis-idaea | Mean | 14.0 | 4.4 | 3.9 | 4.8 | 4.7 |
| Median | 14.0 | 4.2 | 3.7 | 4.5 | 4.9 | |
| Minimum | 11.4 | 3.6 | 2.2 | 3.2 | 3.0 | |
| Maximum | 15.9 | 6.5 | 6.4 | 8.0 | 5.8 | |
| Std.Dev. | 0.7 | 0.6 | 0.8 | 1.0 | 0.6 | |
| Vaccinium uliginosum | Mean | 14.6 | 4.1 | 3.3 | 4.1 | 4.1 |
| Median | 14.8 | 4.0 | 3.4 | 4.3 | 4.0 | |
| Minimum | 12.3 | 3.5 | 2.0 | 2.5 | 3.0 | |
| Maximum | 16.2 | 5.4 | 4.5 | 7.4 | 5.4 | |
| Std.Dev. | 0.8 | 0.2 | 0.5 | 0.8 | 0.7 | |
| Значения шкал по Д.Н. Цыганову, 1983 | ||||||
| Vaccinium myrtillus | Minimum | 10 | 1 | 1 | 1 | 2 |
| Maximum | 19 | 7 | 7 | 6 | 9 | |
| Vaccinium vitis-idaea | Minimum | 10 | 1 | 1 | 1 | 1 |
| Maximum | 17 | 7 | 5 | 6 | 8 | |
| Vaccinium uliginosum | Minimum | 11 | 1 | 1 | 1 | 1 |
| Maximum | 20 | 7 | 7 | 4 | 6 | |
Примечание. Экологические факторы см. табл. 1.
Расчетные значения среднего и медианы по каждому из факторов достаточно близки, а разброс их незначителен, о чем говорят небольшие величины стандартных отклонений (табл. 4). Таким образом, расчетные значения по экологическим факторам, приведенные в табл. 4, можно рассматривать, как исправленные балльные оценки по каждому из 5 факторов шкал Д.Н. Цыганова.
Оценка значимости различий расчетных величин экологических факторов между тремя видами показывает, что они хорошо различаются на высоком уровне достоверности (табл. 5). Исключение имеют только черника и брусника по двум факторам – увлажненности и освещенности. Поэтому нет ничего удивительного в том, что и основные статистические параметры этих двух видов по двум данным факторам тоже слабо различаются (табл. 4).
Таблица 5.
Значимость различий (Р) экологических факторов трех видов рода Vaccinium по тесту Дункана (Duncan test)
| Виды | Vaccinium myrtillus | Vaccinium uliginosum |
|---|---|---|
| HD | ||
| Vaccinium uliginosum | 0.000009 | |
| Vaccinium vitis-idaea | 0.781490 | 0.000011 |
| TR | ||
| Vaccinium uliginosum | 0.000024 | |
| Vaccinium vitis-idaea | 0.000050 | 0.000011 |
| NT | ||
| Vaccinium uliginosum | 0.000009 | |
| Vaccinium vitis-idaea | 0.011882 | 0.000011 |
| RC | ||
| Vaccinium uliginosum | 0.000009 | |
| Vaccinium vitis-idaea | 0.000313 | 0.000011 |
| LC | ||
| Vaccinium uliginosum | 0.000011 | |
| Vaccinium vitis-idaea | 0.075583 | 0.000009 |
Примечание. Полужирным выделены значения Р < 0.05; экологические факторы см. табл. 1.
Заключение. Биотопическое распределение Vaccinium myrtillus, V. vitis-idaea, V. uliginosum имеет много общего. На большей части своего экологического ареала эти виды перекрываются. Черника и брусника не встречаются ни в одном из биотопов отдельно хотя бы от одного вида из трех. Для голубики уникальным биотопом является ивняк влажнотравный в поймах малых рек. Однако экологические оптимумы трех рассматриваемых видов различаются довольно сильно. Для черники – это леса различного породного состава черничного типа, для брусники – лиственничники, для голубики – сосняки долгомошные. Статистический анализ показывает, что по предпочтительности к экологическим факторам все три вида имеют различия на высоком уровне достоверности, за исключением черники и брусники, предпочтения которых для исследованной территории не имеют достоверных различий по отношению к режиму увлажненности и освещенности.
Список литературы
Бабешина Л.Г., Дмитрук В.Н., Дмитрук С.Е. Экологические группы сфагновых мхов Томской области // Доклады Томского государственного университета систем управления и радиоэлектроники. 2004. Т. 1. № 9. С. 61–63.
Бабешина Л.Г., Рогова Н.С., Рыжакова Н.К., Зверев А.А., Меркулов В.Г. Корреляционная зависимость между содержанием химических элементов в сфагновых мхах и их экологическими оптимумами по трофности и увлажнению // Вестник Томского государственного университета. 2011. Т. 2. № 14. С. 122–131.
Баландина Т.П. Vaccinium uliginosum L. // Биологическая флора Московской области. М.: издательство МГУ. 1983. Т. 7. С. 177–187.
Баландина Т.П., Вахрамеева М.Г. Vaccinium myrtillus L. // Биологическая флора Московской области. М.: издательство МГУ. 1978. Т. 4. С. 167–178.
Баландина Т.П., Вахрамеева М.Г. Vaccinium myrtillus L. // Биологическая флора Московской области. М.: издательство МГУ. 1980. Т. 5. С. 132–146.
Беляева Н.Г., Черненькова Т.В. Связь свойств местообитаний и состава сообществ хвойно-широколиственных лесов // Экология. № 2. 2018. С. 101–109.
Василевич В.И., Бибикова Т.В. Травяные ельники Европейской России // Ботанический журнал. 2004. Т. 89. № 1. С. 13–27.
Дегтева С.В., Железнова Г.В., Пыстина Т.Н., Шубина Т.П. Ценотическая и флористическая структура лиственных лесов европейского Севера. Спб.: Наука, 2001. 269 с.
Демаков Ю.П., Богданов Г.А., Богданова Л.Г. Влияние погодных условий на урожайность ягодников в заповеднике “Большая Кокшага” // Принципы и способы сохранения биоразнообразия. Йошкар-Ола, 22–26 сентября 2010 г. Йошкар-Ола: Издательство МарГУ-ЦЭПЛ РАН, 2010. С. 305–307.
Джонгман Р.Г.Г., Тер Брак С.Дж.Ф., Ван Тонгерен О.Ф.Р. Анализ данных в экологии сообществ и ландшафтов. Пер. с англ. под ред. А.Н. Гельфана, Н.М. Новиковой, М.Б. Шадриной. М.: Издательство Российской академии сельхознаук, 1999. 306 с.
Жукова Л.А., Дорогова Ю.А., Турмухаметова Н.В., Гаврилова М.Н., Полянская Т.А. Экологические шкалы и методы анализа экологического разнообразия растений. Под ред. Л.А. Жуковой. Йошкар-Ола: Издательство Марийского гос. университета. 2010. 368 с.
Залесов С.В., Панин И.А. Ресурсы ягодных кустарничков в ельнике мшистом Североуральской среднегорной лесорастительной провинции // Лесной вестник. 2017. Т. 21. № 1. С. 21–27.
Заугольнова Л.Б. Понятие оптимумов у растений // Журн. общей биологии. 1985. Т. 46. № 4. С. 444–452.
Заугольнова Л.Б., Ханина Л.Г., Комаров А.С., Смирнова О.В., Попадюк Р.В., Островский М.А., Зубкова Е.В., Глухова Е.М., Паленова М.М., Губанов В.С., Грабарник П.Я. Информационно-аналитическая система для оценки сукцессионного состояния лесных сообществ. Пущино: ОНТИ ПНЦ РАН. 1995, 51 с.
Зубкова Е.В. О некоторых особенностях диапазонных экологических шкал растений Д.Н. Цыганова // Известия Самарского научного центра Российской Академии Наук. 2011. Т. 13. № 5. С. 48–53.
Конюхова О.М., Масленникова К.А., Канарский А.В. Взаимосвязь урожайности черники обыкновенной с фитобиоценозом // Вестник Казанского технологического университета. 2013. Т. 16. № 22. С. 222–224.
Корчагин А.А. Видовой (флористический) состав растительных сообществ и методы его изучения // Полевая геоботаника. Заложение полевых профилей и пробных площадей. М.–Л.: Наука, 1964. С. 39–62.
Кутенков С.А., Кузнецов О.Л. Разнообразие и динамика заболоченных и болотных лесов Европейского Севера России // Разнообразие и динамика лесных экосистем России / Под ред. А.С. Исаева. М.: товарищество научных изданий КМК, 2013. С. 152–204.
Кучеров И.Б., Зверев А.А. Лишайниковые сосняки средней и северной тайги Европеской России // Вестник Томского гос. университета. Биология. 2012. Т. 3. № 19. С. 46–80.
Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. Еловые леса национального парка “Кенозерский” (Архангельская обл.) // Ботанический журн. 2010. Т. 95. № 9. С. 1268–1296.
Лугинина Е.А., Егошина Т.Л., Капустина Н.В. Ресурсная характеристика голубики топяной (Vaccinium uliginosum L.) в таежной зоне России // Известия Самарского научного центра Российской Академии Наук. 2017. Т. 19. № 2(3). С. 468–472.
Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное состояние основных концепций). Уфа: Гилем, 1998. 413 с.
Попов С.Ю. Актуализация ландшафтной карты Пинежского заповедника методами многомерного анализа // Nature Conservation Research. Заповедная наука. 2016. Т. 1. № 1. С. 11–22.
Попов С.Ю. Растительность Пинежского заповедника (юго-восток Беломорско-Кулойского плато) // Флора и растительность Беломорско-Кулойского плато. А-рхангельск: Северный Арктический Федеральный Университет, 2017. С. 131–184.
Попов С.Ю., Федосов В.Э. Ценотическое распределение и экологические предпочтения сфагновых мхов (Sphagnaceae) в северной тайге европейской россии (Пинежский заповедник, Архангельская область) // Труды Карельского НЦ РАН. 2017. № 9. С. 3–29.
Попов С.Ю., Яковлева А.И. Растительность ключевого участка Пинежского заповедника // Компоненты экосистем и биоразнообразие карстовых территорий Европейского Севера России. Архангельск: ФГУ заповедник “Пинежский”, 2008. С. 85–122.
Пучнина Л.В. Растительность // Структура и динамика природных компонентов Пинежского заповедника. Архангельск: ФГУ заповедник “Пинежский”, 2000. С. 78–90.
Пучнина Л.В., Попов С.Ю., Мязгова Н.А., Чуракова Е.Ю., Ланчиков И.В. Характеристика болот и болотной флоры Пинежского заповедника // Компоненты экосистем и биоразнообразие карстовых территорий Европейского Севера России. Архангельск: ФГУ заповедник “Пинежский”, 2008. С. 123–135.
Работнов Т.А. Луговедение. М.: Изд-во МГУ, 1974. 384 с.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипов Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 472 с.
Сабуров Д.Н. Леса Пинеги. Л.: Наука, 1972. 173 с.
Смирнов В.Э., Ханина Л.Г. Методы анализа состояния растительного покрова // Восточноевропейские леса. История в голоцене и современность. М.: Наука. Т. 1., 2004. С. 290–313.
Старицын В.В., Беляев В.В. О современном состоянии ресурсов брусники (Vaccinium vitis-idaea L.) и черники (Vaccinium myrtillus L.) в лесах Архангельской области // Arctic Environmental Research. 2014. № 2. С. 71–77.
Сукачев В. Н. Проблемы фитоценологии. Л.: Наука. Т. 3., 1975. 544 с.
Тетерюк Л.В. Опыт применения фитоиндикационных экологических шкал для выявления неблагоприятных факторов на границе распространения вида // Экология. 2000. Т. 31(4). С. 276–281.
Титова А.А., Горячкин С.В. Температурный режим как фактор устойчивости лугов карстовых логов и формирования специфических почв под ними // Компоненты экосистем и биоразнообразие карстовых территорий Европейского Севера России. Архангельск: ФГУ заповедник “Пинежский”, 2008. С. 9–15.
Халафян А.А. Математическая статистика с элементами теории вероятностей. М.: Бином, 2010. 491 с.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
Чертов О.Г. Экология лесных земель. Л.: Наука, 1981, 190 с.
Шмидт В.М. Математические методы в ботанике. Л.: издательство Ленинградского университета, 1984. 288 с.
Ellenberg H. Zeigerwerte der Gefässpflanzen Mitteleuropas. Gottingen: Goltze, 1974. 97 p.
Jaakola L., Määttä-Riihinen K., Kärenlampi S., Hohtola A. Activation of flavonoid biosynthesis by solar radiation in bilberry (Vaccinium myrtillus L.) leaves // Planta. 2004. V. 5. P. 721–728.
McCune B., Grace J.B. Analysis of Ecological communities. Gleneden Beach, Oregon: MjM Software Design, 2002. 304 p.
Дополнительные материалы отсутствуют.